基于COI基因和D-loop区部分序列比较雷州半岛东、西岸湖栖鳍虾虎鱼群体的遗传多样性
2017-05-24龙水生郭昱嵩刘楚吾王中铎
廖 健, 龙水生, 贺 亮, 郭昱嵩, 刘楚吾, 刘 丽, 王中铎
基于基因和D-loop区部分序列比较雷州半岛东、西岸湖栖鳍虾虎鱼群体的遗传多样性
廖 健, 龙水生, 贺 亮, 郭昱嵩, 刘楚吾, 刘 丽, 王中铎
(广东海洋大学水产学院南海水产经济动物增养殖广东普通高校重点实验室, 广东湛江524025)
为比较分析雷州半岛东、西两岸湖栖鳍虾虎鱼()遗传差异, 以基因和D-loop区部分序列为分子标记, 获取雷州半岛湖栖鳍虾虎鱼群体序列44条(九龙山群体21条, 高桥群体23条)、D-loop序列53条(九龙山群体25条, 高桥群体28条)。基于基因部分序列分析结果显示: 619 bp的44条序列中, 统计的变异位点29个; T、C、A、G碱基组成分别为: 27.8%、31.0%、23.7%、17.5%, 且A + T(51.5%)高于C + G(48.5%); 单倍型多样性比较, 九龙山群体(0.695)>高桥群体(0.324); 遗传分化分析表明东、西群体分化显著(= 0. 0.591, 0.01 << 0.05), 基因流()为5.37, 遗传分化系数()为0.147, 固定指数()为0.042(0.001<<0.01), 核苷酸差异数()为2.547, 核苷酸歧义度()为0.004。而基于D-loop区部分序列分析结果表明: 563 bp的53条序列中, 变异位点133个; T、C、A、G碱基组成分别为: 32.3%、14.8%、35.5%、17.4%, A + T(67.8%)明显高于C + G(32.2%); 单倍型多样性与分析结果类似, 九龙山群体(0.797)>高桥群体(0.661); 遗传分化分析表明东、西群体分化极显著(= 0.970,< 0.001),为0.315(<0.001),值为0.55,、分别为: 0.157、3.506、0.006; AMOVA分析揭示群体内变异程度高于群体间; 聚类分析表明, 基于基因与D-loop区部分序列构建的邻位链接法聚类树(NJ树)均存在按照采样点聚类的现象, 但D-loop更为明显。综上, 雷州半岛东、西两岸湖栖鳍虾虎鱼群体出现较为明显的分化现象。
; D-loop; 湖栖鳍虾虎鱼(); 雷州半岛群体; 遗传分化
湖栖鳍虾虎鱼()隶属于硬骨鱼纲(Osteichthyes)、鲈形目(Perciformes)、虾虎鱼科(Gobiidae)、鳍虾虎鱼属(), 是暖水性的小型底栖鱼类, 无经济价值。其头、体呈透明状, 经体积分数为5%的甲醛溶液浸泡后呈现乳白色。成熟个体约为2 cm, 据FishBase(http: //www.fishbase.org. cn/search.php)记载最长记录为2.4 cm, 雌鱼个体通常较雄鱼大。已有文献及FishBase记载, 目前该物种仅发现于菲律宾吕宋岛的淡水湖泊中, 为该地区特有种[1]。《中国动物志》硬骨鱼纲、鲈形目、虾虎鱼亚目中表明产于中国的鳍虾虎鱼属仅1种, 即大鳞鳍虾虎鱼()[2]。廖健[3]和柏琴[4]调查发现在中国雷州半岛沿岸红树林海区及河口地带可以采集到鳍虾虎鱼属的湖栖鳍虾虎鱼[3-4]。鉴于日益恶化的全球水域环境及现有的红树林资源状况, 结合湖栖鳍虾虎鱼特殊的形态特征: 鱼体终生呈透明状, 内部器官结构清晰可见, 且广泛存在于雷州半岛红树林海区, 是筛选雷州半岛地方性水域环境监测物种的优质鱼类之一。
线粒体基因组(mtDNA)是脱离于核基因之外的遗传物质之一, 其结构类似于原核生物, 呈环状, 具有结构简单、分子片段短、进化速率较核DNA快、遵循母系遗传等特征[3, 5-6]。细胞色素氧化酶I基因()位于线粒体DNA中, 是线粒体基因组13种氨基酸编码序列之一, 受到较大的选择压力, 进化速率适中[7]。近十年来,被广泛应用于物种分类及种内种间遗传分化研究。D-loop区又称控制区, 是mtDNA上变异频率较高的一段区域, 不参与氨基酸的编码, 选择压力处在较低水平, 为线粒体基因组进化速率最快的区域[8]。D-loop区也是研究群体多样性及系统进化的优质分子标记, 受到了越来越多的群体遗传研究者的关注。
