菊叶香藜精油合成关键HMGR基因的 生物信息学分析
2018-08-20付苏宏张勇群郝豆豆
付苏宏,雷 鸣,张勇群,施 静,郝豆豆
(西藏自治区人民政府驻成都办事处医院 分子医学实验室,四川 成都 610041)
菊叶香藜(Dysphaniaschraderiana)为藜科1年生草本植物,具有强烈的刺激性气味,广泛生存于青藏高原草甸,此外也生长于海拔2 700~4 500 m的山坡、村边、河滩等[1]。菊叶香藜生长于辐射强、温差大的高原环境中,具有较好的极端环境耐受力,可用于改善生态环境[2]。此外,菊叶香藜具有广泛的药理活性,《藏药志》记载菊叶香藜具有抗肿瘤活性[3],全草入药可清热解毒,用于治疗感冒、头疼、皮肤瘙痒等[4]。菊叶香藜因含有挥发油而具有强烈的气味,因此对其精油的研究也日益增多。石梦菲[5]研究发现,菊叶香藜精油可抑制大肠杆菌和枯草芽孢杆菌的生长;雷鸣等[6]发现,菊叶香藜精油可显著抑制玉米象活力;刘志龙等[7]研究发现,菊叶香藜精油在防治螨虫方面具有显著效果,现已申请专利。石梦菲等[5]利用气相色谱-质谱联用(GC-MS)对菊叶香藜精油成分进行分析,结果发现,菊叶香藜精油的主要成分为脂肪族化合物(54.232%)、萜类化合物及其含氧衍生物(31.945%),其中单萜类化合物和倍半萜类化合物含量较高,主要萜类化合物有α-杜松醇(α-Cadinol,13.966%)、四甲基环癸二烯甲醇(Hedycaryol,11.164%)和δ-杜松烯(δ-Cadinene,7.292%),均为倍半萜类化合物。
高等植物萜类化合物的前体物质异戊烯基焦磷酸(IPP)可通过发生在细胞质中的甲羟戊酸(MVA)途径和发生在质体中的2-C-甲基-D-赤藓糖醇-4-磷酸(MEP)途径合成,IPP及其异构体二甲基烯丙基焦磷酸(DMAPP)经过下游多种酶的催化发生结合、缩合、修饰等方式生成结构多样的萜类化合物[8-9]。3-羟基-3-甲基戊二酰辅酶A还原酶(3-Hydroxy-3-methyl glutaryl coenzyme A reductase,HMGR)是MVA途径中的第一个限速酶,目前,小麦[10]、甘草[11]、拟南芥[12]、杜仲[13]、苹果[14]、人参[15]等多个植物的HMGR基因已经被克隆。本研究针对菊叶香藜萜类化合物合成中的关键酶HMGR进行深入剖析,利用生物信息学的方法对菊叶香藜HMGR基因编码蛋白质的理化性质、亲疏水性、二级结构、信号肽、跨膜结构、亚细胞定位、功能结构域、三级空间结构、系统进化等多个方面进行预测与分析,旨在为菊叶香藜精油生物合成的分子机制研究提供理论依据。
1 材料和方法
1.1 材料
菊叶香藜HMGR基因序列来源于菊叶香藜转录组数据库中经过比对和功能注释得到的候选基因序列(本实验室前期研究),命名为DsHMGR,已上传至GenBank数据库(序列号:MG582592);试验中所涉及到的其他氨基酸序列均来源于NCBI(National center for biotechnology information)中已登录的数据序列,相关信息如表1所示。

表1 物种信息汇总
1.2 分析方法
运用GenBank的在线软件ORF Finder和本地软件Jellyfish 2.0查找开放阅读框(ORF);利用在线软件ProtParam分析蛋白质的理化性质;运用在线分析软件ProtScale分析蛋白质亲/疏水性;运用在线SOPMA软件分析和预测蛋白质氨基酸序列二级结构;运用在线工具SignalP 4.1分析蛋白质信号肽;利用在线程序TMHMM进行蛋白质跨膜结构域的预测;运用在线软件Cell-PLoc 2.0分析蛋白质亚细胞定位;采用在线工具CDD(Conserved domain datebase)预测蛋白质的保守结构域;蛋白质三维结构以人类HMGR蛋白(PDB ID:3CD0)为模板,利用Phyre2进行三维结构同源建模;利用本地软件Clustalx进行多条氨基酸序列比对;利用本地软件MEGA 5.0构建Neighbor-joining系统进化树,相关在线生物信息学分析软件如表2所示[16]。

