中甸犏牛SLC11A1基因3′非翻译区多态位点研究及其与抗病性关联的评估
2018-09-28王利平席冬梅熊和丽李国治邓卫东
王利平,席冬梅,熊和丽,李国治,李 鹏,邓卫东*
(1.云南农业大学 动物科学技术学院,云南 昆明 650201;2.新乡学院 生命科学与技术学院,河南 新乡 453000)
SLC11A1基因,又名天然抗性相关巨噬细胞蛋白1基因(NRAMP1),编码一种跨膜蛋白,定位于巨噬细胞晚期吞噬溶酶体的膜中,该基因是主要的自然抗性相关候选基因,与多种胞内寄生病原菌的抵抗作用相关[1-3]。研究表明,SLC11A1蛋白除了对小鼠的后天获得性免疫有影响外,在小鼠先天免疫中也起着关键性的作用,可促进巨噬细胞的细菌杀伤作用[4-6]。水牛、瘤牛、黄牛以及荷斯坦牛中,SLC11A1基因与布鲁氏菌病、乳腺炎等一些疾病的抗性有关[7-11];SLC11A1基因3′非翻译区(3′UTR)的多态性与对抗或易感布鲁氏菌、牛分枝杆菌和禽分枝杆菌有关[12-16],相关研究主要集中在SLC11A1基因3′UTR区的GT重复多态性上[7,17-19]。
中甸犏牛由中甸牦牛与迪庆黄牛杂交而成,表现出很强的杂种优势[20]。中甸犏牛作为青藏高原地区特有的重要肉乳兼用牛种,是农牧民生产肉、酥油、奶渣等生活必需品的重要原料来源,特别适应中国云南省海拔3 000 m以上环境恶劣的青藏高原[21]。
目前,关于中甸犏牛SLC11A1基因3′非翻译区多态位点研究尚未见报道。为此,本研究首次克隆了中甸犏牛SLC11A1基因并扩增出3′UTR区序列,检测其多态性,评价了其变异与抗病性的关系,以期为中甸犏牛的抗病育种提供辅助选择的分子标记,为进一步研究SLC11A1基因多态性对动物机体的抗病性影响提供理论依据。
1 材料和方法
1.1 样品采集
在中国云南省迪庆州香格里拉中甸屠宰场采集114份中甸犏牛肝脏样品,迅速放置-20 ℃冰箱,用于下一步的试验研究。
1.2 主要仪器与试剂
