青稞籽粒灌浆期淀粉代谢酶活性与淀粉积累特征的关系研究
2018-04-09,,,,,
, , , , ,
(石河子大学农学院, 新疆 石河子 832003)
青稞亦称裸大麦,是生长在我国西北、西南的一种重要的谷类作物。淀粉是青稞籽粒的主要成分之一,占籽粒干重的65%左右。淀粉又可分为直链淀粉和支链淀粉,两者的比例是决定淀粉理化特性的重要指标,而淀粉的理化特性与青稞食品的质地和表观性状密切相关[1]。
青稞通过光合作用生成光合产物,光合产物从源器官通过一系列的维管系统以蔗糖的形式运输到库器官,之后再经过相关酶催化生成淀粉。淀粉代谢相关酶主要有腺苷二磷酸葡萄糖焦磷酸化酶(adenosine diphosphate glucose pyrophosphorylase,AGPP)、可溶性淀粉合成酶(soluble starch synthase,SSS)、束缚态淀粉合成酶(granule bound starch synthase,GBSS)、淀粉分支酶(starch branching enzyme,SBE)[2-4](合成方向)、α-淀粉酶[5](EC 3.2.1.1)和β-淀粉酶[6](EC 3.2.1.2)(降解方向)。有关淀粉积累和淀粉代谢酶的关系已经进行了许多研究[4,7],但关于青稞灌浆期内籽粒的淀粉积累特征、淀粉代谢酶活性的动态变化及二者的关系报道相对较少。本实验以3个不同品种青稞为材料,对籽粒灌浆过程中淀粉积累的特征和淀粉代谢酶活性进行系统研究,旨在探索青稞籽粒灌浆期淀粉积累特征和代谢酶学机制,为新品种的选育及品质鉴定提供理论依据。
1 材料与方法
1.1 材料与设计
选取青稞品种北青6号(淀粉含量为97.95%±1.34%[8])、昆仑12号(总淀粉含量为95.57%±0.56%[8])和甘垦5号(支链淀粉含量高达98.94%[9])为研究对象。2015年4—7月在石河子大学农学院试验站种植,人工条播,行距20 cm,株距2 cm,小区面积5 m2,3次重复。青稞开花期,挂牌、标记穗子中上部颖花在同一天开花的穗子,于花后5,10,15,20,25,30,35 d取样。每个小区取40穗,分为两部分,一部分用于淀粉含量的测定,处理方法:先将籽粒在105 ℃恒温下杀青30 min,然后将温度调至70 ℃恒温烘干至恒质量;另一部分用于酶活性测定,处理方法:将剥取的籽粒用锡箔纸包裹,侵入液氮中冷激,取出放入-80 ℃低温下备用。
1.2 测定项目与方法
1.2.1酶活性与淀粉含量的测定
参照程方民等[10]的方法进行青稞籽粒粗酶液提取;用Nakamura等[11]的方法测定AGPP、SSS和GBSS的活性变化;用李太贵等[12]的方法测定SBE的活性变化;参照何照范等[13]的双波长比色法测定淀粉含量;α-淀粉酶与β-淀粉酶活性的测定参照DNS还原糖法。
1.2.2淀粉积累特征参数的计算
根据淀粉积累量(y)与开花后天数(t)建立Logistic方程y=k/(1+eA+Bt),式中,A、B为参数,K为生长终值量。采用Curve Expert 1.4对籽粒支链淀粉和直链淀粉积累过程进行拟合[14],通过对方程求一阶导数可以得到淀粉积累速率方程,并可得出淀粉积累特征参数。积累速率最大时的日期Tmax=-A/B;积累活跃生长期(大约完成淀粉总积累量的90%)D=[ln(1/9)-A]/B;平均积累速率Vmean=K/D;最大积累速率Vmax=-KB/4;积累起始势C0=K/(1+eA)[14]。
2 结果与分析
2.1 青稞籽粒灌浆期淀粉合成酶活性
从图1 a可看出,3个青稞品种的腺苷二磷酸葡萄糖焦磷酸化酶活性均呈单峰曲线变化。北青6号的最大腺苷二磷酸葡萄糖焦磷酸化酶活性峰值和达到最大酶活性峰值的时间与昆仑12号和甘垦5号存在差异,北青6号在花后25 d达到峰值外,昆仑12号和甘垦5号均在花后20 d达到峰值;酶活性峰值表现为北青6号>昆仑12号>甘垦5号。
