西南部分地区圈养非人灵长类贾第虫感染情况调查及多位点基因序列分析
2018-03-21田一男黄祥明但佳明杨亭玉彭广能钟志军
田一男,魏 斌,李 平,黄祥明,李 威,涂 蕊,但佳明,袁 莉,杨亭玉,彭广能,钟志军,*
(1.四川农业大学 动物医学院,动物疫病与人类健康四川省重点实验室,四川 成都 611130; 2.邛崃市农业和林业局,四川 邛崃 611530; 3.成都大熊猫繁育研究基地 四川省濒危野生动物保护生物学省部共建实验室,四川 成都 610081)
贾第虫(Giardiaduodenalis)是重要的人兽共患性寄生虫之一,可感染人、家畜和多种野生动物[1]。人与动物感染贾第虫后表现的症状多样,包括无明显临床表现或腹泻、脱水、腹痛、呕吐、体重减轻等急性或慢性症状[2],其轻重程度取决于动物的营养、免疫力以及寄生虫的毒力等因素。常见的感染方式包括水、食物、粪便等经口感染,人与动物接触以及动物与动物间接触等,其中多次报道由于水源污染而引发贾第虫病暴发[4-5]。
采用分子分型技术已鉴定出8个(A-H)贾第虫集聚体,其中A和B为最常见的人兽共患集聚体,C和D可感染犬、狼等动物,E主要感染牛、羊等家畜,F常感染猫,G和H分别在啮齿类和海豹等海洋生物中发现,非人灵长类中主要感染集聚体A和B[6]。
非人灵长类动物作为珍贵野生动物资源以及重要的实验动物,与人类直接或间接接触的机会较多,其中圈养非人灵长类与人接触的概率更大。本研究选择bg、tpi和gdh基因,通过巢式PCR扩增基因序列并构建系统发育树,分析西南部分地区圈养非人灵长类贾第虫感染情况及其基因型,对保护珍贵野生动物以及人类公共健康具有重要意义。
1 材料与方法
1.1 主要试剂
粪便DNA提取采用QIAGEN公司的PowerSoil®DNA试剂盒;2×TaqPCR Master Mix购自北京天根生化科技有限公司。引物由英潍捷基(上海)贸易有限公司合成。
1.2 样品采集
于2016年3月至5月以及2016年9月至11月期间,在西南部分地区国家猕猴繁殖实验基地、贵阳动物园、雅安碧峰峡动物园、成都市高新区猕猴养殖场、成都动物园等采集非人灵长类新鲜粪便共207份,包括猕猴粪便101份、长臂猿粪便36份、金丝猴粪便9份、食蟹猴粪便60份。每份样品约50 g,分别用洁净自封袋装好、标记,样品置于4 ℃冰箱保存待检。
1.3 DNA提取及PCR扩增测序
采用改良饱和蔗糖溶液漂浮法处理207份非人灵长类粪便,按照PowerSoil®DNA提取试剂盒说明书操作提取DNA,-20 ℃保存备用。
bg、tpi和gdh基因引物设计及扩增条件参照Sulaiman等[7]、Appelbee等[8]和Caccio等[9]的报道。反应体系为25 μL∶2×TaqPCR Master Mix 12.5 μL、Deionized Water 7.5 μL、0.1%BSA 1.0 μL、上下游引物各1.0 μL、模板DNA 2.0 μL。取6 μL扩增产物于1%琼脂糖凝胶孔中,在100 V电压下电泳1 h,然后于凝胶成像系统仪中拍照,扩增阳性产物送华大基因公司测序。测序结果在GenBank中BLAST比对校正后提交至NCBI。将校正后序列与NCBI上参考序列使用Clustal X比对,然后使用Mega 6的NJ法构建系统发育树。
2 结果
2.1 西南部分地区圈养非人灵长类贾第虫感染情况