TG16-WS台式高速离心机为上海卢湘仪离心机仪器设备有限公司产品,LifeECO基因扩增仪为杭州博日科技有限公司产品,培清JS-780全自动凝胶成像分析仪购自上海培清科技有限公司。肝脏DNA组提取试剂盒为北京全式金公司产品,2×Easy PCR Super Mix、DNA Marker均购自北京硕擎生物技术有限公司。
1.3 犏牛基因组DNA的提取
采用北京全式金公司的DNA提取试剂盒提取肝脏基因组DNA,1%琼脂糖凝胶电泳检测基因组DNA的质量,-20 ℃保存备用。
1.4 PCR扩增及测序检测核苷酸变异和3个GT重复多态位点
参考NCBI上牛SLC11A1基因序列(U12862)设计引物[22],上游引物序列为:5′-GCCACGGGTGGAATGAGT-3′;下游引物序列为:5′-TGAGCTAGGAAATAGCAGG-3′,引物由昆明硕擎生物技术有限公司合成。以提取的基因组DNA为模板,PCR扩增出犏牛SLC11A1基因3′非翻译区276 bp长度的片段(对应于U12862核苷酸位点1 738—2 013)。25 μL的PCR反应体系含2×Easy PCR Super Mix 12.5 μL,ddH2O 8.5 μL,DNA模板2 μL,上游引物(10 pmol/L) 1 μL;下游引物 (10 pmol/L) 1 μL。PCR反应程序: 94 ℃ 预变性3 min,94 ℃变性30 s,55 ℃ 退火 30 s,72 ℃ 延伸30 s,35个循环,最后72 ℃延伸5 min。PCR产物经1.5%的琼脂糖凝胶电泳检测后送昆明硕擎生物技术公司进行双向测序。
1.5 统计分析
用在线软件Shesis(http://analysis.bio-x.cn/SHEsisMain.htm) 统计3′非翻译区3个多态位点(MS1、MS2和MS3)的等位基因、基因型频率和连锁不平衡值(D′和r2),并构建主要多态位点的单倍型。用PopGen 32软件计算期望杂合度(Exp_Het)和期望纯合度(Exp_Hom)、观察杂合度(Obs_Het)和观察纯合度(Obs_Hom)、卡方值(χ2)、哈迪-温伯格值(Hardy-Weinberg,HW)、基因多样性指数(Nei)和多态信息含量(PIC),应用Clustalx 1.83程序进行序列比对。
