枯水椪柑果肉挥发性物质及萜烯类合成途径基因表达特征
2018-05-02姚世响邓丽莉曾凯芳
姚世响 - 曹 琦 谢 姣 邓丽莉 - 曾凯芳 -
(1. 西南大学食品科学学院,重庆 400715;2. 重庆市特色食品工程技术研究中心,重庆 400715)
枯水常称粒化,是柑橘的一种常见生理性病害,多发生于采后贮藏中[1]。枯水果实在外观方面与正常果实并无二致,果皮表面新鲜,但果肉品质显著劣变,糖酸风味变淡,严重时不堪食用。枯水是当前影响柑橘果实贮藏寿命的重要因子,严重制约柑橘的商品价值。自1934年报道柑橘枯水至今,已过去80多年,科学家[2-5]从多方面对柑橘枯水进行了研究,但对枯水的发生机制仍无明确结论。
柑橘果肉的香气物质来源于果实中合成的挥发性物质[6]。目前对不同柑橘果实挥发性物质的分析研究发现萜烯类物质是含量最大的一类[7-8]。对柑橘萜烯类物质代谢途径的研究是近年的研究热点[9-10]。尽管柑橘在贮藏后期发生枯水时,会普遍伴随着香气物质的变淡,但这种现象缺乏试验证据的支撑。椪柑(CitrusreticulataBlanco cv. Ponkan)含糖量高、香气浓郁,深受消费喜爱,在重庆地区有较大面积的栽培[11]。本研究拟以椪柑为试验材料,运用气相-质谱(gas chromatography-mass spectrometry, GC-MS)技术和高通量测序技术(RNA-Sequencing, RNA-Seq),分析椪柑枯水时果肉挥发性物质和萜烯类化合物合成相关基因及基因表达的变化模式,探讨挥发性物质与椪柑枯水的关系及其机制,对阐明椪柑枯水的发生机制具有重要意义。
1 材料与方法
1.1 材料与试剂
椪柑:2015年12月于重庆市北碚区柑橘果园采收完全成熟的椪柑果实(C.reticulataBlanco cv. Ponkan)。果实在采收后迅速运回实验室,挑选大小、性状和色泽一致的果实,经咪鲜胺和2,4-二氯苯氧乙酸(2,4-dichlorophenoxyacetic acid,2,4-D)保鲜处理后,单果用聚乙烯薄膜套袋后贮藏于冷库(8~10 ℃)。在贮藏期间,定期取样观察枯水发生情况,在贮藏90 d后选择没有枯水症状的果实作为对照组,橘瓣大部分枯水的果实作为枯水样品;
癸酸乙酯:>98%,上海西格玛奥德里奇公司;
甲基叔丁基醚(methyl tert-butyl ether,MTBE):色谱纯,上海西格玛奥德里奇公司。
1.2 仪器与设备
气相色谱质谱联用仪:GCMS-QP2010 plus型,日本岛津制作所;
真空离心浓缩仪:ZLS-1型,湖南赫西仪器装备有限公司。
1.3 方法
1.3.1 GC-MS样品制备 准确称取1.00 g果肉样品,研磨成粉末。加入42.5 μg癸酸乙酯(内标)和5 mL MTBE,充分混匀后于低温超声60 min[12]。将有机相进行真空离心浓缩,然后用微孔滤膜(直径0.22 μm)过滤,供上机分析。