从图1 b可看出,非糯性青稞品种北青6号和昆仑12号分别在花后20 d和花后25 d左右达到酶活性峰值;颗粒结合型淀粉合成酶(GBSS)灌浆前期的酶活性变化在昆仑12号和北青6号中差异并不显著,而在灌浆后期两者之间的GBSS酶活性差异变大,昆仑12号的颗粒结合型淀粉合成酶活性明显大于北青6号,昆仑12号的直链淀粉最终积累量也较北青6号高。另外,颗粒结合型淀粉合成酶(GBSS)活性在糯性青稞品种甘垦5号中未检测到。由此推断,颗粒结合型淀粉合成酶(GBSS)活性与青稞籽粒直链淀粉合成有关,决定直链淀粉积累量。
从图1 c可看出,昆仑12号、北青6号和甘垦5号的SSS活性均在花后20 d左右达到峰值,然而在不同类型的青稞品种中,SSS的活性变化之间存在明显的差异。青稞品种北青6号在整个灌浆期SSS活性变化比较平缓,酶活性峰值也较小。SSS主要参与支链淀粉的合成,不同支链淀粉含量的青稞品种可溶性淀粉合成酶酶活性峰值表现为甘垦5号>昆仑12号>北青6号。这是因为甘垦5号的淀粉主要组成成分为支链淀粉,其百分含量可达98.94%,从而在整个灌浆过程中,甘垦5号的可溶性淀粉合成酶活性均大于另外2个青稞品种。
由图1 d可看出,3个青稞品种淀粉分支酶活性均呈单峰曲线,且各品种均在花后20 d左右达到峰值。各品种青稞淀粉分支酶活性大小为甘垦5号>北青6号>昆仑12号,支链淀粉含量较高的青稞品种淀粉分支酶活性峰值也较大。
2.2 籽粒灌浆期淀粉降解酶活性

图1 3个青稞品种籽粒灌浆期淀粉合成酶的活性变化
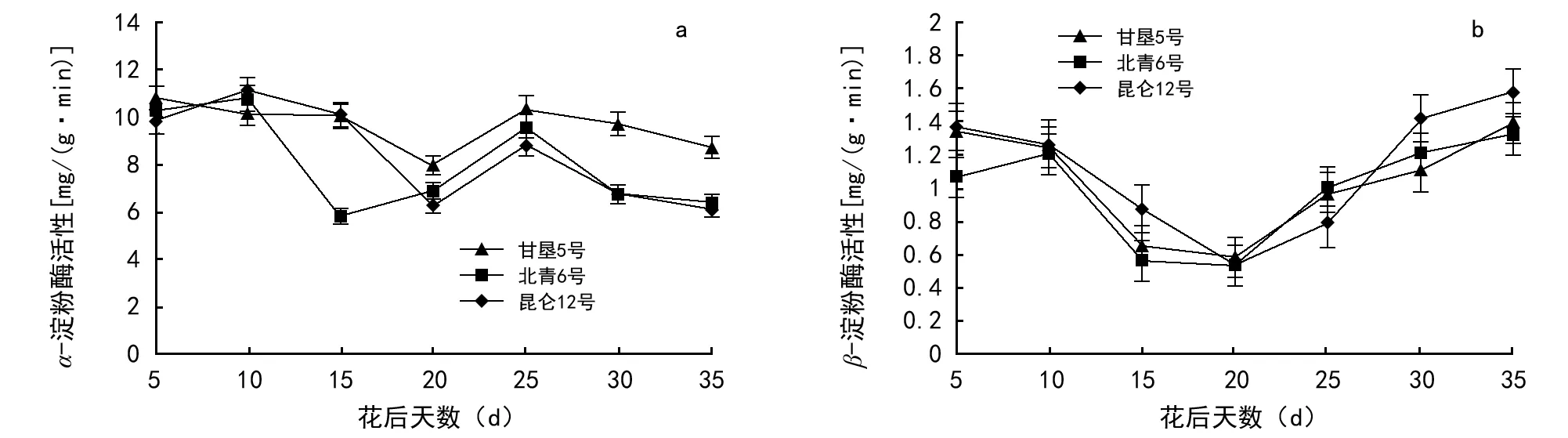
图2 3个青稞品种籽粒灌浆期淀粉分解酶的活性变化
本实验测定了3个青稞品种灌浆期淀粉降解酶(α-淀粉酶)的活性变化,结果(图1)发现,3个青稞品种的α-淀粉酶活性呈现降—升—降的趋势,且3个品种之间无显著差异。花后10 d左右最高,北青6号在花后15 d酶活性达到峰谷,而甘垦5号和昆仑12号在花后20 d达到峰谷,这可能是由于品种之间的差异造成的;3个品种的α淀粉酶活性均在花后25 d达到峰值,然后随灌浆时间延长而逐渐下降。3个品种的β-淀粉酶活性也呈降—升—降的趋势,品种之间无显著差异。花后20 d达到峰谷,花后30 d再次达到峰值,之后随灌浆时间的延长逐渐升高。
2.3 3个青稞品种籽粒直、支链淀粉和淀粉的积累
从表1可看出,青稞籽粒灌浆过程中直、支链淀粉的积累过程符合Logistic方程y=k/(1+eA+Bt)的拟合。由图3 a和表1可知,昆仑12号的直链淀粉最终积累量、起始势C0、最大灌浆速率Vmax、最小灌浆速率Vmin均高于北青6号,积累速率最大时日期Tmax和积累活跃生长期D值在2个品种之间差异较小。说明相对于北青6号,昆仑12号在籽粒灌浆期时直链淀粉的积累起始时间较早,且在整个灌浆过程中昆仑12号的直链淀粉积累速率快于北青6号,因此最终积累量高于北青6号。由此推断,直链淀粉最终积累量是由积累启动时间的早晚和积累速率的快慢决定的。另外,甘垦5号淀粉的主要组成成分是支链淀粉,在本实验中未检测到甘垦5号的直链淀粉。
从图3 b可以看出,在灌浆前期,3个青稞品种支链淀粉积累量表现为昆仑12号>北青6号>甘垦5号;而在灌浆后期,各青稞品种支链淀粉积累量表现为北青6号>昆仑12号>甘垦5号。主要是因为灌浆前期昆仑12号的C0大于北青6号和甘垦5号,而北青6号的Vmax、Vmin均大于昆仑12号和甘垦5号,且达到最高速率的时间较晚,而相对积累时间较长,因此北青6号的最终积累量相对较高。甘垦5号的C0、Vmax、Vmin相对较小,在整个灌浆过程中,甘垦5号的支链淀粉积累量相对较低。支链淀粉最终积累量也是由积累时间的早晚和积累速率的快慢决定的。