2 结果与分析
2.1 中甸牦牛SLC11A1基因3′非翻译区3个多态位点等位基因及其频率
在检测到的3个多态位点中,MS1位点包含GT10、GT12、GT14、GT15、GT16、GT17共6个等位基因。MS2位点有GT5、GT6、GT7共3个等位基因。MS3位点有GT12、GT13、GT14、GT15、GT16共5个等位基因。GT15、GT5、GT13分别是这3个位点频率最高的等位基因,频率分别为0.284、0.685、0.433(图1)。
2.2 中甸牦牛SLC11A1基因3′非翻译区3个多态位点基因型和基因型频率
通过对114份犏牛肝脏样品SLC11A1基因3′非翻译区双向测序检测到MS1位点有16个基因型,MS2位点有5个基因型,MS3位点有12个基因型。基因型GT12/15、GT5/5、GT13/13分别是这3个多态位点频率最高的基因型,频率依次为0.204、0.500、0.324(图2)。
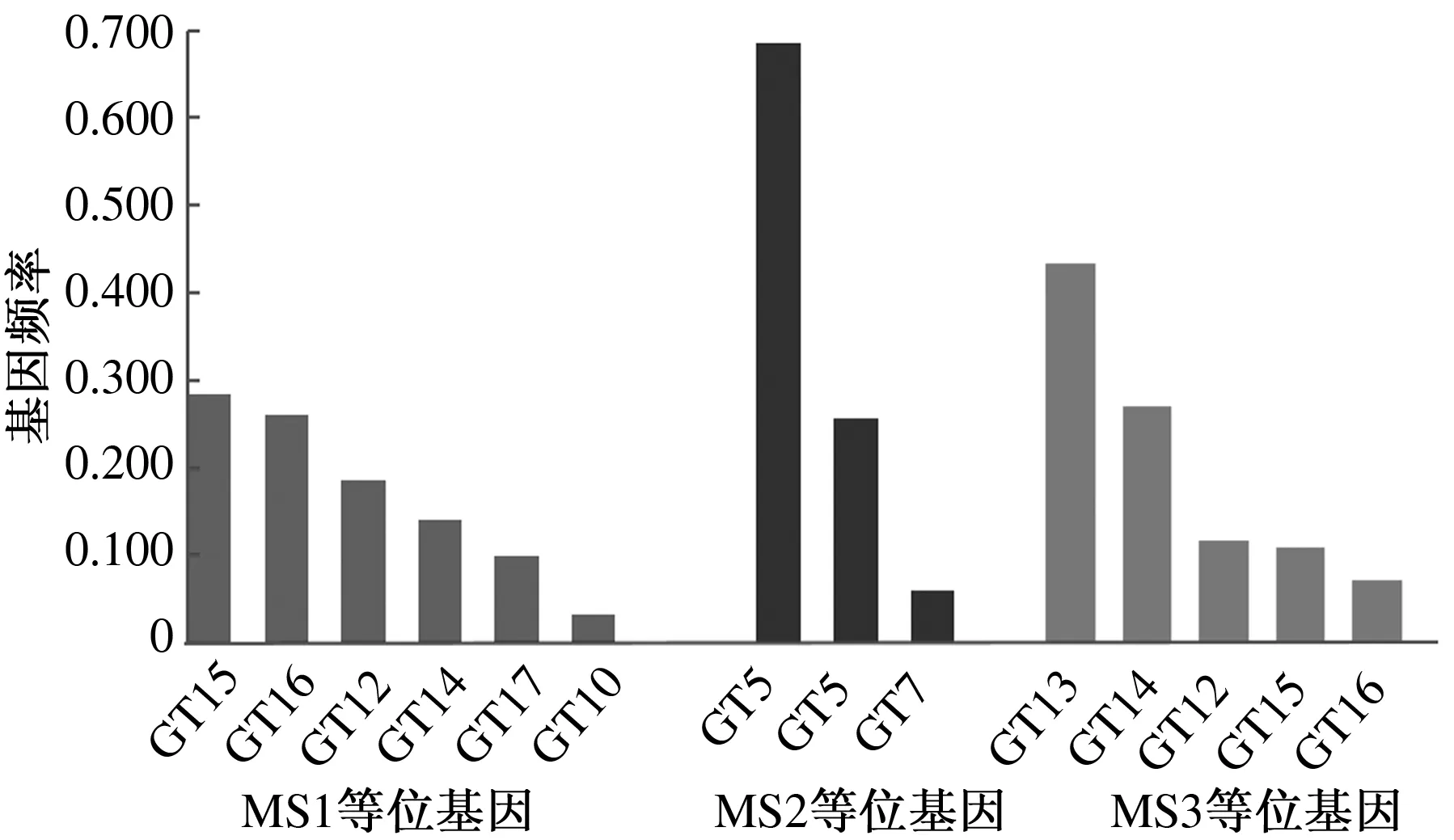
图1 中甸犏牛SLC11A1基因3′非翻译区等位基因
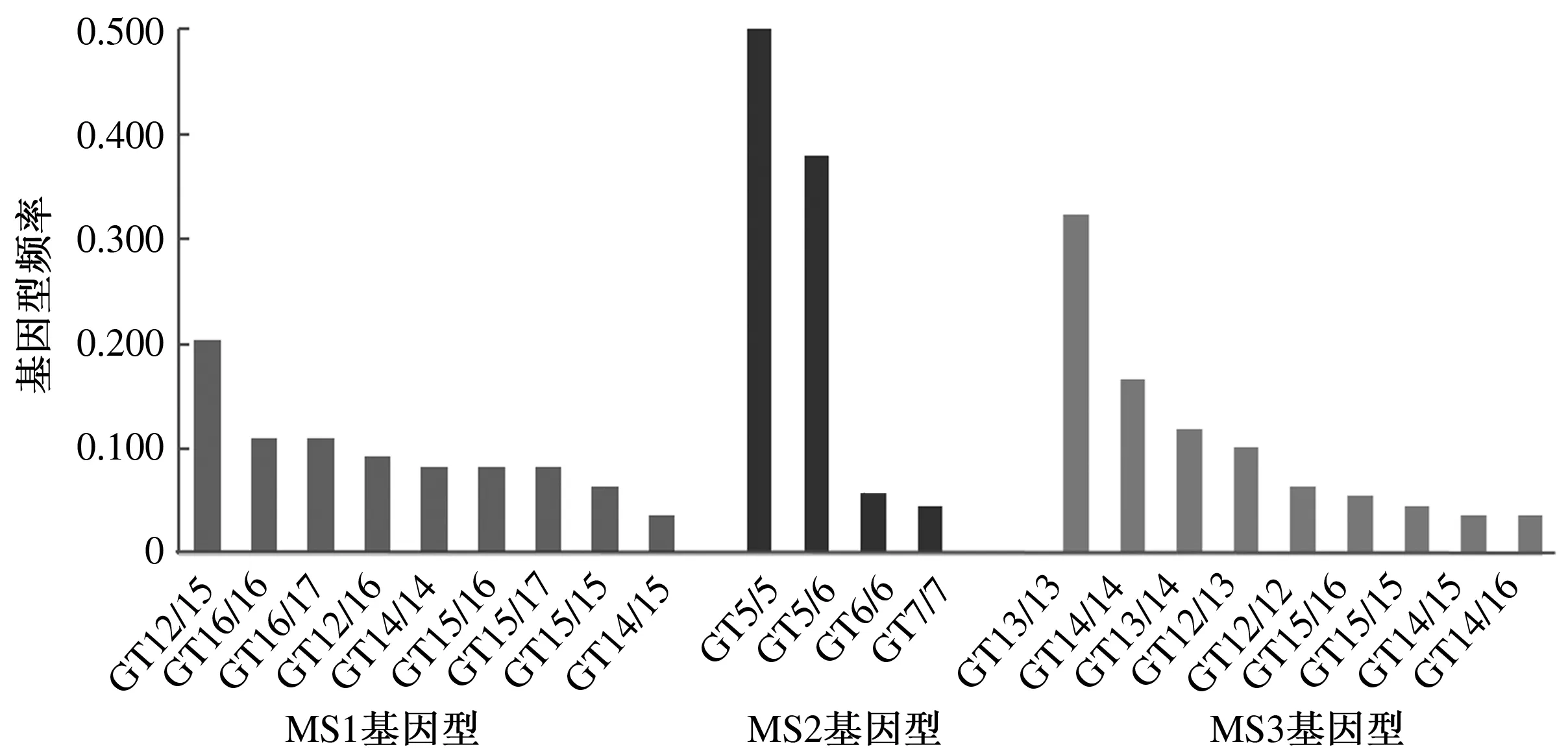
图2 中甸犏牛SLC11A1基因3′非翻译区基因型及基因型频率
MS1位点有6个等位基因和16个基因型,基因型GT12/12是黄牛频率最高的基因型被报道过,曾命名为NRAMP1.1或A[9,23]。研究证实,基因型GT12/12与抵抗牛布鲁氏杆菌抗性有关,其他杂合基因型与易感性有关[9,14]。此外,在MS1位点首次发现了GT10、GT17基因型。MS2位点有3个等位基因和5个基因型,基因型GT5/5频率最高,这个多态位点也是第一次被研究。MS3位点有5个等位基因和12个基因型,GT13和GT13/13分别是频率最高的等位基因和基因型,与以前的研究一致[8,17,24]。荷斯坦牛中GT13/13基因型频率是100%,并且该基因型与抵抗布鲁氏杆菌抗性有关[8,25]。瘤牛、瘤牛与黄牛的杂交牛中均发现了GT13,可能与牧场人为的杂交育种、基因迁移、选择育种有关[24],也可能与水牛和黄牛中SLC11A1基因较高的mRNA的转录率和稳定表达量有关[12-13,26]。
