胸腺肽α1诱导MUC1特异性Th2型免疫应答
2018-03-30孙敏英周红月翟瑞萍陈潭秀袁红艳台桂香
孙敏英,周红月,接 晶,谢 飞,翟瑞萍,陈潭秀,袁红艳,台桂香
(1.吉林大学基础医学院医学生物学实验中心,吉林 长春 130021;2.吉林大学基础医学院免疫学教研室,吉林 长春 130021)
胸腺肽α1(thmosin α1,Tα1)由28个氨基酸多肽组成,具有高度保守酸多肽结构。Tα1主要分布于胸腺组织,也存在于淋巴组织(如脾、淋巴结)和非淋巴组织(如肺、肾和脑)[1],其分泌和释放并不受其他激素或者释放因子所调节[2]。尽管Tα1确切的免疫机制还不清楚,但是实验[3]证实:Tα1具有强大的免疫调节作用,是一种细胞免疫增强剂和生物反应调节因子,在机体免疫应答过程中十分重要。大量的实验数据[4-7]证实:Tα1不仅自身具有延缓肿瘤复发、改善免疫功能的作用,在与其他抗肿瘤药物联合使用时还可提高癌症治疗效果。MUC1是Mucins 黏蛋白家族成员,存在于正常腺管上皮细胞及其来源的肿瘤细胞表面,由多肽核芯(核芯肽)和侧枝糖链构成,其核芯肽胞外段为20个氨基酸(SAPD TRPAP GSTAPPA HGVT) 的串联重复序列(variable numbers tandem repeats,VNTRs)。正常组织的 MUC1 与肿瘤组织不同,前者分布于腺管上皮细胞分泌极,与免疫细胞相对隔离,糖基化丰富;而后者广泛分布并异常丰富地表达于癌细胞表面,糖基化不完全,因此暴露出正常情况下隐蔽的表位,成为免疫细胞攻击的靶点[8-9],是制备肿瘤疫苗的理想分子。目前研制的以MUC1为靶点的疫苗包括糖疫苗、蛋白或多肽疫苗和DNA疫苗[10-12],其中蛋白疫苗具有一定的临床应用前景。本课题组前期研究[13-14]以MUC1为靶点,研制黏蛋白与麦芽糖结合蛋白融合蛋白(mucin1-maltose binding protein fusion protein,MUC1-MBP)的抗肿瘤疫苗,表明MUC1-MBP联合卡介苗(bacillus calmette-guérin,BCG)佐剂在小鼠体内可以诱导细胞毒性T淋巴细胞(cytotoxic T lymphocyte,CTL) 细胞毒的杀伤活性和Th1反应,可以抑制肺癌转移及实体瘤的生长,具有明显的肿瘤治疗作用[13-16]。但由于BCG是减毒活疫苗,而且其活菌数较难控制,导致疫苗稳定性差,且高剂量BCG的不良反应较大,因此探索良好的肿瘤疫苗佐剂一直是本课题组的重要研究课题。由于Tα1具有免疫调节作用,本研究探讨Tα1是否能作为MUC1-MBP的佐剂用于抗肿瘤治疗,旨在为开发新的佐剂提供依据。
1 材料与方法
1.1实验动物、主要试剂和仪器67只雌性C57BL/6小鼠,体质量为18~20 g,6~8周龄,动物合格证号:SCXK(京)2014-0004,购自北京阜康生物科技股份有限公司。小鼠黑色素瘤细胞B16-MUC1由本室建立并保存,重组MUC1-MBP和MUC1-GST融合蛋白由本室重组构建及纯化[6]。BCG购自石家庄开创生化公司,Tα1购自成都地奥九泓制药厂,伊思柯夫改良培养液(Iscove’s modified Dubecco’s medium,IMDM)和 G418 购自美国 Gibco公司,FBS和小鼠淋巴细胞分层液购自天津市灏洋生物制品有限公司,ConA购自美国Sigma公司,小鼠抗人 MUC1购自美国Abcam公司,FITC-山羊抗小鼠 IgG和山羊抗鼠 IgG-HRP 购自北京中杉金桥生物技术有限公司,鼠抗MBP 多克隆抗体购自基因公司,大鼠抗鼠 IgG1-HRP 和山羊抗鼠 IgG2a-HRP购自美国Southern Biotech公司,小鼠细胞因子干扰素γ(IFN-γ)、白细胞介素2(IL-2)、白细胞介素4(IL-4)和白细胞介素10(IL-10) ELISA 检测试剂盒购自美国eBioscience公司。低温冰箱和CO2孵箱购自日本SANYO公司,酶标仪购自美国BIOTEK公司,紫外分光光度计购自日本岛津公司。