1.3.2 气相-质谱(GC-MS)条件
(1) 气相:DB-5MS石英毛细管柱(长30 m、内径0.25 mm、厚0.25 μm)为色谱柱。升温程序:以3 ℃/min升温至50 ℃(停留1 min),以2 ℃/min升温至70 ℃(停留3 min),以3 ℃/min升温至160 ℃(停留2 min),以8 ℃/min 升温至220 ℃(停留2 min);高纯度氦气作为载气,流速为0.80 mL/min);进样口温度:250 ℃;采用不分流进样模式。
(2) 质谱:EI源,接口温度250 ℃,电子能量70 eV;离子源温度230 ℃;扫描范围30~400 (m/z)。
1.3.3 挥发性物质的鉴定与定量 GC-MS获得的质谱图基于数据库(NIST08和NIST08S)进行检索,对挥发性物质进行鉴定,用内标法对各挥发性组分进行定量。
1.3.4 RNA-Seq分析 椪柑样品RNA用TRIzol试剂提取。cDNA文库用Ultra RNA Kit for Illumina试剂盒构建。高通量测序用Illumina Hiseq完成。测序数据用TopHat v 2.0.12软件与基因组序列进行比对,参考基因组用Citrusclementina数据(https://www.citrusgenomedb.org/analysis/156)。用FPKM(number of fragments per kilobase of transcript sequence per millions base pairs sequenced)表示基因表达水平。
1.4 数据分析与处理
挥发性成分含量的差异显著性用t检验进行统计学判断(P<0.05)。基因表达数据的差异显著性用校正的P值进行统计学判断。用Excel 2010软件进行作图。
2 结果与讨论
2.1 椪柑果肉挥发性物质组分的鉴定
图1为果肉正常果实果肉的TIC图(总离子色谱图)。由表1可知,本次试验共鉴定出13种挥发性物质,其中7种萜烯类物质、1种醇类物质、3种醛类物质和2种酯类物质。果肉挥发性物质组分似乎要少于果皮,本课题组用相同的GC-MS条件从椪柑果皮中鉴定了52种挥发性组分[8];另外前人[13-14]在印度东北地区和日本高知县的椪柑果皮中分别鉴定到37和39种挥发性组分。本研究鉴定的椪柑果肉挥发性物质组分相对较少,可能与果皮和果肉的差异相关,因为果皮油胞层可以大量合成主要成分为挥发性物质的精油;另外也与椪柑果实已经贮藏3个月有关,果实在贮藏中后期挥发性物质含量会逐渐降低,在研究中也观察到此时椪柑果实香气较淡。

图1 椪柑果肉挥发性物质总离子色谱图
Figure 1 Total ion current chromatogram of volatile compounds in juice sacs of Ponkan fruit