2.3 中甸牦牛SLC11A1基因3′非翻译区3个多态位点单倍型构建和单倍型频率
3个多态位点间共构建出44个单倍型,GT12/5/13、GT16/5/13和GT15/5/13是频率较高的单倍型,频率依次为0.130、0.115、0.086。除此之外,还有其他7个单倍型GT15/6/14、GT17/6/14、GT15/5/12、GT15/5/14、GT16/5/12、GT16/5/14、GT14/6/14频率高于或等于0.030(图3)。同时对MS1和MS2位点也进行了单倍型构建,共构建出24个单倍型,GT12/13、GT12/14、GT14/15是频率较高的单倍型,频率依次为0.148、0.034、0.053,除此之外,还有其他有8个单倍型GT15/12、GT15/13、GT15/14、GT15/15、GT16/13、GT16/14、GT17/13、GT17/14频率高于0.03(图4)。
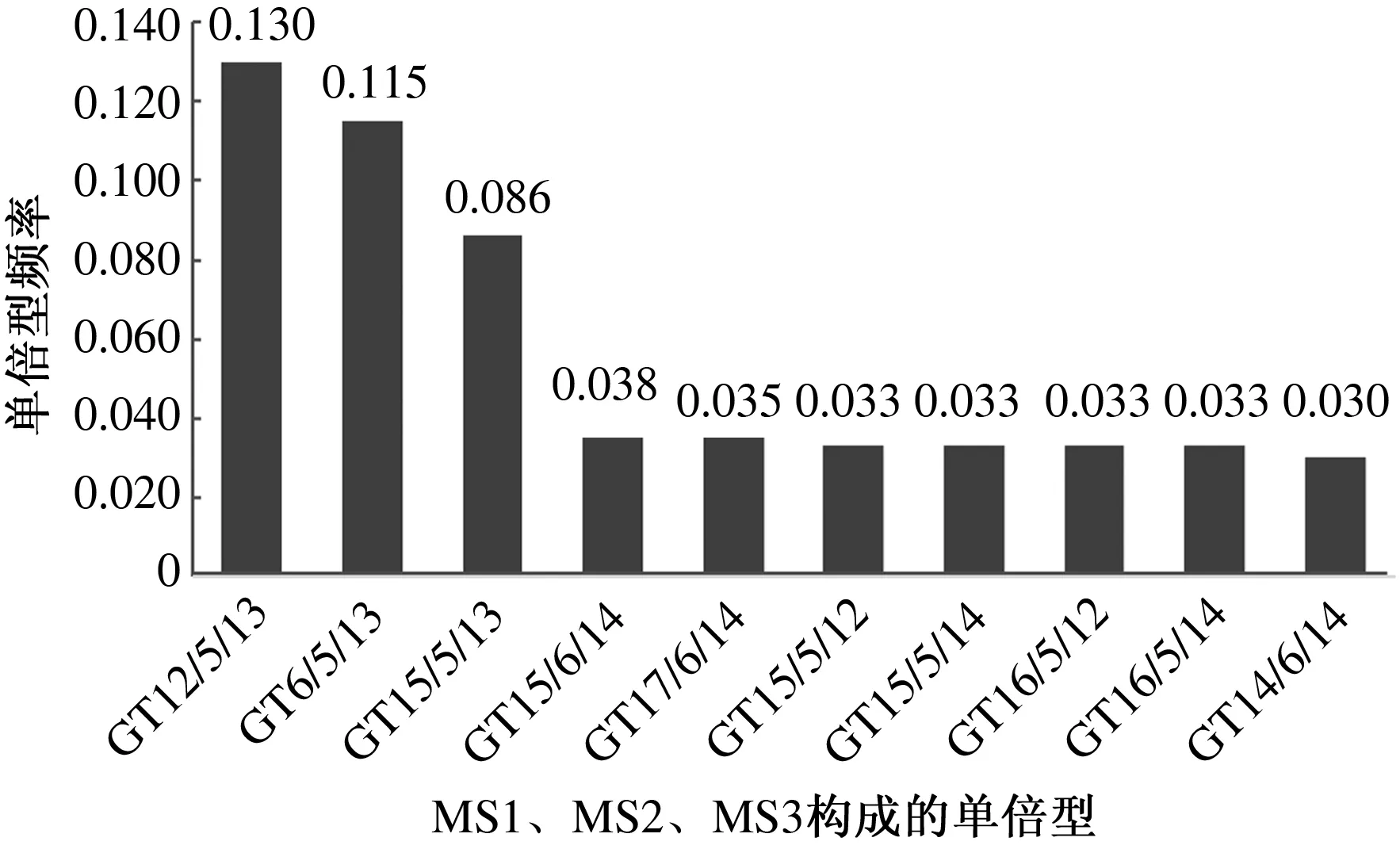
图3 中甸犏牛SLC11A1基因3′非翻译区3个多态位点(MS1、MS2、MS3)构成的单倍型频率

图4 中甸犏牛SLC11A1基因3′非翻译区2个多态位点(MS1与MS3)构成的单倍型频率
2.4 中甸犏牛SLC11A1基因3′非翻译区3个多态位点多态信息参数
MS1、MS2、MS3 多态位点的多态信息含量从MS3的0.397到MS1的0.754不等,基因多样性指数最小的是MS2位点,为0.456,最大的是MS1位点0.789。观察纯合度从MS1的0.296到MS3的0.620,观察杂合度从MS3的0.380到MS1的 0.704,期望杂合度从MS2的0.458到MS1的 0.793(图5)。利用在线软件Shesis对3个位点进行两两位点的D′和r2的计算,对位点间连锁紧密程度进行度量,位点MS1与MS2、MS1与MS3、MS2与MS3的D′值分别是0.332、0.337、0.355,r2值分别是0.023、0.026、0.044。多态位点MS1中杂合基因型的频率为0.704、MS2中纯合基因型的频率为0.602、MS3中纯合基因型的频率为0.620。