1.2小鼠分组与免疫方案①T细胞免疫活性检测组。将21只C57BL/6小鼠随机分成3组,每组7只,分别为生理盐水组(注射生理盐水)、MUC1-MBP+ BCG组(注射MUC1-MBP+BCG)和MUC1-MBP+ Tα1组(注射MUC1-MBP+Tα1),其中MUC1-MBP+ BCG组为阳性对照组。 免疫方案:小鼠颈背部皮下多点注射,MUC1-MBP 50 μg/只,BCG 作为佐剂1 mg/只,Tα1为30 μg/只,隔周1 次,共注射3次。②MUC1特异性抗体检测组:将25只C57BL/6小鼠随机分成5组,每组5只,分别为生理盐水组(注射生理盐水)、MUC1-MBP 50 μg组、MUC1-MBP 50 μg+ Tα1 15 μg组、MUC1-MBP 50 μg+ Tα1 30 μg 组和MUC1-MBP 50 μg+ Tα1 60 μg组,各组分别用生理盐水调至400 μL。免疫方案:小鼠颈背部皮下多点注射及两侧腹股沟皮下注射,第1次免疫后2周,同等剂量加强免疫2次。③肿瘤模型组。将21只C57BL/6小鼠随机分成3组,每组7只,分别为生理盐水组(注射生理盐水)、MUC1-MBP+BCG组(注射MUC1-MBP+BCG)和MUC1-MBP+ Tα1组(注射MUC1-MBP+Tα1),其中MUC1-MBP+BCG组为阳性对照组。免疫方案:小鼠颈背部皮下多点注射,MUC1-MBP 50 μg/只,BCG作为佐剂1 mg/只, Tα1为30 μg/只,隔周1次,共注射3次。
1.3脾指数测定第3次免疫后第4~7天,先称每只小鼠体质量,杀鼠,无菌取脾脏,称脾脏质量,计算脾指数。脾指数 = 小鼠脾质量/体质量。
1.4 检测各组小鼠的刺激指数(stimulus index,SI)和特异性SI在第3 次免疫后第4~7 天杀鼠,无菌取脾脏、研磨、细胞计数,制备脾细胞悬液,采用小鼠淋巴细胞分层液分离小鼠脾脏单个核细胞,采用IMDM调整细胞浓度至1×107mL-1每孔100 μL加入96孔板中。非特异性淋巴细胞增殖反应:实验分为阴性对照组和ConA刺激组(终浓度5 mg·L-1),每组设3复孔,37℃的 CO2培养箱中培养48 h后,加入WST-1检测试剂每孔10 μL,继续反应1 h,通过酶标仪检测450 nm处吸光度(A)值。MUC1特异淋巴细胞增殖反应:实验分为阴性对照组、IL-2单独刺激组(终浓度100 U·mL-1)和MUC1多肽刺激组(MUC1多肽20 mg·L-1+IL-2 100 U·mL-1)。37℃ 、CO2培养箱3 d,半量换液,继续培养5 d后,吸取每孔细胞培养上清100 μL用于细胞因子的检测,加入WST-1检测试剂每孔10 μL,继续反应1 h,通过酶标仪检测450 nm波长处测量吸光度(A)值,计算SI。SI= 实验孔A值/对照孔A值。