2.2 枯水对椪柑果肉挥发性物质组分及含量的影响
由表1可知,正常和枯水椪柑果肉中的挥发性组分均为13种,组分种类完全相同,表明挥发性物质的组成在枯水时没有发生改变。正常椪柑果实中萜烯类物质是最主要的挥发性物质,其相对含量高达88%,d-柠檬烯的相对含量为75%,表明d-柠檬烯是椪柑果实中丰度最高的萜烯类挥发性物质组分。这种挥发性物质的组成特征在莽山野柑[6]、柠檬[15]、脐橙[8]和柚类果实[16]也有类似的报道。本研究用内标法对挥发性组分进行定量分析,结果显示,枯水椪柑果肉的挥发性物质总量仅为正常果实的32%(P<0.05),其中d-柠檬烯含量仅为正常果实的13%(P<0.05)。13种挥发性组分中的5种在枯水时的含量有显著性变化(P<0.05)。
2.2.1 萜烯类物质 由表1可知,本研究共鉴定到7种萜烯类物质,其中单萜烯类有4种,分别为水芹烯、β-月桂烯、d-柠檬烯和γ-松油烯,倍半萜烯类有3种,分别为α-古巴烯、瓦伦西亚烯和α-芹子烯。这7种萜烯类物在果肉中的相对含量在枯水时降为正常果实的22.1%(P<0.05)。其中单萜烯类物质组分β-月桂烯、d-柠檬烯和γ-松油烯在枯水时含量显著降低,分别为正常果实的77.3%,13.1%,41.3%,而水芹烯的含量在枯水时没有明显改变。倍半萜烯类物质组分瓦伦西亚烯含量在枯水时升高为对照果实的163%(P<0.05),而α-古巴烯和α-芹子烯的含量则没有发生明显改变。
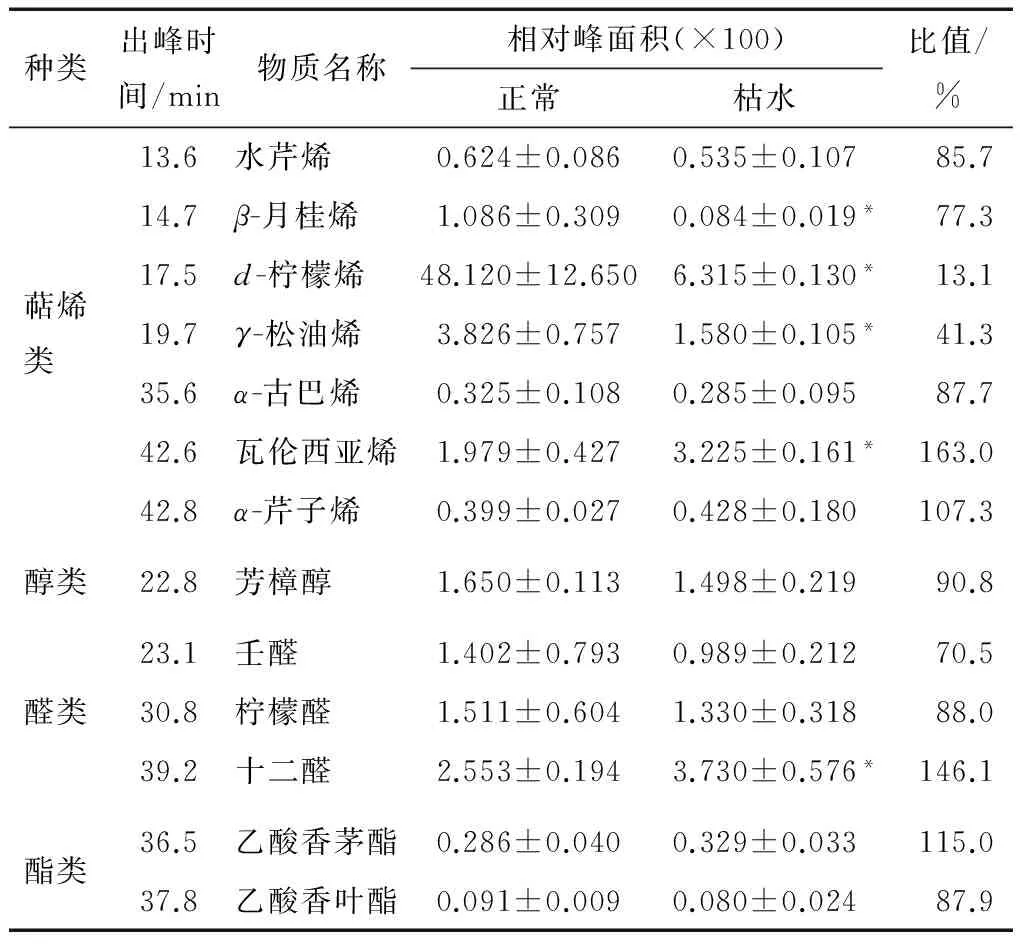
表1 椪柑果肉挥发性物质在正常果实和枯水果实中的含量†Table 1 Contents of volatile compounds in juice sacs from normal or granulated Ponkan fruit
† “*”表示差异显著,P<0.05。
2.2.2 醇类物质和醛类物质 由表1可知,本研究鉴定到1种醇类物质,即芳樟醇。芳樟醇为萜烯类衍生物,其含量在枯水时降低为正常果实的90.8%,但这种差异并不具备统计学显著性(P>0.05)。本研究还鉴定并定量了3种醛类物质,相对含量在枯水时为正常果实的110.7%,其中壬醛和柠檬醛含量分别为正常果实的70.5%和88.0%,但这种差异性并没有统计学意义的显著性(P>0.05)。十二醛相对含量在枯水时则显著升高,变为正常果实的146.1%(表1)。
2.2.3 酯类物质 本研究鉴定到2种酯类物质:乙酸香茅酯和乙酸香叶酯。这2种酯类挥发性物质的含量在枯水时分别为对照果实的115.0%和87.9%(P>0.05),因此酯类物质总量在枯水没有发生显著性变化(见表1)。
2.3 枯水对椪柑果肉萜烯类合成途径基因表达的影响