多态位点MS1、MS3都有较高的多态变异性,MS2多态水平较低。MS1的杂合度较高与多态信息含量一致。可能是因为中甸犏牛生活在青藏高原地区,自由放牧,选择压力较低,采样群体数量有限有关,与以前对哥伦比亚克里奥尔牛的研究结果一致[27-28]。D′和 r2值说明这3个多态位点的任何2个之间都存在连锁不平衡现象。

图5 中甸犏牛SLC11A1基因3′非翻译区多态位点
2.5 中甸犏牛SLC11A1基因3′非翻译区3个多态位基因型和其他牛种抗性基因型的比较
通过与其他品种牛[9,14-15,25]SLC11A1基因3′非翻译区与疾病相关的基因型(MS1的GT12/GT12和MS3的GT13/GT13)的χ2比较分析发现,中甸犏牛与婆罗门瘤牛×布兰科牛、荷斯坦牛×布兰科牛、布兰科牛、非洲牛的GT12/12基因型频率差异显著(P<0.01);中甸犏牛与西班牙黄牛、荷兰牛的GT13/13基因型频率差异显著(P<0.01),与非洲牛差异不显著(P>0.05)(表1、表2)。可见,中甸犏牛MS1的GT12/12和MS3的GT13/13基因型与抗病性有关。
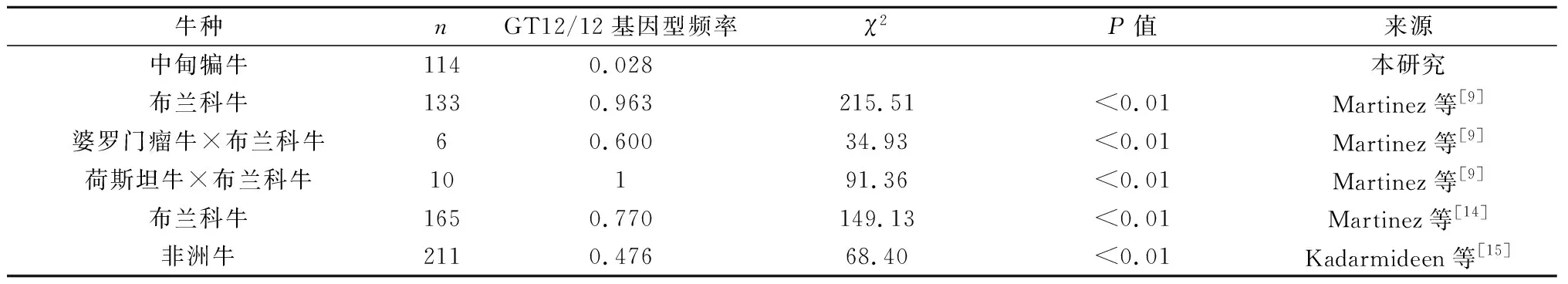
表1 中甸犏牛多态位点MS1的同质基因型GT12/12与其他品种牛相对应基因型的χ2检验结果
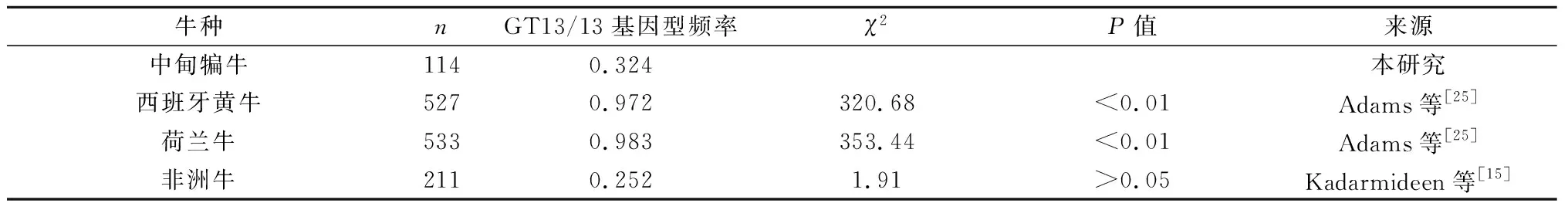
表2 中甸犏牛多态位点MS3的同质基因型GT13/13与其他品种牛相对应基因型的χ2检验结果
2.6 犏牛SLC11A1基因3′非翻译区核苷酸变异
本研究除了检测到犏牛SLC11A1基因3′非翻译区的GT重复多态外,在15头犏牛中还发现了在核酸序列1 782位点处有碱基T→G转换突变,在42头犏牛中发现1 805位点处有碱基G→A的突变,在10头犏牛中发现1 907位点处有碱基G→T的突变(图6)。