1.5 ELISA法检测各组小鼠脾细胞培养上清中细胞因子IFN-γ、IL-2、IL-4和IL-10水平收集ConA刺激2 d和MUC1刺激5 d的淋巴细胞培养上清,按照IFN-γ、IL-2、IL-4和IL-10试剂盒说明书进行操作。包被抗体(抗IFN-γ、IL-2、IL-4和IL-10的抗体)包被96孔酶标板,每个样本设双复孔,4℃过夜。采用PBS-0.05%Tween-20洗板5次,加入封闭液封闭1 h后,洗涤2次,加入标准品(IFN-γ:2 000.000、1 000.000、500.000、250.000、125.000、62.500、32.500和15.625 ng·L-1;IL-2标准品配制:1 200、600、300、150和75 ng·L-1;IL-4 标准品配制:500.0、250.0、125.0、62.5、32.5、15.6、7.8和3.9 ng·L-1;IL-10标准品配制:200.0、100.0、50.0、 25.0和12.5 ng·L-1)和样品,每孔100 μL,4℃过夜。洗涤5次,加入检测抗体室温孵育1 h,洗涤5次,加入酶标抗体Avindin-HRP,室温孵育30 min。洗7次,加入TMB,室温15 min。加入硫酸终止后,采用酶标仪于450 nm波长测定A值。计算IFN-γ、IL-2、IL-4和IL-10水平。
1.6ELISA法检测MUC1总抗体效价及亚类第3次免疫后第4 天,眶上静脉采血,分离血清,通过间接ELISA试剂盒检测MUC1特异性抗体IgG、IgG1和IgG2a。首先将包被抗原MUC1-GST包被96孔板,每个样本设双复孔,4℃过夜。采用PBS-0.05%Tween-20洗板3次,加入封闭液封闭1.5 h后,洗涤2次,加一抗(待测血清)不同用药组每只小鼠血清倍比稀释(1∶500、1∶1 000、1∶2 000、1∶4 000、1∶8 000、1∶16 000、1∶32 000、1∶64 000),每孔100 μL,均设双复孔,37℃孵育1.5 h。洗涤6次,加酶标二抗HRP(辣根过氧化物酶)(山羊抗小鼠 IgG,1∶10 000;山羊抗小鼠 IgG1,1∶4 000;山羊抗小鼠 IgG2a,1∶1 000),每孔100 μL,37℃孵育1.5 h。洗涤7次,加入TMB,室温15 min。加硫酸终止后,采用酶标仪于450 nm波长处测定A值。
1.7肿瘤预防实验模型构建第3次免疫后1周,在小鼠右后背胁处接种黑色素瘤细胞B16-MUC1,每只2.0 × 106个。隔天称体质量,用游标卡尺测量肿瘤的最大直径(a)和最小半径(b),计算肿瘤体积(V):V=1/2ab2,直至第23天第1只小鼠死亡,观察小鼠生存期。
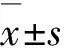
2 结 果
2.1各组小鼠脾指数和淋巴细胞增殖反应MUC1-MBP+ BCG组和MUC1-MBP+ Tα1组小鼠脾指数明显高于生理盐水组(P< 0.01)。非特异性淋巴细胞增殖反应:MUC1-MBP+ BCG组和MUC1-MBP+ Tα1组SI均高于生理盐水组(P< 0.05),以MUC1-MBP+BCG组升高最为明显。MUC1特异性淋巴细胞增殖反应:MUC1-MBP+BCG组和MUC1-MBP+Tα1组特异性SI均高于生理盐水组(P< 0.05)。见表1。
表1各组小鼠脾指数、SI和特异性SI


GroupSpleenindexSISpecificSINormalsaline0.0039±0.00071.4570±0.35991.4290±0.1799MUC1⁃MBP+BCG0.0052±0.0017∗∗2.2860±0.2249∗1.9570±0.3599∗MUC1⁃MBP+Tα10.0061±0.0008∗∗1.9140±0.4981∗1.8430±0.2699∗
*P< 0.05,**P< 0.01vsnormal saline group.