椪柑果肉挥发性物质在枯水时降低为正常果实的32%,其中相对含量为88%的萜烯类物质在枯水时降低为正常果实的22%。这意味着萜烯类物质含量的降低是椪柑果实枯水时挥发性物质含量减少的主要原因。为分析萜烯类化合物含量降低的原因,本研究采用高通量测序技术对椪柑果实枯水时基因的表达特征进行系统分析,共鉴定出8个枯水特异性表达的萜烯类物质合成途径关键酶的编码基因,表达量均显著下降,分别是ACAT2(gene encoding acetoacetyl-CoA thiolase,乙酰乙酰辅酶A硫解酶基因)、DXR(gene encoding 1-deoxy-D-xylulose 5-phosphate reductoisomerase,1-去氧-D-木酮糖-5-磷酸还原异构酶基因)、HDS(gene encoding 4-hydroxy-3-methylbut-2-enyl diphosphate synthase,4-羟基-3-甲基-2-丁烯-焦磷酸合成酶基因)、HDR(gene encoding 4-hydroxy-3-methylbut-2-enyl diphosphate reductase,4-羟基-3-甲基-2-邻苯基二磷酸还原酶基因)、KAO2(gene encoding ent-kaurenoic acid hydroxylase,贝壳杉烯酸羟化酶基因)、KAO3(gene encoding ent-kaurenoic acid hydroxylase,贝壳杉烯酸羟化酶基因)、CYP82G1(gene encoding cytochrome P450 monooxygenase,细胞色素P450单加氧酶基因)、TCS(terpenoidcyclases,萜类环化酶基因)。
2.3.1 萜类骨架代谢合成途径基因 本研究共鉴定出4个萜烯类骨架代谢合成途径的差异表达基因,见图2。萜烯类化合物主要通过甲羟戊酸(Mevalonate,MVA)途径和2C-甲基-D-赤藓糖醇-4-磷酸(2C-methyl-D-erythritol 4-phosphate,MEP)途径合成,异戊烯焦磷酸(isopentenyl pyrophosphate,IPP)和二甲基烯丙基焦磷酸酯(dimethylallyl diphosphate,DMAPP)是萜烯类物质合成的前体。本研究鉴定到基因ACAT2(CICLE_v10015452mg),其编码的乙酰乙酰辅酶A硫解酶负责在IPP合成途径中将两分子乙酰辅酶A合成乙酰乙酰辅酶A。ACAT2在枯水时表达量由正常果实的61.5下降到44.9(P<0.05)。本研究鉴定到一个编码DXR(1-deoxy-D-xylulose 5-phosphate reductoisomerase,1-去氧-D-木酮糖-5-磷酸还原异构酶)的基因DXR(CICLE_v10028082mg),参与MEP合成途径。DXR在枯水时表达量显著下调(P<0.05),由正常果实的207.4下降至枯水果实的99.3。