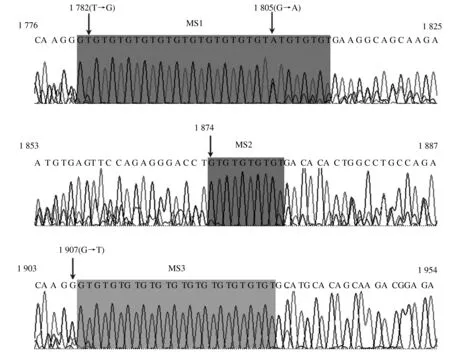
图6 中甸犏牛SLC11A1基因3′非翻译区微卫星多态位点(MS1、MS2、MS3)及其碱基突变位点
包含3个多态位点及核苷酸位点变异的犏牛SLC11A1基因3′非翻译区的7种比较典型的单倍型已提交GenBank注册,登录号分别是MH376753、MH376754、MH376755、MH376756、MH376757、MH376758、MH376759。
3 结论与讨论
3.1 不同品种牛SLC11A1基因3′非翻译区微卫星多态位点和等位基因不同
本研究扩增了中甸犏牛SLC11A1基因3′非翻译区1 738—2 013长度为276 bp的碱基片段,发现了3处分别以位点1 781、1 874、1 908(位点参照GenBank登录号U12862)为起始点的微卫星多态MS1、MS2、MS3(图6)。Kumar 等[24]扩增过此区域1 814—1 988长度为175 bp的序列,并检测到从1 908位点开始的多态位点MS3有GT13—16重复4个基因型。Borriello等[12]在水牛上也扩增过此区域1 745—1 955长度为211 bp的序列,3个多态位点分别开始于1 781、1 876、1 912位点(参照GenBank登录号U27105),并依据这3个位点所有GT重复总数分为2个单倍型,A(33个GT重复)和B(36个GT重复)。Martinez等[14]扩增出了水牛1 745—1 955长度为211 bp的序列,检测到等位基因4个单倍型(DQ095780、DQ095781、DQ376109、DQ376110)。Ganguly等[29]扩增出水牛1 804—1 996序列,并检测到从1 912位点开始的GT13—16重复4个基因型。
3.2 不同品种牛SLC11A1基因3′非翻译区微卫星多态位点核苷酸变异不同