2.2各组小鼠脾细胞培养上清中IFN-γ、IL-2、IL-4和IL-10水平与生理盐水组比较,MUC1-MBP+BCG组小鼠脾细胞培养上清中IFN-γ和IL-2水平明显升高(P<0.05);IL-4和IL-10水平略升高,但差异无统计学意义(P>0.05)。与生理盐水组比较,MUC1-MBP+Tα1组小鼠脾细胞培养上清中 IL-4水平明显升高(P<0.01);IFN-γ、IL-2、IL-4和IL-10水平升高,但差异无统计学意义(P>0.05)。见表2。
表2各组小鼠脾细胞培养上清中IFN-γ、IL-2、IL-4和IL-10水平
Tab.2Levels of IFN-γ,IL-2,IL-4 and IL-10 in culture supernatant of spleen cells of mice in various groups

GroupIFN⁃γIL⁃4IL⁃2IL⁃10Normalsaline152.90±49.4947.14±13.50145.00±44.9142.86±13.50MUC1⁃MBP+BCG471.40±135.00∗72.86±17.99471.40±179.90∗53.57±8.99MUC1⁃MBP+Tα1200.00±95.74113.10±44.99∗∗185.70±89.9757.14±17.99
*P< 0.05,**P< 0.01vsnormal saline group.
2.3各组小鼠抗MUC1特异性抗体效价随着Tα1剂量的增加,相应的抗体效价也有所提高,并且高剂量的Tα1能促进抗体的产生。见表3。抗体亚类检测结果显示:Tα1能够明显诱导IgG1的产生,并且有剂量依赖关系,而IgG2a产生不明显。见表4。
2.4各组小鼠生存期注射肿瘤细胞1周之内肿瘤生长缓慢,第8 天生理盐水组最先触及肿瘤结节,第11 天 MUC1-MBP+Tα1组触及肿瘤结节,第14 天MUC1-MBP+BCG组触及肿瘤结节。生理盐水组小鼠生存期为 (26±4) d,MUC1-MBP+BCG组小鼠生存期为(28.7±6.3) d,MUC1-MBP+ Tα1组小鼠生存期为(28±6) d。见图1。MUC1-MBP+BCG组和MUC1-MBP+Tα1组与生理盐水组小鼠生存期比较差异均无统计学意义(P>0.05)。
表3ELISA法检测免疫小鼠抗MUC1特异性抗体效价
Tab.3Anti-MUC1 specific antibody titers in serum of immunized mice detected by ELISA

Antigen(MUC1⁃MBP)(50μg) Tα1(μg)Titerofantibody --0 +- 1∶500-1∶1000 +15 1∶500-1∶2000 +301∶1000-1∶4000 +601∶4000-1∶8000
表4间接ELISA法检测MUC1特异性抗体亚型
Tab.4Subtypes of MUC1 specific antibodies detected by indirect ELISA


GroupIgG1IgG2aIgG1/IgG2aA0.0484±0.00040.0498±0.00100.9631±0.0205B0.9871±0.0008∗0.0481±0.000820.6300±0.4359C3.6320±0.0256∗∗0.0515±0.001972.4700±0.6094D3.7980±0.0800∗∗0.0575±0.000466.6000±2.2710E1.8380±0.1255∗∗0.6089±0.00712.9460±0.2863
*P< 0.05,**P< 0.01vsA group.A:Control; B:MUC1-MBP (50 μg); C:MUC1-MBP (50 μg)+ Tα1(5 μg);D:MUC1-MBP (50 μg)+ Tα1(130 μg);E:MUC1-MBP (50 μg)+ Tα1 (60 μg).