HDS催化 1-羟基-2-甲基-2-丁烯-4-焦磷酸(1-Hydroxy-2-methyl-2-butenyl 4-diphosphate,HMBPP)的合成。本研究鉴定到一个编码HDS的基因HDS(CICLE_v10027881mg),表达量在枯水时显著下调(P<0.05),由正常果实的154.5下降至枯水果实的84.8。本研究还鉴定到HDR(CICLE_v10028384mg),编码的4-羟基-3-甲基-2-邻苯基二磷酸还原酶参与催化HMBPP生成IPP的反应。HDR在枯水时表达量由正常果实的67.9下降至52.0(P<0.05)。
2.3.2 二萜合成途径基因 本研究共鉴定出3个二萜合成途径中的差异表达基因,见图3。它们分别是编码贝壳杉烯酸羟化酶(ent-kaurenoic acid hydroxylase,KAO)和细胞色素P450单加氧酶(cytochrome P450 monooxygenase,CYP82G1)的KAO2、KAO3和CYP82G1。这3个基因在枯水时均呈显著下调的趋势(P<0.05),表达量从正常果实中的11.0,47.0,113.3下降为枯水果实中的4.9,27.3,48.9。
2.3.3 萜类物质合成途径其它基因 本研究还鉴定到编码萜类环化酶(terpenoidcyclases,TCS)的基因TCS,见图4。萜类环化酶又称萜类合成酶,是一类包括单萜合成酶、倍半萜合成酶和二萜合成酶在内的一大类酶家族。本研究鉴定的TCS在枯水表达量显著下降(P<0.05),从正常果实中的5.8下降为枯水果实中的3.2。

图2 萜烯类骨架代谢合成途径基因在椪柑果肉 枯水时的表达特征
Figure 2 Changes in expression pattern of genes involved in biosynthesis pathway of terpenoid backbone in pulp of Ponkan fruit during section-drying
果实挥发性物质的含量与其合成和降解速度密切相关,当合成减慢而降解加快时,总量必然减少。本研究基于以上基因表达结果得出结论:萜烯类物质合成途径在枯水时显著下调,是椪柑果实香气丧失和挥发性物质含量降低的重要原因。本研究所鉴定的挥发性物质组分和萜烯类合成途径差异表达基因的数量分别为13种和8个,数量较少。这可能与试验样品有关,果实已经贮藏3个月,香味偏淡,如果比较分析贮藏前与贮藏后的果实,可能会鉴定到更多的挥发性物质种类和萜烯合成相关的差异表达基因。本研究的结果基于贮藏后的正常和枯水果之间的比较,在以后研究中可对果实内部正常汁胞和枯水汁胞进行比较,结果能更好地反映枯水时的生理状态[17]。
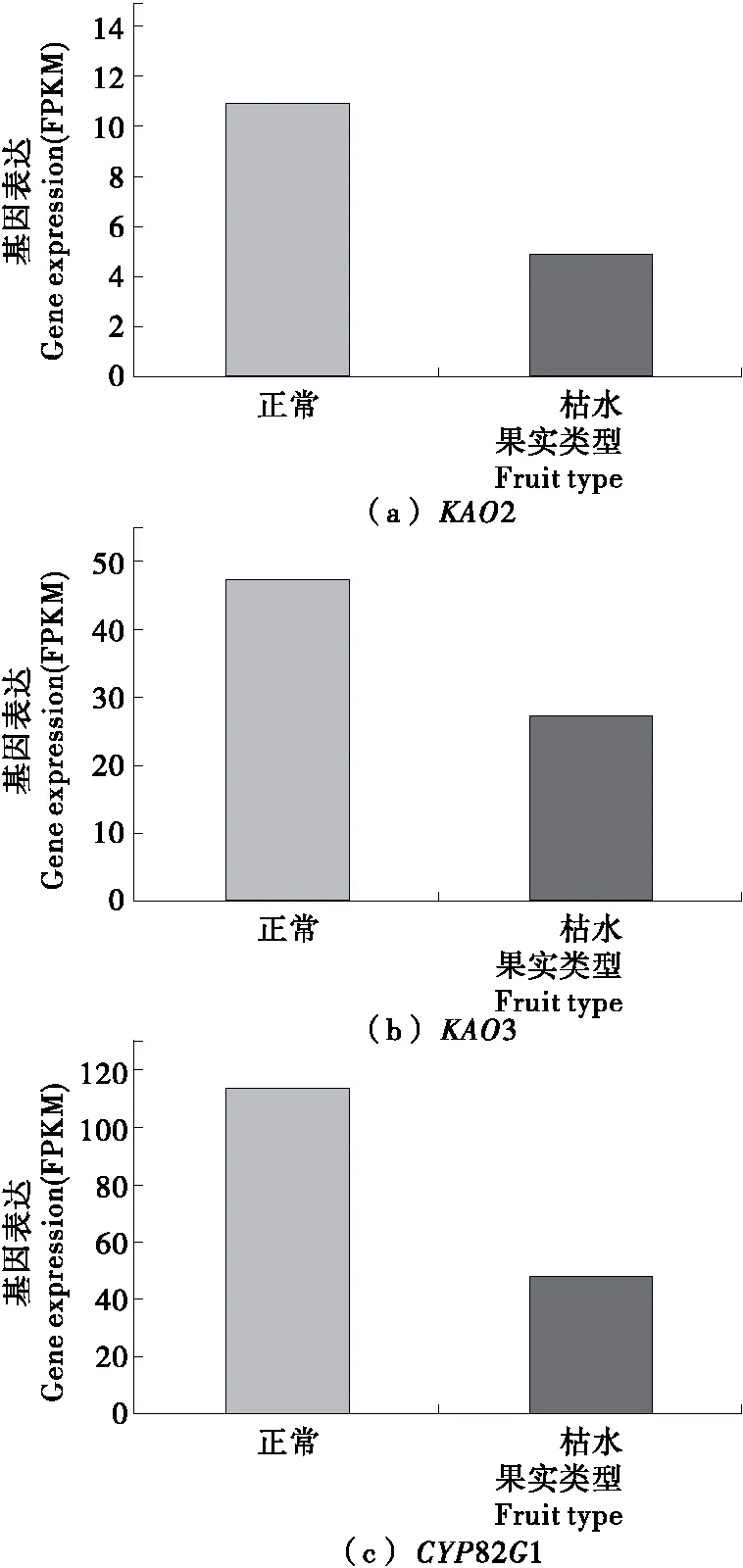
图3 二萜合成途径相关基因在椪柑果肉枯水时的表达特征