图3 3个青稞品种籽粒灌浆期淀粉(包括直链淀粉、支链淀粉和总淀粉)积累量的变化
表13个青稞品种支链淀粉和直连淀粉积累特征参数

淀粉组分品种logistic方程:y=k/(1+e(A+Bt))R2Tmax(d)D(d)Vmean[mg/(g·d)]Vmax[mg/(g·d)]C0支链淀粉甘垦5号y=9.3580/(1+e(3.7796-0.1942X))0.982319.467430.78450.3040.45420.2089Amylopectin北青6号y=19.0082/(1+e(4.1391-0.1770X))0.980923.388535.80410.53090.8410.2982昆仑12号y=16.3885/(1+e(3.3795-0.1681X))0.97720.107733.18080.49390.68860.5399直连淀粉北青6号y=5.8343/(1+e(11.5215-0.4972X))0.989623.170527.58930.21150.72530.0001Amylose昆仑12号y=7.0463/(1+e(10.5734-0.4536X))0.982323.312428.15690.25030.7990.0002
注:Tmax为速率最大时的日期;D为活跃生长期(完成淀粉总积累量的90%);Vmean为平均积累速率;Vmax为最大积累速率;C0为积累起始势。
从图3 c可看出,籽粒灌浆期北青6号与昆仑12号从开花到花后20 d淀粉的积累量与花后25 d以后淀粉的积累量呈极显著差异。而支链淀粉含量较高的甘垦5号在整个灌浆过程中呈现逐渐上升的趋势,但是淀粉积累量不存在时期上的显著差异。这可能是由于品种之间的差异造成的。
2.4 淀粉代谢酶活性与淀粉积累速率的相关性分析
前人研究表明,在4个淀粉合成酶中除AGPP活性与淀粉积累速率极显著正相关外,SSS、GBSS和SBE等单一酶活性的与淀粉积累速率并无必然联系[15]。由表2可知,青稞籽粒灌浆期淀粉合成酶AGPP、SSS、GBSS和SBE活性与总淀粉积累速率和支链淀粉积累速率均呈极显著或显著正相关。而直链淀粉积累速率与 AGPP、GBSS和SBE呈显著正相关,而SSS未呈现显著性相关,可能是由于SSS主要参与支链淀粉合成导致的。而灌浆期总淀粉积累速率、直链淀粉和支链淀粉积累速率与3个青稞品种的α-淀粉酶呈负相关,与3个青稞品种的β-淀粉酶呈显著或极显著负相关。α-淀粉酶与β-淀粉酶均为淀粉降解酶,说明β-淀粉酶活性对灌浆期总淀粉积累速率、直链淀粉和支链淀粉积累速率的调节作用较α-淀粉酶的调节作用强。
表2代谢酶活性与淀粉积累速率的相关性分析

酶品种直链淀粉积累速率支链淀粉积累速率淀粉积累速率AGPP甘垦5号-0.878∗∗0.878∗∗北青6号0.829∗0.983∗∗0.925∗∗昆仑12号0.6680.934∗∗0.871∗SSS甘垦5号-0.857∗0.857∗北青6号0.5560.848∗0.838∗昆仑12号0.570.886∗∗0.841∗GBSS甘垦5号-0.952∗∗0.592∗∗北青6号0.748∗0.854∗0.924∗∗昆仑12号0.751∗0.993∗∗0.970∗∗SBE甘垦5号---北青6号0.765∗0.878∗∗0.935∗∗昆仑12号0.760∗0.966∗∗0.944∗∗α⁃淀粉酶甘垦5号--0.0066-0.0066北青6号-0.0627-0.3668-0.2725昆仑12号-0.187-0.2947-0.3272β⁃淀粉酶甘垦5号--0.8621∗-0.8621∗北青6号-0.7543∗-0.7687∗-0.8522∗昆仑12号-0.9601∗∗-0.9355∗∗-0.9601∗∗
注:“*”、“**”分别代表0.05和0.01显著性水平。
3 讨 论