3 讨 论
本文作者将BCG与MUC1-MBP联合作为阳性对照组,并与Tα1联合MUC1-MBP免疫C57BL/6小鼠进行对比。本研究免疫活性检测结果显示:MUC1-MBP+ BCG组和MUC1-MBP+Tα1组小鼠脾指数、有丝分裂原SI、MUC1多肽特异性SI均有所升高,说明MUC1-MBP联合Tα1和联合BCG均可诱导非特异性淋巴细胞增殖和特异性淋巴细胞增殖。由于CTL杀伤活性离不开Th1的辅助,因此,本文作者分析了诱导活化的Th类型。采用MUC1多肽刺激5 d,检测了脾脏特异性T淋巴细胞分泌的细胞因子IFN-γ、IL-2、IL-4和IL-10水平,结果显示:MUC1-MBP联合Tα1明显提高了IL-4和IL-10的分泌,而联合BCG提高了IFN-γ和IL-2的分泌水平,提示Tα1诱导MUC1特异的Th2型应答,而BCG却诱导MUC1特异的Th1应答,该结果与本文作者前期的研究[15-18]结果一致。抗体的产生提示体液免疫应答的激活,也提示Th2的活化。而且,IgG1在IL-4诱导下才能产生,IgG2a在IFN-γ诱导下产生,因此检测IgG1和IgG2a能间接提示Th2和Th1应答。本文作者进一步通过ELISA法检测对免疫小鼠的血清IgG和亚类IgG1和IgG2a进行分析,结果发现:当免疫3次后,与MUC1-MBP组比较,MUC1-MBP+ Tα1组产生了较高效价的MUC1特异性IgG,并随着Tα1剂量增加而升高,提示Tα1在促进MUC1-MBP产生体液免疫应答中有佐剂作用。进一步检测抗体亚类结果发现:随着Tα1剂量的增加,IgG1抗体的产生能力逐渐增强,而IgG2a未见产生,进一步提示Tα1诱导MUC1特异性Th2应答。

A:Tumor growth situation; B: Survival time curve.
Fig.1Tumor growth and survival curves of tumor-bearing mice in various groups 23 d after inoculation
Th1应答不仅辅助CTL的杀伤,而其本身释放的IL-2、IFN-γ和TNF-α在治疗肿瘤中发挥重要的作用,Th2应答主要辅助体液免疫应答产生抗体,抗体在肿瘤治疗早期发挥作用,到晚期不仅发挥不了抗肿瘤作用,可能还会起到相反的作用[19-21]。因此,本文作者采用荷瘤鼠模型,深入探讨了Tα1联合MUC1-MBP抗肿瘤作用。本文作者将小鼠免疫3次后,给小鼠注射稳定转染MUC1的黑色素瘤细胞B16-MUC1,结果显示:Tα1联合MUC1-MBP作用下肿瘤生长在早期较对照组缓慢,而到后期与对照组无明显差别,未明显延长小鼠生存期,与本文作者的预期一致。而MUC1-MBP+BCG组作为阳性对照组也未见明显延长生存期,考虑可能是BCG活疫苗本身不稳定导致,其诱导产生的Th1应答强度较低,不足以诱导有效的抗肿瘤应答。
综上所述,Tα1促进MUC1-MBP诱导产生特异性Th2应答,明显促进体液免疫应答的产生,具有佐剂作用。Th2型应答的佐剂不适合应用于治疗性疫苗,但适合用于预防性疫苗。本研究结果也提示在临床中使用Tα1应该慎重,盲目应用有可能产生相反的作用。
[参考文献]
[1] GoldsteinAL.From lab to bedside: emerging clinical applications of thymosin alpha-1[J].Expert Opin Biol Ther,2009,9(5):593-608.
[2] Naylor PH,Quadrini K,Garaci E,et al.Immunopharmacology of thymosin alpha 1 and cytokine synergy[J].Ann NY Acad Sci,2007,1112: 235-244.
[3] Romani L,Moretti S,Fallarino F,et al.Jack of all trades: thymosin α1 and its pleiotropy [J].Ann NY Acad Sci,2012,1269(1):1-6.
[4] Schulof RS,Lloyd MJ,Cleary PA,et al.A randomized trial to evaluate the immunorestorative properties of synthetic thymosin-alpha 1 in patients with lung cancer[J].J Biol Response Mod,1985,4(2):147-158.