PCR扩增结果显示,207份样品中16份样品为贾第虫感染阳性(感染率为7.73%)。其中猕猴、长臂猿、食蟹猴的感染率分别为0.99%(1/101),38.89%(14/36),1.67%(1/60),金丝猴未检出贾第虫感染。5个采样点中仅2个采样点检出贾第虫,分别为贵阳动物园,感染率为30.0%(15/50),成都市高新区猕猴养殖场,感染率为1.67%(1/60)。从本次检测的省份来看,四川地区非人灵长类贾第虫感染率为0.64%(1/157),贵阳地区非人灵长类贾第虫感染率为30.0%(15/50)。所有阳性样品均完成bg、tpi和gdh基因位点扩增,经鉴定,均为集聚体B。将序列提交至NCBI获得登录号分别为KY696790-KY696821。
2.2 基因序列分析
在NCBI中搜索bg、tpi和gdh相关参考序列,用Mega 6与本次研究所得序列进行同源性分析。基于gdh基因位点,16条序列均鉴定为BIV亚型,其中1条与巴西人贾第虫分离株(GenBank:EF507672)完全一致,其余15条与巴西人贾第虫分离株(GenBank:EF507682)完全一致。
基于bg基因位点(图1),16条序列形成5个基因亚型(GenBank:KY696824、KY696825、KY696828、KY696832、KY696836),包括已知基因亚型的环尾狐猴分离株Bb-4(GenBank:KJ888977)、人分离株BIII(GenBank:KF922976)、巴巴利猕猴分离株BIII-1(GenBank:EU637581)、猕猴分离株B1(GenBank:KM211793),以及一个新基因亚型,命名为BIII-2。其中以BIII(n=7)基因亚型为主。
基于tpi基因位点(图2),16条序列均鉴定为集聚体B,形成6个亚型,包括已知基因亚型水源分离株B7(GenBank:JQ863259)、山魈分离株WB6(GenBank:KJ888987)、猕猴分离株WB8(GenBank:KF679738)、人分离株EB5(GenBank:KT948110)、人分离株BIV(GenBank:AB18783),以及一个新基因亚型B9(GenBank:KY696810)。其中WB8(n=7)是最主要的基因亚型。
3 讨论
本次调查,西南部分地区非人灵长类贾第虫总感染率为7.73%,高于广西、秦岭以及全国范围内非人灵长类贾第虫感染率(2.4%,2.0%,2.2%和1.3%)[10-13]。本试验中,四川地区非人灵长类贾第虫感染率为0.64%,与Li等[11]报道的四川地区非人灵长类贾第虫感染率(0.5%)一致。而贵州地区的感染率(30.0%)远高于之前Ye等[14]报道贵州地区感染率(8.5%)。总体而言,本次试验中四川地区非人灵长类感染率低于贵州地区非人灵长类贾第虫感染率(30%),也低于北京(13.89%)、上海(2.34%)、河北(0.98%)、河南(2.73%)等地的贾第虫感染率。但四川地区非人灵长类感染率高于广东(0.3%)地区[11]。本试验采集的5个动物园样品中,贵阳动物园检测出阳性率为30.0%,成都市高新区猕猴养殖场检测出阳性率为1.67%,其他动物园未检出贾第虫感染。贵阳动物园的非人灵长类贾第虫感染率仅低于长沙野生动物园感染率(44.0%),高于石家庄动物园(11.2%)、武汉动物园(7.6%)、太原动物园(13.6)、北京动物园(22.2%)、上海动物园(8.2%)、上海野生动物园(20.9%)等[15]。不同动物园出现的感染率差异可能与各地饲养密度、管理差异以及贾第虫的单宿主生活史等相关[16]。