关于AGPP、SSS、GBSS和SBE这几种淀粉合成的关键酶,前人在水稻[12]、小麦[13]、玉米[14]等多种籽粒淀粉中淀粉合成酶作用的研究相对较多,且不同外界环境条件[15](包括温度、水分、氮肥)、不同品质[16]中淀粉合成酶的活性也不尽相同。本试验结果(表2)表明,青稞籽粒灌浆期支链淀粉积累速率和总淀粉积累速率与4种淀粉合成酶的活性变化均呈显著或极显著正相关。本试验中所用青稞材料根据总淀粉含量排序依次是北青6号>昆仑12号>甘垦5号,相关性分析表明,AGPP的酶活性变化与淀粉的积累速率(包括支链淀粉积累速率、直链淀粉积累速率及总淀粉积累速率)呈显著或极显著正相关,从而说明较高的AGPP活性为淀粉积累量的增加提供了更强的供给能力。因此,AGPP酶在提高青稞淀粉含量中起着关键性的作用,这与曹颖妮[2]的研究结果相一致。
邓万洪等研究发现,小麦籽粒直链淀粉含量与Waxy蛋白含量呈高度正相关,利用反义RNA技术沉默GBSS基因得到GBSS转基因植株的高代品系,其GBSS酶活性与直链淀粉含量均极低[16]。本研究结果与前人的研究一致,GBSS酶活性与直链淀粉合成密切相关,与直链淀粉合成密切相关,与直链淀粉积累速率显著相关,且直链淀粉含量较高的青稞GBSS酶活性较高。
王月福等认为,如果SSS酶活性越高,利用底物ADPG合成支链淀粉的能力就越强[17]。本试验中支链淀粉含量高的甘垦5号其SSS活性也较高,相关性分析表明, SSS的酶活性变化与支链淀粉积累速率呈极显著正相关,同时AGPP和SBE的酶活性变化也同样与支链淀粉积累速率呈显著正相关,从而可以推断,支链淀粉的合成可能是由AGPP、SSS、SBE 3个酶协同完成的,而SSS酶是支链淀粉合成的关键酶。
谷类作物籽粒中的支链淀粉是在SBE与SSS共同作用下形成的,并且对支链淀粉结构形成起重要作用[18]。本试验研究发现,青稞籽粒灌浆期SBE活性与直链淀粉和支链淀粉积累速率均呈显著正相关,这说明SBE不仅参与支链淀粉的合成,也参与直链淀粉的合成。且SBE的活性值接近或滞后于SSS活性峰值,由此推断,SSS酶活性在最终支链淀粉积累量的过程中起着决定性的作用,而SBE主要参与支链淀粉精细结构的形成。
谷类中α-淀粉酶与β-淀粉酶在籽粒灌浆期的重要性已经有比较普遍的阐述[19]。本试验研究发现,灌浆期总淀粉积累速率、直链淀粉和支链淀粉积累速率与3个青稞品种的α-淀粉酶呈负相关,与3个青稞品种的β-淀粉酶呈显著或极显著负相关。α-淀粉酶与β-淀粉酶均为淀粉降解酶,说明β-淀粉酶活性对灌浆期总淀粉积累速率、直链淀粉和支链淀粉积累速率的调节作用较α-淀粉酶的调节作用强。
灌浆初期淀粉合成酶(AGPP、GBSS、SSS、SBE)活性较小,而淀粉降解酶(α-淀粉酶、β-淀粉酶)活性较强,淀粉合成能力较弱,合成的淀粉部分被α-淀粉酶和β-淀粉酶分解为葡萄糖、麦芽糖、α-1,6糖苷键的糊精和极限糊精,淀粉的合成速率较低,淀粉的积累量相对较少;随着籽粒的灌浆进行,淀粉合成酶AGPP、GBSS、SSS和SBE活性均逐渐增大,淀粉合成速率也随之上升,从而使得淀粉快速积累,淀粉分解酶α-淀粉酶和β-淀粉酶分解方向活性缓慢下降,籽粒中淀粉的合成速率大于分解速率,淀粉整体合成速率加快,导致淀粉大量积累;乳熟期以后,α-淀粉酶和β-淀粉酶分解方向活性有所回升,AGPP、GBSS、SSS和SBE合成方向活性下降,籽粒内的合成速率小于分解速率,淀粉整体合成速率减慢,但淀粉继续积累。青稞籽粒灌浆过程中淀粉含量积累是由于淀粉合成酶活性的增强,而不是分解酶活性的减弱。因此,青稞籽粒中淀粉积累是淀粉代谢酶(合成和分解方向)综合作用的结果。
参考文献:
[1]邹弈星,潘志芬,邓光兵,等.青藏高原青稞的淀粉特性[J].麦类作物学报,2008,28(1):74-79.