目前, 对于湖栖鳍虾虎鱼的研究, 国内外仅局限于资源调查及分类工作[1]。柏琴发现湖栖鳍虾虎鱼为雷州半岛东部红树林海区的优势物种[4]。本研究在此基础上, 分别以基因、D-loop区部分序列作为分子标记进一步探讨湖栖鳍虾虎鱼雷州半岛东部九龙山群体与西部高桥群体之间的遗传结构及分化情况。从而为湖栖鳍虾虎鱼的种质资源保护及合理开发利用提供科学依据。
1 材料与方法
1.1 样本采集
实验用鱼利用孔径149mm的手抄网分别采集于雷州半岛东岸的九龙山红树林国家湿地公园(N20°39¢51, ” E110°17'13")及西岸的高桥红树林保护区(N21°36'24, ” E109°47'8"), 见图1。其中用于扩增的44个样本(高桥群体23尾, 体长在1.15~ 2.05 cm;九龙山群体21尾, 体长在1.38~2.16 cm)采集于2014年4月~2015年4月, 用于D-loop扩增的53个样本(高桥群体28条, 体长在0.86~2.08 cm; 九龙山群体25条体长在0.95~2.18 cm)采集于2015年8~11月, 采集的样本利用体积分数75%的乙醇保存分装至2 mL EP管后带回实验室−40℃保存备用。
1.2 DNA提取、扩增及测序
每个样本剪取0.5 g肌肉进行基因组DNA的提取, 采用传统苯酚-氯仿法进行反复抽提[9]。沉淀的DNA干燥处理后, 溶于50 μL TE缓冲液中。基因的扩增引物采用硬骨鱼类通用引物, Fish F, 5¢-TC AACCAACCACAAAGACAATGGCAC-3¢; Fish R, 5¢- TAGACTTCTGGGTGGCCAAAGAATCA-3¢[10]。D-loop区序列扩增参照宋娜等报道的引物序列, Dloop F: 5¢-CCCATCTCTAGCTCCCAAAGC-3¢; Dloop R: 5¢- CTGTAGAGTGAACGCTTGGCATG -3¢[11]。引物由上海生工生物工程有限公司合成。PCR体系中10× Buffer(含Mg2+)2.5 μL、2.5 mmol/L dNTP 2 μL、5 μmol/L引物各1 μL、DNA模板1 μL、500 u/L Taq DNA聚合酶0.2 μL, 补充灭菌ddH2O至25 μL。基因的PCR程序设置为: 94℃ 1 min 20 s; 94℃ 30 s, 55℃ 30 s, 72℃ 50 s, 35个循环; 72℃ 5 min; 4℃保存。D-loop扩增程序: 94℃ 1 min 20 s, 94℃ 30 s, 54℃ 30 s, 72℃ 50 s, 循环35次, 72℃ 10 min, 4℃保存。PCR产物经10 mg/mL琼脂糖凝胶电泳检测后寄至上海生工生物有限公司进行序列测定。
1.3 数据分析
测序返回数据经MEGA6.0进行多序列比对, 人工校正后, 统计变异位点数、单一突变位点、简约信息位点、碱基组成、Kimura 2-parameter遗传距离[12], 利用Modeltest确定最适合的进化模式[13], 邻位连接法(Neighbor-Joining, NJ)构建系统进化树, Bootstrap 置信值重复抽样1 000次, 若转换/颠换比值小于2, 另需利用DAMBE进行替代饱和性检验, 构建进化树前进行加权[14]。单倍型数目、单倍型多样性、核苷酸多样度、平均核苷酸差异以及中性检验的Tajima’D及Fu - Li D值的统计利用DnaSP v5完成[15], 近邻统计、Lei模型基因流、遗传分化系数、固定指数、核苷酸差异数、核苷酸歧义度均由DnaSP v5中基因流和遗传分化估算得到[16], 分子方差分析(AMOVA)由DnaSP v5分组处理而后由Arlequin3.2.2.5统计得到。
2 结果分析
2.1 序列分析