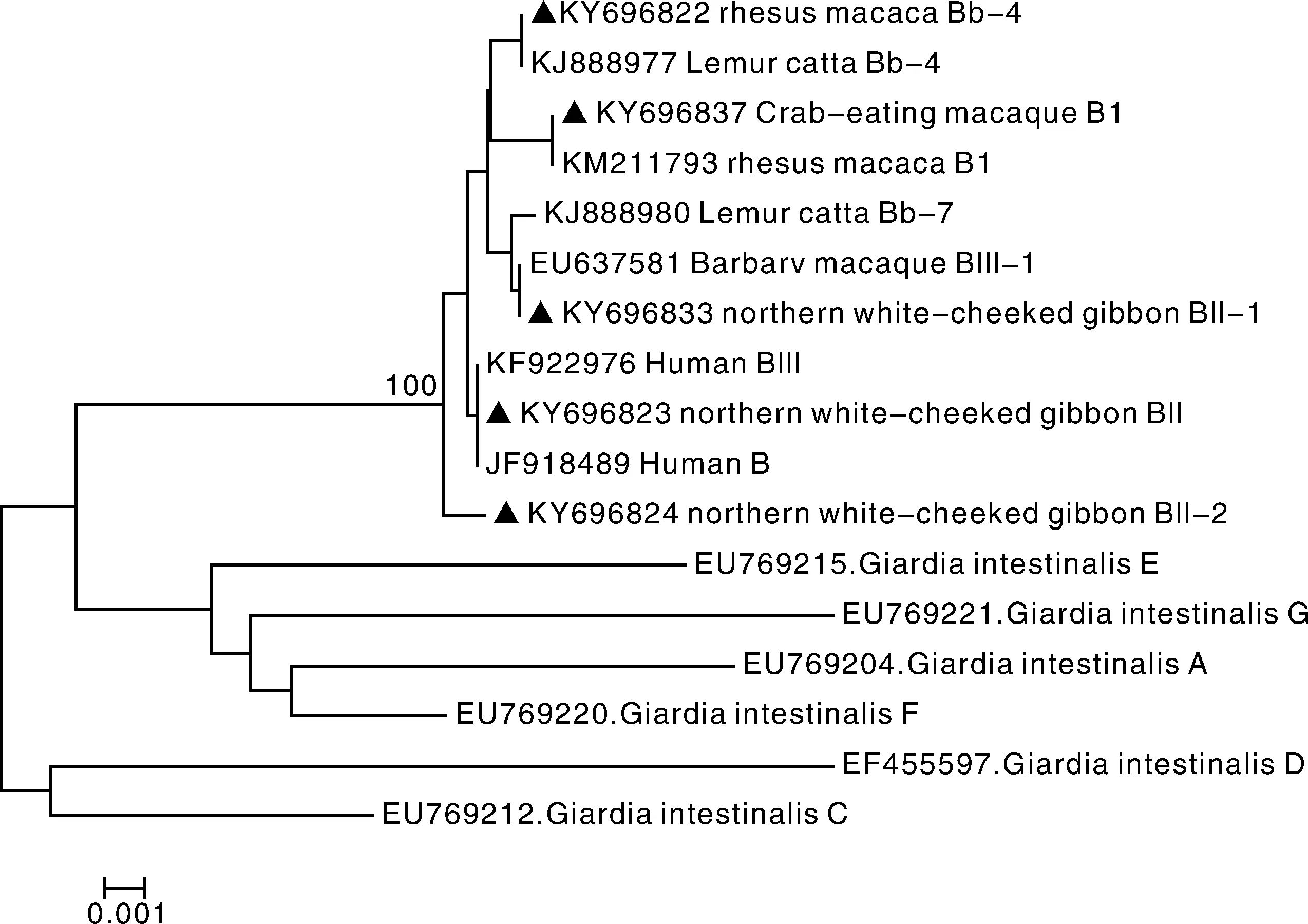
▲,本实验所得序列▲ Represents the sequence obtained from this experiment图1 非人灵长类贾第虫bg基因位点系统发育树Fig.1 Phylogenetic tree based on bg gene sequence of G. duodenalis in non-human primates
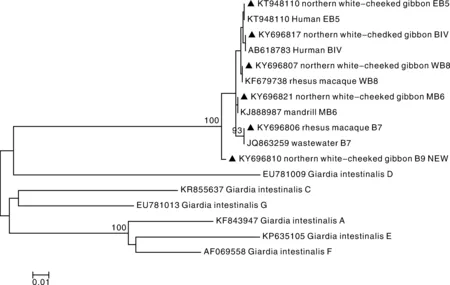
▲,本实验所得序列▲ Represents the sequence obtained from this experiment图2 非人灵长类贾第虫tpi基因位点系统发育树Fig.2 Phylogenetic tree based on tpi gene sequence of Giardia duodenalis in non-human primates
贾第虫(G.duodenalis)是一种可感染人、家畜和多种野生动物的人兽共患寄生虫。非人灵长类中已发现贾第虫集聚体A、B和E,其中集聚体B是感染非人灵长类的主要集聚体[1,13,17],也是感染人的主要集聚体之一[6]。此次调查采用β-giardin、tpi和gdh等多位点基因序列分析显示,西南部分地区贾第虫阳性样品均为集聚体B,与先前报道的结论一致。序列分析结果显示gdh基因位点较保守,所有样本均鉴定为BIV亚型,Karim等也曾于国内非人灵长类中检测出此亚型[15]。而bg基因位点和tpi基因位点的差异性较大,分别鉴定出5和6个亚型。其中bg位点中的BIII亚型和tpi位点中的EB5,BIV和B7均为人兽共患亚型。
本研究首次采用多位点基因序列分析法对西南部分地区圈养非人灵长类贾第虫感染情况进行调查,同时对感染虫种亚型进行鉴定。结果表明,西南部分地区非人灵长类贾第虫bg基因和tpi基因多样性高,亚型种类丰富,基于各位点鉴定出的亚型与国内其他地区已鉴定出的非人灵长类亚型存在一定差异,表明贾第虫集聚体亚型分布存在地理差异,可能由于传染源不同所致[1]。本实验鉴定的亚型中BIII、BIV、EB5都曾在人类中被发现,提示西南部分地区非人灵长类感染的贾第虫存在人兽共患风险,但人兽共患集聚体B在非人灵长类源贾第虫中是否为优势集聚体还需进一步研究。由于本次试验中发现了非人灵长类中存在人兽共患贾第虫亚型,因此在对圈养非人灵长类的饲养、保护和研究中,饲养员、研究人员和游客因能近距离直接或间接接触到圈养非人灵长类动物,因此对相关的人员存在潜在的威胁。此外,圈养非人灵长类动物可能通过粪便排泄有感染性的包囊而污染周围环境,所以在圈养环境下需要特别注意饮用水、生活用水以及食品安全等[4-5]。除此之外有必要对圈养非人灵长类动物进行定期、长期的贾第虫等人兽共患寄生虫感染的监测工作及驱虫工作。
[1] RYAN U, CACCIS M. Zoonotic potential ofGiardia[J].InternationalJournalforParasitology, 2013, 43(12/13):943-956.