表2 生物信息学在线分析软件汇总
2 结果与分析
2.1 菊叶香藜DsHMGR基因序列分析
DsHMGR基因序列全长1 986 bp,GC含量为0.504,ORF Finder预测结果表明,DsHMGR包含一段157 bp的5′端非翻译区、一段104 bp的3′端非翻译区以及一段1 725 bp的开放阅读框,开放阅读框为158—1882位,编码574个氨基酸(图1),与已报道的植物HMGR基因碱基数和编码氨基酸残基数基本一致。

图1 菊叶香藜DsHMGR基因及所编码蛋白质序列
2.2 菊叶香藜DsHMGR蛋白理化性质分析
利用在线软件ProtParam对DsHMGR蛋白和3种已公布的藜科植物HMGR蛋白进行分析,并比较它们的理化性质和氨基酸组成(表3)。4种HMGR蛋白的氨基酸长度为574~581 aa,分子质量61.68~62.51 ku,等电点(pI值)6.30~7.89,不稳定系数均大于40,为不稳定蛋白质,各类氨基酸的组成基本一致,无明显差别,非极性氨基酸为主要氨基酸,比例为44.6%~45.9%。其中DsHMGR分子质量61.77 ku,pI值为7.46,不稳定系数为49.25,大于40,为不稳定蛋白质。此外,DsHMGR蛋白中非极性氨基酸所占比例最大,其比例达到了45.8%;极性中性氨基酸的比例也较高,为32.4%;酸性氨基酸与碱性氨基酸的比例相当,分别为10.3%和11.5%。DsHMGR蛋白中含量最多的氨基酸依次为亮氨酸(Leu,9.9%)、丝氨酸(Ser,9.6%)、丙氨酸(Ala,9.1%)和缬氨酸(Val,8.5%),含量最低的为色氨酸(Trp,0.3%)。

表3 藜科植物HMGR蛋白的理化性质及氨基酸组成
2.3 菊叶香藜DsHMGR蛋白亲/疏水性分析
蛋白质的疏水氨基酸残基由于水的作用会相互靠近,形成一个疏水内核和亲水表面。蛋白质的疏水作用是蛋白质折叠的主要驱动力,发挥着维持蛋白质构象的重要作用。利用在线软件ProtScale对DsHMGR蛋白进行亲/疏水性预测。正值表示疏水,其绝对值越大表示越疏水;负值表示亲水,其绝对值越大表示越亲水;介于-0.5~+0.5主要表示为两性氨基酸。DsHMGR蛋白预测结果如图2所示,在N端具有2个明显的疏水峰,分别位于40—58位和73—103位区域,提示在这2个位置可能有跨膜结构。在DsHMGR蛋白多肽链的第51位处有最高分值 3.122,其疏水性水性较强;第6位有最小分值-2.844,其亲水性较强。

图2 DsHMGR蛋白的亲/疏水性分析
2.4 菊叶香藜DsHMGR蛋白信号肽分析
信号肽是一段含有15~30个氨基酸残基的短肽,位于分泌蛋白的N端,主要包括3个区域:带正电的N末端区域、疏水的主要功能区域以及较长的带负电的C末端区域。在分泌蛋白合成结束后,其N端的信号肽将被切除。利用在线软件SignalP 4.1对DsHMGR蛋白的信号肽及其位置进行分析,结果见图3。典型的信号肽具有较高的S值,在剪切点位点处有最大的C值,Y值有最高峰。由图3知,DsHMGR蛋白的S值较小,C值未出现一个明显的最大值,Y值也未发现有最高峰,均不具有信号肽的特点,由此可推断DsHMGR蛋白不含有信号肽,为非分泌性蛋白。