[5] Maio M,Mackiewicz A,Testori A,et al.Large randomized study of thymosin alpha 1 interferon alfa or both in combination with dacarbazine in patients with metastatic melanoma[J].J Clin Oncol,2010,28:1780-1787.
[6] Stefanini GF,Foschi FG,Castelli E,et al.Alpha-1 thymosin and transcatheter arterial chenoembolization in hepatocellular carinoma patients-a preliminary experience[J].Hepatogastroenterology,1998,45(19):209-215.
[7] Cheng SQ,Wu MC,Chen H,et al.Anti-viral therapy using lamivudine and thymosin is helpful to prevent recurrence in hepatocellular carcinoma with coexisting active hepatitis B[J].Zhonghua Zhong Liu Za Zhi,2005,27(2):114-116.
[8] Acres B,Limacher JM.MUC1 as a target antigen for cancer immunotherapy[J].Expert Rev Vaccines,2005,4(4):493-502.
[9] Nath S,Mukherjee P.MUC1:a multifaceted oncoprotein with a key role in cancerprogression[J].Trends Mol Med,2014,20(6):332-342.
[10]Vassilaros S,Tsibanis A,Tsikkinis A,et al.Up to 15-year clinical follow-up of a pilot Phase Ⅲ immunotherapy study in stage Ⅱ breast cancer patients using oxidized mannan-MUC1[J].Immunotherapy,2013,5(11):1177-1182.
[11]Quoix E,Ramlau R,Westeel V,et al.Therapeutic vaccination with TG4010 and first-line chemotherapy in advanced non-small-cell lung cancer: a controlled phase 2B trial [J].Lancet Oncol,2011,12(12): 1125-1133.
[12]Wurz GT,Kao CJ,Wolf M,et al.Tecemotide:Anantigen-specific cancer immunotherapy[J].Hum Vaccin Immunother,2014,10(11):3383-3393.
[13]Hu B,Wang J,Guo Y,et al.Pre-clinical toxicity and immunogenicity evaluation of a MUC1-MBP/BCG anti-tumor vaccine[J].Int Immunopharmacol,2016,33:108-118.
[14]Wang F,Ni W,Liu G,et al.Escherichiacolimaltose-binding protein (MBP) directly induces mouseTh1activation through upregulating TLR2 and downregulating TLR4expressions [J].Immunobiology,2015,220(6):782-788.
[15]台桂香,张凤吉,朱迅.重组人MUC1-MBP融合蛋白的抗肿瘤作用[J].中国肿瘤生物治疗杂志,2003,10(3):202-205.
[16]Fang F,Ma J,Ni W,et al.MUC1 and maltosebinding protein recombinant fusion protein combined with Bacillus Calmette-Guerin induces MUC1-specific and nonspecific anti-tumor immunity in mice[J].Mol Med Rep,2014,10(2):1056-1064.
[17]赵小霞,马吉春,方芳,等.大肠杆菌麦芽糖结合蛋白(MBP)诱导小鼠Th1细胞的活化作用[J].中国免疫学杂志,2009,25(6):504-507.
[18]Ni W,Wang F,Liu G,et al.TLR9 played a more important role than TLR2 in the combination ofmaltose-bindingprotein and BCG-induced Th1 activation[J].Mol Immunol,2016,79:32-37.
[19]Li Q,Zhang H,Yu L,et al.Down-regulation of Notch signaling pathway reverses the Th1/Th2 imbalance in tuberculosis patients[J].Int Immunopharmacol,2018, 54:24-32.
[20]Li L,Jing YB,Liu J,et al.Study on the correlation of the effect of entecavir on Th1/Th2 cytokines level in the treatment of chronic hepatitis[J].Zhonghua Gan Zang Bing Za Zhi,2017,25(8):606-611.
[21]Kaminuma O,Saeki M,Nishimura T,et al.Differential contribution of adhesion molecules to Th1 and Th2 cell-mediated lung and bowel inflammation[J].Biol Pharm Bull,2017,40(10):1801-1805.