Figure 3 Changes in expression pattern of genes involved in biosynthesis pathway of diterpenoid in pulp of Ponkan fruit during section-drying

图4 萜类环化酶基因在椪柑果肉枯水时的表达特征
Figure 4 Change in expression pattern of gene encoding terpenoidcyclases (TCS) in pulp of Ponkan fruit during section-drying
本研究结果为后续深入研究椪柑果实的枯水机制提供了新的思路。一方面,可以从挥发性物质合成途径的相关基因入手,探讨其在枯水时表达量下降的调控机制,可以利用酵母单杂交技术筛选上游转录因子,最终解析枯水时挥发性物质合成的分子机制。另一方面,借助高通量代谢组学技术[18],高通量测序技术[19]、鸟枪法蛋白质组学技术[20]等,分析不同椪柑种类中不同枯水程度的汁胞挥发性物质的变化规律、转录组和蛋白质组的动态变化模式,在组学水平分析椪柑枯水的分子网络及调控机制。
3 结论
本研究发现枯水与正常椪柑果实相比,果肉挥发性物质的含量显著降低,但是组分种类没有变化。基因表达分析的结果表明萜烯类化合物合成的相关基因(ACAT2、DXR、HDS、HDR、KAO2、KAO3、CYP82G1、TCS)在枯水时表达量下调,意味着萜烯类物质的合成在枯水时受到抑制,这是椪柑枯水时挥发性物质降低的重要原因。本研究揭示了柑橘果肉挥发性物质在枯水时的降解特征,并分析萜烯类物质代谢途径相关基因的表达特征,初步阐明了椪柑枯水时挥发性物质降低的生物学机制。萜烯类合成途径相关基因的表达在枯水时为何下降,目前还不清楚,对其调控机制的研究是未来的重点。
[1] RITENOUR M A, ALBRIGO L G, BURNS J K. Granulation in Florida citrus[J]. Proceedings of the Florida State Horticultural Society, 2004, 117: 358-361.
[2] 丁健. 柑橘果实粒化变异体的遗传背景及其性状形成的机理研究[D]. 武汉: 华中农业大学, 2009: 17.
[3] WU Jia-ling, PAN Teng-fei, GUO Zhi-xiong, et al. Specific lignin accumulation in granulated juice sacs ofCitrusmaxima[J]. Journal of Agricultural and Food Chemistry, 2014, 62(50): 12 082-12 089.