[2]曹颖妮.小麦籽粒灌浆期淀粉合成酶学机制及基因表达模式的研究[D]:西北农林科技大学,2012.
[3]王自布,李卫华,齐军仓,等.小麦籽粒胚乳淀粉合成酶基因表达及酶活性分析[J].核农学报,2010,24(6):1 117-1 123.
[4]Radchuk VV,Borisjuk L,Sreenivasulu N,et al.Spatiotemporal profiling of starch biosynthesis and degradation in the developing barley grain[J].Plant physiology,2009,150(1):190-204.
[5]张海萍,阎长生.小麦迟熟α-淀粉酶的研究进展[J].麦类作物学报,2003,23(1):81-85.
[6]Zanella M,Borghi GL,Pirone C,et al.β-amylase 1(BAM 1) degrades transitory starch to sustain proline biosynthesis during drought stress[J].Journal of experimental botany,2016,67(6):1819-1826.
[7]赵俊晔,于振文.小麦籽粒蛋白质和淀粉代谢及其与品质形成关系的研究进展[J].麦类作物学报,2005,25(3):106-111.
[8]张文会,强小林.青藏区主要青稞品种淀粉理化特性分析[J].食品研究与开发,2014,35(4):14-18.
[9]李润喜,李学才,毋玲玲.黑裸大麦甘垦5号的营养功能特性及利用[J].甘肃科技,2012,28(18):169-170.
[10]程方民,蒋德安.早籼稻籽粒灌浆过程中淀粉合成酶的变化及温度效应特征[J].作物学报,2001,27(2):201-206.
[11]Nakamura Y,Yuki K,Park S-Y,et al.Carbohydrate metabolism in the developing endosperm of rice grains[J].Plant and Cell Physiology,1989,30(6):833-839.
[12]李太贵,沈波.Q酶在水稻籽粒垩白形成中作用的研究[J].作物学报,1997,23(3):338-344.
[13]何照花.谷物中直.支链及总淀粉双波长测定法[J].生物化学与生物物理进展,1981(1):70-72.
[14]李世清,邵明安,李紫燕,等.小麦籽粒灌浆特征及影响因素的研究进展[J].西北植物学报,2004,23(11):2 031-2 039.
[15]刘霞,姜春明,郑泽荣,等.藁城8901和山农1391淀粉合成酶活性和淀粉组分积累特征的比较[J].中国农业科学,2005,38(5):897-903.
[16]邓万洪,晏本菊,任正隆.糯性小麦研究进展[J].麦类作物学报,2007(1):166-171.
[17]王月福,于振文.小麦籽粒灌浆过程中有关淀粉合成酶的活性及其效应[J].作物学报,2003,29(1):75-81.
[18]Smith AM,Denyer K,Martin CR.What controls the amount and structure of starch in storage organs[J].Plant physiology,1995,107(3):673.
[19]Barrero JM,Mrva K,Talbot MJ,et al.Genetic,hormonal,and physiological analysis of late maturity α-amylase in wheat[J].Plant physiology,2013,161(3):1 265-1 277.