图3 DsHMGR蛋白的信号肽分析
2.5 菊叶香藜DsHMGR蛋白跨膜结构域与亚细胞定位分析
在蛋白质中,20个左右的氨基酸残基所形成的α螺旋,与生物膜的脂质双分子层相互作用,将蛋白质锚定在生物膜中,这样的结构即跨膜结构域。利用在线软件TMHMM分析DsHMGR蛋白的跨膜结构域,如图4示,在N端存在2个概率约为1的最大峰,即2个跨膜结构域,分别位于氨基酸序列的37—59位和80—102位区域。同时运用在线软件Cell-PLoc 2.0对DsHMGR蛋白进行蛋白质亚细胞定位预测,结果发现,DsHMGR蛋白定位到内质网上。此外,在DsHMGR蛋白的氨基酸序列的C端发现有内质网膜蛋白的定位信号序列KKVL基序(412—415位),进一步确证DsHMGR蛋白是定位到内质网上的。因此推断菊叶香藜的DsHMGR蛋白是内质网膜上的跨膜蛋白,通过N端的2个跨膜结构将HMGR锚定到内质网上,C端则延伸至细胞质中。

图4 DsHMGR蛋白的跨膜结构分析
2.6 菊叶香藜DsHMGR蛋白二级结构分析
蛋白质的二级结构是由肽链主链骨架原子所构成的局部空间结构,不涉及氨基酸残基侧链的构象。蛋白质的二级结构主要包括α-螺旋、β-折叠、β-转角和无规则卷曲,空间上相邻的二级结构可协同完成特定的功能。利用SOPMA在线软件对DsHMGR蛋白进行二级结构预测,结果如图5所示,在DsHMGR蛋白的整个肽链中均分布有α-螺旋,是DsHMGR蛋白主要的二级结构,散布于整个肽链中,所占比例41.64%;其次为无规则卷曲,所占比例35.89%;延伸链和β-转角分别为15.85%和6.62%。

h代表α-螺旋; e代表延伸链; t代表β-转角; c代表无规则卷曲图5 DsHMGR蛋白的二级结构分析
2.7 菊叶香藜DsHMGR蛋白的保守结构域与空间结构分析
利用NCBI的CDD查找DsHMGR蛋白的保守结构域,结果显示,DsHMGR蛋白归属于HMGR家族,存在HMGR蛋白特征保守活性位点:具有催化功能的4个氨基酸残基Glu(253)、Lys(385)、Asp(461)、His(559);2个HMG-CoA结合位点EMPVGFIQIP(221—231位)和TTEGCLVA(251—258位);2个NAD(P)H结合位点DAMGMNM(347—353位)和GTVGGGT(496—502位)。在DsHMGR蛋白与其他物种HMGR蛋白的氨基酸序列比对(图6)中,DsHMGR蛋白与其他植物HMGR蛋白在C端和中间区域氨基酸序列保守性较高,HMG-CoA结合位点和NAD(P)H结合位点作为植物HMGR重要的功能结构域保守性也较高;而在HMGR蛋白的N端,除跨膜区域的保守性相对较高,其余区域的氨基酸序列则在数量和种类上均存在较大差异,预示着DsHMGR蛋白与其他物种HMGR蛋白具有相同的催化功能,但细胞定位与调控可能不同。
蛋白质的空间结构决定蛋白质功能,因此对其进行深入研究具有重要指导意义。利用线串法,将DsHMGR基因所编码的氨基酸序列提交在线三级结构预测软件Phyre2,结果显示,DsHMGR蛋白与人类HMGR蛋白(PDB ID:3CD0)具有56%的序列相似性,以该人类HMGR蛋白为模板,通过同源建模构建DsHMGR蛋白的三级结构,其结果如图7所示。与大多数植物的HMGR蛋白三级结构类似,DsHMGR蛋白具有3个功能结构域:N-结构域、L-结构域和S-结构域,跨膜区位于N-结构域内,HMG-CoA结合位点位于L-结构域内,NAD(P)H结合位点位于S-结构域内。HMG-CoA、NAD(P)H与DsHMGR蛋白结合后,Glu(253)、Lys(385)、Asp(461)、His(559)4个氨基酸残基发挥催化功能,首先由NAD(P)H和质子化的Lys(385)提供两分子H生成3-羟基-3-甲基-5-羧基戊醛辅酶A基单硫缩醛(Mevaldyl-CoA)和NAD(P)+,然后Mevaldyl-CoA脱去一分子HSCoA生成3-羟基-3-甲基-5-醛基戊酸(Mevaldehyde),最后NAD(P)H和质子化的His(559)为Mevaldehyde提供两分子H生成甲羟戊酸(Mevalonate)和NAD(P)+(图8)。