本研究中1 782位点的碱基T→G转换突变和1 805位点处的碱基G→A的突变和以前在黄牛中的研究一致[23]。1 907位点处的碱基G→T突变是首次在中甸牦牛中发现的。1 782位点如果是碱基G,GT数量不会增加;如果是T碱基,GT重复数就增加1个。1 805位点如果是A碱基,整个GT数目会减少4个。虽然这些核苷酸变异的功能尚未明确,总体上,SLC11A1基因3′非翻译区变异与对病原微生物易感或抗性的关系已经被多次研究[12,25,30-33],这些多态位点的定位对于研究多态性与抗病性的关系具有重要的理论和实际指导意义。
3.3 中甸犏牛SLC11A1基因3′非翻译区多态位点与抗性的关联
在黄牛、瘤牛、水牛及其他反刍动物品种中都研究过SLC11A1基因3′非翻译区的多态性[7-9,11-4,18-19,23,25,34]。Borriello等[12]报道水牛中SLC11A1 AA基因型 (33GTs, DQ095780)对布鲁氏杆菌易感。Barthel等[26]报道,黄牛中含有GT13转染的细胞SLC11A1蛋白的表达量较含有GT16的高,从而限制了布鲁氏杆菌的增殖。在水牛中研究发现,含有GT13的牛能显著提高巨噬细胞的功能[29]。山羊中也发现了SLC11A1基因3′非翻译区的2个多态位点A和B, 并且发现其多态性与布鲁氏杆菌感染有关联[18]。Abraham 等[19]对山羊的研究发现,221~239 bp的B多态位点共有2个等位基因,117 bp(B7)和119 bp(B8)。含有B8等位基因的山羊较含有B7等位基因的山羊有更高的感染副结核杆菌的比率。上述研究表明,SLC11A1基因多态与对病原微生物的抗性或易感性有密切关系,并且往往频率较高的同质基因型和抗性有较密切的关系[29]。而本研究中基因型 GT12/12 和GT13/13 刚好符合这一规律。并且通过与其他牛种相对应基因频率χ2检验发现,中甸犏牛MS1的GT12/12和MS3的GT13/13基因型与抗病性有关联[8,14-15,25]。
综上,本研究揭示了中甸犏牛SLC11A1基因3′非翻译区的多态性,并对其多态性与对病原微生物的抗性进行了评估,对中甸犏牛SLC11A1基因3′非翻译区多态性的了解有助于抗病育种策略的制定和抗性牛的选育。