[4] ZHANG Jing, WANG Miao, CHENG Fan-sheng, et al. Identification of microRNAs correlated with citrus granulation based on bioinformatics and molecular biology analysis[J]. Postharvest Biology and Technology, 2016, 118: 59-67.
[5] BURNS J K, ACHOR D S. Cell-wall changes in juice vesicles associated with section drying in stored late-harvested grapefruit[J]. Journal of the American Society for Horticultural Science, 1989, 114(2): 283-287.
[6] LIU Cui-hua, CHENG Yun-jiang, ZHANG Hong-yan, et al. Volatile constituents of wild citrus Mangshanyegan (CitrusnobilisLauriro) peel oil[J]. Journal of Agricultural and Food Chemistry, 2012, 60(10): 2 617-2 628.
[7] 张涵, 鲁周民, 王锦涛, 等. 4种主要柑橘类香气成分比较[J]. 食品科学, 2017(4): 192-196.
[8] XIE Jiao, DENG Li-li, ZHOU Ya-han, et al. Analysis of changes in volatile constituents and expression of genes involved in terpenoid metabolism in oleocellosis peel[J]. Food Chemistry, 2017, 243: 269-276.
[9] LI Xiang, XU Ya-ying, SHEN Shu-ling, et al. Transcription factor CitERF71 activates the terpene synthase geneCitTPS16 involved in the synthesis of E-geraniol in sweet orange fruit[J]. J Exp Bot, 2017, 68(17): 4 929-4 938.
[10] 冯桂蓉, 谢姣, 邓丽莉, 等. 柑橘果实萜烯类挥发性物质研究进展[J]. 食品与机械, 2017, 33(10): 200-204.
[11] 姚世响, 谢姣, 曾凯芳. 温州蜜柑和椪柑橘络主要矿质元素含量的比较分析[J]. 光谱学与光谱分析, 2017(4): 1 250-1 253.
[12] DING Yu-duan, CHANG Ji-wei, MA Qiao-li, et al. Network analysis of postharvest senescence process in citrus fruits revealed by transcriptomic and metabolomic profiling[J]. Plant Physiology, 2015, 168(1): 357-642.
[13] CHUTIA M, BHUYAN P, PATHAK M. Antifungal activity and chemical composition ofCitrusreticulataBlanco essential oil against phytopathogens from North East India[J]. LWT-Food Science and Technology, 2009, 42(3): 777-780.
[14] SAWAMURA M, THI M T N, ONISHI Y, et al. Characteristic odor components ofCitrusreticulataBlanco (ponkan) cold-pressed oil[J]. Bioscience, Biotechnology, and Biochemistry, 2004, 68(8): 1 690-1 697.
[15] LOTA M L, DE ROCCA S D, TOMI F, et al. Volatile components of peel and leaf oils of lemon and lime species[J]. Journal of Agricultural and Food Chemistry, 2002, 50(4): 796-805.
[16] NJOROGE S M, KOAZE H, KARANJA P N, et al. Volatile constituents of redblush grapefruit (Citrusparadisi) and pummelo (Citrusgrandis) peel essential oils from Kenya[J]. Journal of Agricultural and Food Chemistry, 2005, 53(25): 9 790-9 794.
[17] YAO Shi-xiang, CAO Qi, XIE Jiao, et al. Alteration of sugar and organic acid metabolism in postharvest granulation of Ponkan fruit revealed by transcriptome profiling[J]. Postharvest Biology and Technology, 2018, 139: 2-11.
[18] 吴超. 高产油微藻基于基因组的初级代谢网络重构和代谢流分析[D]. 北京: 清华大学, 2015: 8-9.
[19] VANBUREN R, BRYANT D, EDGER P P, et al. Single-molecule sequencing of the desiccation-tolerant grassOropetiumthomaeum[J]. Nature, 2015, 527(7 579): 508-511.
[20] 姚世响. 稳定同位素标记水稻幼苗的定量蛋白质组学新方法的研究[D]. 北京: 清华大学, 2015: 4-10.