图6 HMGR蛋白多序列比对分析
2.8 菊叶香藜DsHMGR蛋白进化分析
选取14条来自GenBank数据库的HMGR蛋白序列,其物种信息详见表1,利用本地软件MEGA 5.0对DsHMGR蛋白和这14条HMGR蛋白的氨基酸序列进行系统进化分析,采用Neighbor-joining法构建HMGR分子系统进化树,进行聚类关系分析,其结果如图9所示。整个进化树聚为两大支,真菌灵芝单独聚为一支,植物聚为一支。在植物分支中,单子叶植物和双子叶植物又各自聚为一支,双子叶植物中大戟科、五加科、葫芦科、菊科、藜科各自聚为一支,具有明显的种族特异性。菊叶香藜与藜科植物位于同一分支中,说明菊叶香藜与藜科植物的亲缘关系最近。
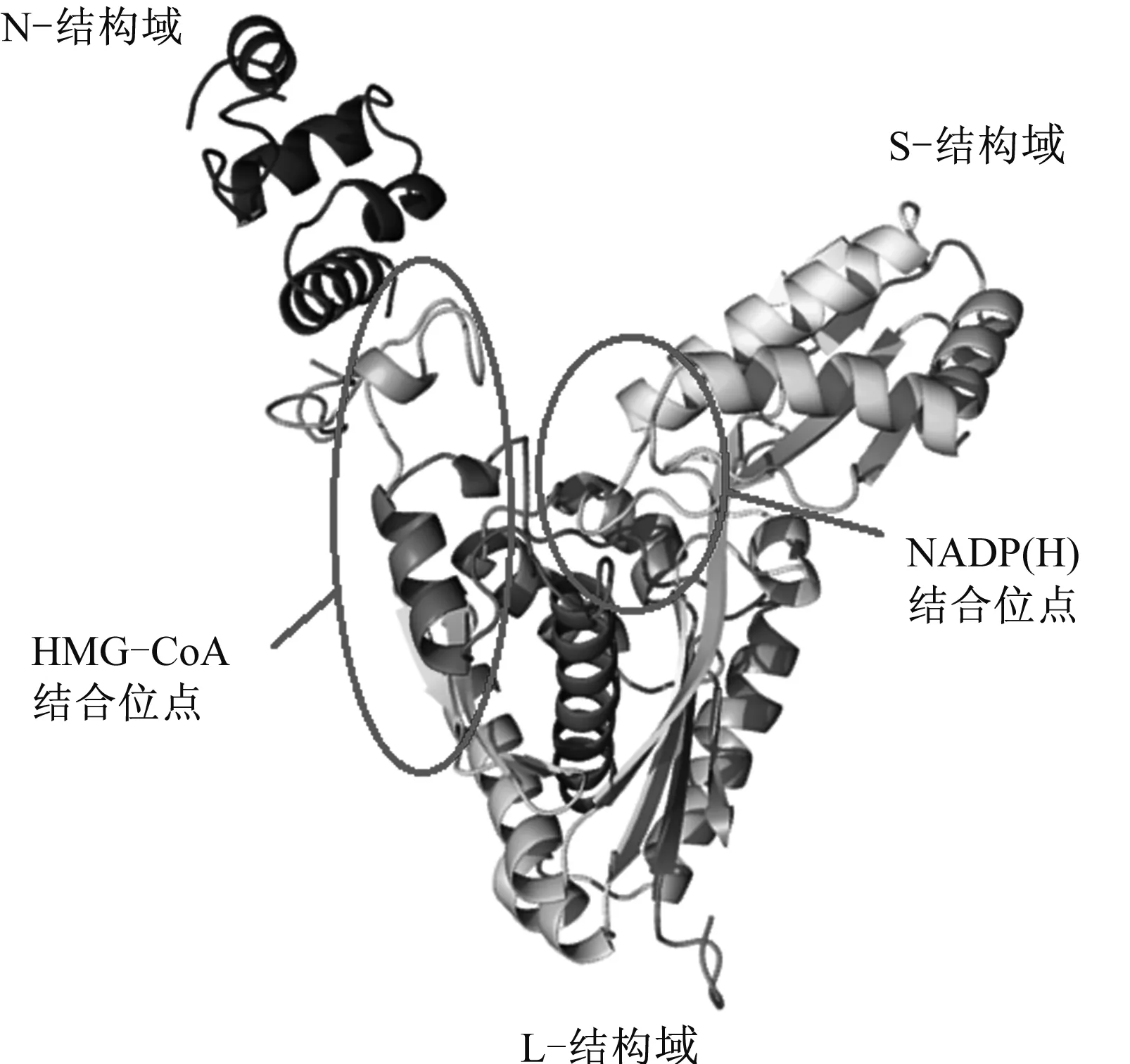
图7 DsHMGR蛋白三级结构模型预测