基于基因的序列分析结果表明, 测序所得序列经Mega6.0序列比对、人工校正后所得序列长619 bp, 分别与NCBI及BOLD数据库进行比对, 相似度均在98%~100%。分析44条序列结果显示, 619 bp中变异位点数为29个, 包含单一突变位点9个, 简约信息位点20个。T、C、A、G碱基组成分别为: 27.8%、31.0%、23.7%、17.5%, 其中A + T(51.5%)高于C + G(48.5%), 且碱基在密码子第1、2和3位的分布并不均匀, 呈现一定的偏倚性, 其中碱基T明显偏向第1位(41%), 碱基G则在第3位出现频率最高(29.7%)(表1); 基于D-loop区序列分析结果表明, 563bp中变异位点133个, 单一突变位点117个, 简约信息位点16。T、C、A、G碱基组成为: 32.3%、14.8%、35.5%、17.4%, 同样A + T(67.8%)高于C+G (32.2%), D-loop区序列4种碱基在密码子的1, 2和3位分布较为均匀, 没有出现明显的偏倚性。所得序列上传至GenBank获取登录号: KU900420- KU900437, KU900440-KU900465, D-loop区登录号为: KU900367-KU900419。

表1 基于COI基因和D-loop部分序列的碱基组成(%)
2.2 遗传变异
湖栖鳍虾虎鱼两个群体的基因及D-loop区部分序列的遗传多样性参数统计结果显示(表2), 基于基因的44条序列总的单倍型比例、单倍型多样性、核苷酸多样度、平均核苷酸差异分别为: 20.45%、0.590、0.004、2.468。基于D-loop区的53条序列总的单倍型比例、单倍型多样性、核苷酸多样度、平均核苷酸差异分别为: 30.19%、0.863、0.005、2.946。

表2 基于COI基因及D-loop区部分序列的高桥与九龙山群体遗传多样性参数
遗传分化、基因流表明, 基于基因部分序列的高桥群体和九龙山群体的近邻统计()值为0.591(0.01<<0.05), 表明湖栖鳍虾虎鱼的雷州半岛东岸九龙山群体与西岸高桥群体分化显著, Lei模型基因流()为5.37, 种群间存在较强的基因交流, 遗传分化系数()为0.147, 固定指数()为0.042 (0.001<<0.01), 核苷酸差异数()为2.547, 核苷酸歧义度()为0.004。而基于D-loop区序列的高桥群体和九龙山群体为0.970(<0.001), 表明东、西两岸的湖栖鳍虾虎鱼群体分化极显著,值为0.55,为0.157,为0.315(<0.001),、分别为3.506、0.006, 见表3。
基于基因部分序列对雷州半岛东、西岸两个湖栖鳍虾虎鱼群体的分子方差分析(AMOVA)结果指示, 群体内变异(95.825%)程度显著高于群体间(4.175%)。而基于D-loop区部分序列的AMOVA分析显示, 群体内遗传变异(68.488%)同样高于群体间(31.512%), 且约为2倍关系, 见表4。
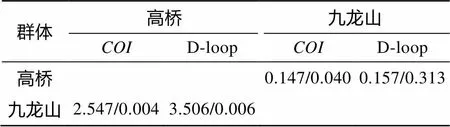
表3 基于COI基因及D-loop区部分序列的高桥与九龙山群体遗传分化统计
注 : 左下方为/, 右上方为/
2.3 遗传距离及系统进化
基于基因部分序列的两个群体间的Kimura 2-parameter平均遗传距离为0.0041, 估算转换/颠换值为1.15, 进行替代饱和性分析, 并构建转换颠换对TN93校正后的遗传距离散点图(图2), 由图2可见, 随着遗传距离的增加, 转换()、颠换()均在增大, 且呈线性增长关系, 具有较强的系统发育信号, 而后进行加权构建系统进化树分析。基于D-loop区部分序列的平均遗传距离为0.0144, 转换/颠换值为2.12大于2, 可直接用于建树分析。以大弹涂鱼()为外群构建NJ聚类树可见, 基于基因及D-loop区部分序列构建的NJ树均未完全聚为两个支系, 但部分样本按照采集地点先聚为一支。其中基于基因部分序列的两个群体聚类情况较D-loop区分散, 而依据D-loop区部分序列的两个地理种群大部分按采样点聚类(图3)。

表4 基于COI基因及D-loop区部分序列的湖栖鳍虾虎鱼群体AMOVA多样性分析
注 : 基于基因的=0.042(0.001<<0.01); 基于D-loop序列的=0.315(<0.001)
2.4 中性检验及群体扩张