[2] ECKMANN L. Mucosal defenses against Giardia[J].ParasiteImmunology, 2003, 25(5):259-270.
[3] RAYANI M, UNYAH N Z, HATAM G. Molecular identification ofGiardiaduodenalisisolates from Fars Province, Iran[J].IranianJournalofParasitology, 2014, 9(1):70-78.
[4] PLUTZER J, ONGERTH J, KARANIS P.Giardiataxonomy, phylogeny and epidemiology: Facts and open questions[J].InternationalJournalofHygieneandEnviromentalHealth, 2010, 213(5):321-333.
[5] THOMPSON R C. The zoonotic significance and molecular epidemiology ofGiardiaand giardiasis[J].VeterinaryParasitology, 2004, 126(1/2):15-35.
[6] FENG Y, XIAO L. Zoonotic potential and molecular epidemiology ofGiardiaspecies and giardiasis[J].ClinicalMicrobiologyReviews, 2011, 24(1):110-140.
[7] SULAIMAN I M, FAYER R, BERN C, et al. Triose phosphate isomerase gene characterization and potential Zoonotic transmission of multispeciesGiardiaduodenalis[J].EmergingInfectiousDiseases, 2003, 9(11):1444-1452.
[8] APPELBEE A J, FREDERICK L M, HEITMAN T L, et al. Prevalence and genotyping ofGiardiaduodenalisfrom beef calves in Alberta, Canada[J].VeterinaryParasitology, 2003, 112(4):289-294.
[10] KARIM M R, ZHANG S, JIAN F, et al. Multilocus typing ofCryptosporidiumspp. andGiardiaduodenalisfrom non-human primates in China[J].InternationalJournalforParasitology, 2014, 44(13):1039-1047.
[11] LI J Q, DONG H J, WANG R J, et al. An investigation of parasitic infections and review of molecular characterization of the intestinal protozoa in nonhuman primates in China from 2009 to 2015[J].InternationalJournalforParasitology:ParasitesandWildlife, 2017, 6(1):8-15.
[12] YE J B, XIAO L H, LI J, et al. Occurrence of human-pathogenicEnterocytozoonbieneusi,GiardiaduodenalisandCryptosporidiumgenotypes in laboratory macaques in Guangxi, China[J].ParasitologyInternational, 2014, 63(1):132-137.
[13] DU S Z, ZHAO G H, SHAO J F, et al.Cryptosporidiumspp.,Giardiaintestinalis, andEnterocytozoonbieneusiin captive non-human primates in Qinling Mountains[J].KoreanJournalofParasitology, 2015, 53(4):395-402.
[14] YE J B, XIAO L H, MA J B, et al. Anthroponotic enteric parasites in monkeys in public park, China[J].EmergingInfectiousDiseases, 2012, 18(10):1640-1643.
[15] KARIM M R, WANG R J, YU F C, et al. Multi-locus analysis ofGiardiaduodenalisfrom nonhuman primates kept in zoos in China: Geographical segregation and host-adaptation of assemblage B isolates[J].Infection,GeneticsandEvolution, 2015, 30:82-88.
[16] LEE M F, CADOGAN P, EYTLE S, et al. Molecular epidemiology and multilocus sequence analysis of potentially zoonoticGiardiaspp. from humans and dogs in Jamaica[J].ParasitologyResearch, 2016, 116(1):1-6.
[17] SRICHARERN W, INPANKAEW T, KEAWMONGKOL S, et al. Molecular detection and prevalence ofGiardiaduodenalisandCryptosporidiumspp. among long-tailed macaques (Macacafascicularis) in Thailand[J].Infection,GeneticsandEvolution, 2016, 40:310-314.
[18] 魏小红,陈守义. 贾第鞭毛虫和隐孢子虫的流行病学研究进展[J].热带医学杂志,2014, 14(2):265-268.
WEI X H, CHEN S Y. Current advancement of epidemiology on Giardia and Cryptosporidium[J].JournalofTropicalMedicine, 2014, 14(2):265-268. (in Chinese)