图8 DsHMGR蛋白的催化机制

图9 HMGR蛋白系统进化树
3 结论与讨论
植物的次生代谢产物在其生命活动和环境适应等诸多方面发挥着重要作用,除此以外,在植物次生代谢产物中发现了多种具有药用价值的化合物,如紫杉醇、青蒿素等。目前,探索植物次生代谢产物的生物合成机制以及挖掘、分析相关的酶基因已成为植物次生代谢产物研究的重要内容[15]。菊叶香藜精油表现出显著的抑菌活性和杀虫活性,其主要成分为萜类化合物。目前对菊叶香藜的研究仅限于其提取方法的优化和药理活性,其分子生物学方面的研究还很缺乏,从转录组数据库中挖掘、发现药用植物次生代谢产物生物合成关键酶基因是阐明次生代谢途径及调控机制的重要途径[17]。
MVA途径是植物合成萜类化合物前体的两大途径之一,该途径的第1个关键限速酶是HMGR,植物单萜、倍半萜、二萜、三萜类等活性成分的生物合成均需该酶的参与,通过调节该酶的表达可以决定各种萜类终产物的产量和比例,因此HMGR可作为萜类化合物代谢的重要调控位点,近年来,HMGR的结构及催化机制备受关注[16]。由此本研究对菊叶香藜转录组数据库进行挖掘,研究结论如下:(1)获得了1条HMGR基因全长序列,编码574个氨基酸,与已报道的植物HMGR氨基酸残基数相差不大,非极性氨基酸为主要氨基酸,亮氨酸(Leu,9.9%)为含量最多的氨基酸。(2)对菊叶香藜DsH-MGR蛋白进行理化性质分析,结果表明,DsHMGR蛋白不含信号肽,为不稳定蛋白质,在N端具有 2个疏水区域,含2个跨膜结构域,亚细胞定位预测与内质网膜蛋白的定位信号均显示DsHMGR蛋白定位到内质网上,与大多数植物一致[18-19]。(3)DsHMGR蛋白主要的二级结构为α-螺旋,β-折叠、β-转角和无规则卷曲穿插其中,在空间结构上则折叠为“V”形,具有N、L和S 3个催化活性中心结构域,具有4个催化氨基酸残基(253Glu、385Lys、461Asp、559His),以及2个HMG-CoA结合位点(EMPVGFIQIP和TTEGCLVA)和2个NADP(H)结合位点(DAMGMNM和GTVGGGT),与其他植物具有相同的催化活性[20-22]。(4)DsHMGR蛋白与其他植物HMGR蛋白氨基酸序列比对也显示,HMGR蛋白在C端和连接区具有较高保守性,在N端的序列保守性较低。(5)系统进化分析结果显示,菊叶香藜与藜科植物聚为一支,表明其亲缘关系较近。菊叶香藜萜类化合物合成关键酶HMGR的生物信息学分析,为今后研究菊叶香藜HMGR蛋白的功能、构建表达载体、萜类化合物的生物合成分子机制研究与开发应用奠定了良好的基础。