基于基因的高桥群体Tajima’s D test、Fu - Li D test中性检验结果均不显著, 表明高桥群体符合中性进化假设, 而九龙山的检测结果均表现极显著, 即九龙山群体不符合中性进化。基于D-loop区序列的高桥群体Tajima’s D检验结果显著, 不符合中行进化假设, Fu - Li D检验显著, 九龙山群体Tajima’s D、Fu-Li D检验结果均表现不显著, 表明依据D-loop区序列的九龙山群体符合中行进化假设(表5)。
湖栖鳍虾虎鱼的雷州半岛东、西岸两个群体增长均符合期望值, 且核苷酸不配对分布曲线呈现不明显的单峰(图4)。反映湖栖鳍虾虎鱼近期可能经历了群体扩张过程, 说明了湖栖鳍虾虎鱼的高桥群体和九龙山群体存在分化。
3 讨论
线粒体基因组中的4种碱基A、T、C和G的分布并不均匀, 而具有明显的偏倚性。本研究获取的湖栖鳍虾虎鱼两个群体共44条基因619bp序列的碱基组成中A + T(51.5%)含量较C + G(48.5%)高, 其他硬骨鱼类线粒体基因序列同样具有类似碱基组成特点, 如银鲳、鳕鱼及鲷属鱼类等[17-20]。53条D-loop区序列563bp碱基中, A + T(67.8%)含量远远高于C + G(32.2%), 这与彭士明[7]研究的野生及养殖银鲳鱼群体结果相似。
▲. 外群物种大弹涂鱼(); ○. 高桥群体; ●. 九龙山群体
▲.; ○. Gaoqiao population; ●. Jiulongshan population

表5 基于COI基因(上行)和D-loop区(下行)序列的中性检验参数
注 : ns.不显著; *. 0.01 << 0.05; **. 0.001 << 0.01; ***.< 0.001
A、B. 基于基因的mismatch分布分析; C、D. 基于D-loop区的mismatch分布分析; Obs-GQ. 高桥观测值; Obs-JLS.九龙山群体观测值; Exp. Constant model期望值
A、B. based ongene; C、D. based on control region; Obs-GQ. observed value by the Gaoqiao population; Obs-JLS. observed value by the Jiulongshan population; Exp. expected constant model
线粒体基因组与核基因相比, 具有结构简单、分子小、进化快等特征而被广泛应用于群体分化及遗传多样性的研究[21-22]。依据D-loop区构建的进化树绝大部分个体按照采样点进行聚类, 只有少部分个体出现了东、西群体交错现象, 这可能是由于海洋环流或渔船压舱水等原因致使鱼卵、仔稚幼鱼运输带来的种群交流, 湖栖鳍虾虎鱼鱼卵为漂流性卵水体中悬浮于水层中能随海洋环流或压舱水而进行(又称半浮性卵), 这类鱼卵在产于母体后吸水膨胀, 出现较大的卵周隙, 比重略大于水, 在流动的水体中随着水流迁移, 在静水中则下沉至底部, 这是湖栖鳍虾虎鱼雷州半岛东、西部群体发生低频率交流的内在原因之一。而依据基因构建的进化树中, 东、西部群体交错现象的个体出现得相对多些, 除了湖栖鳍虾虎鱼鱼卵特性及海洋环流及渔船压舱水造成的影响之外, 一定程度上也由基因本身特性所决定。基因与D-loop相比, 其进化速率相对较慢, 这也是基因作为物种鉴定分子依据的原因之一[23]。尽管在聚类分析上并没有如D-loop般明显的分化现象, 但单倍型多样性分析结果显示, 依据基因的九龙山群体单倍型多样性(0.695)>高桥群体单倍型多样性(0.324), 与基因相似, 基于D-loop区的九龙山群体单倍型多样性(0.797)>高桥群体单倍型多样性(0.661)。同时, 统计各群体的变异位点, 结果表明, 44条619 bp的基因中存在变异位点29个, 而在53条563bp的D-loop区序列中检测到变异位点133个。
综上说明线粒体基因及D-loop均可用于湖栖鳍虾虎群体遗传多样性分析, 但基因在聚类分析上不及D-loop区序列敏感, 这与彭士明等[7]基于和D-loop区研究银鲳的野生群体和养殖群体遗传多样性的结果一致。综合了基因及D-loop区各参数数据的分析结果得出结论: 雷州半岛东岸九龙山湖栖鳍虾虎鱼群体与西岸的高桥群体出现了较为明显的分化, 但也存在海洋物理环境带来的种群交流现象。总之, 九龙山群体遗传多样性较高桥群体高。推测原因, 可能与其处在的海域存在一定联系。高桥位于半封闭的北部湾东岸雷州半岛西部, 海洋环境相对稳定, 而九龙山面临着广阔的南海海域, 受到台湾暖流影响。对环境适应能力较强的群体往往具有较高的遗传多样性或遗传变异, 而长期生活在稳定环境下的遗传多样性较低的群体则更容易受到环境变化的影响。与已经报道的鱼类比较[24-26], 尽管雷州半岛东、西两岸的湖栖鳍虾虎鱼出现了较为明显的分化现象, 但两岸群体的遗传多样性仍处在中等水平。这一状况提示对雷州半岛湖栖鳍虾虎鱼资源合理的管理及保护是当务之急的要务, 同时为后期雷州半岛红树林海区水域环境的监测开发地方性标志鱼类提供保障。
[1] Aquino L M G, Tango J M, Canoy R J C, et al. DNA barcoding of fishes of Laguna de Bay, Philippines[J]. Mitochondrial DNA, 2011, 22(4): 143-153.
[2] 伍汉霖, 钟俊生. 中国动物志[M]. 北京: 科学出版社, 2008: 1-14. Wu Hanlin, Zhon Junsheng. Fauna Sinica[M]. Beijing: Science Press, 2008: 1-14.
[3] 廖健, 张顺, 龙水生, 等. 5种虾虎鱼类线粒体基因序列变异及系统进化[J]. 广东海洋大学学报, 2016, 36(1): 7-12. Liao Jian, Zhang Shun, Long Shuisheng, et al. Sequence variation and molecular phylogeny of mitochondrialgene segments from five species of Gobiidae Family[J]. Journal of Guangdong Ocean University, 2016, 36(1): 7-12.
[4] 柏琴. 雷州半岛东部红树林海区鱼类分子分类研究[D]. 湛江: 广东海洋大学, 2014: 21. Bai Qin. Molecular classification of fishes in the eastern of Leizhou mangrove[D]. Zhanjiang: Guangdong Ocean University, 2014: 21.
[5] 杜民, 尹绍武, 刘艳红, 等. 中国裸胸鳝属 10 种鱼类分子系统发育关系的 16S rDNA 分析[J]. 海洋科学, 2013, 37(6): 16-23. Du Min, Yin Shaowu, Liu Yanhong, et al. Analysis of the molecular phylogenetic relationships of 10species from China Seas based on 16S rDNA fragment sequences[J]. Marine Sciences, 2013, 37(6): 16-23.
[6] 孙超, 刘志鸿, 杨爱国. 基于线粒体和16S rRNA基因研究3个地理群体黑龙江河蓝蛤的遗传多样性[J]. 海洋科学, 2015, 39(1): 39-45. Sun Chao, Liu Zhihong, Yang Aiguo, et al. The genetic diversity of three populations ofbased on mitochondrialand 16S rRNA gene[J]. Marine Sciences, 2015, 39(1): 39-45.
[7] 彭士明, 施兆鸿, 侯俊利. 基于线粒体D-loop区与基因序列比较分析养殖与野生银鲳群体遗传多样性[J]. 水产学报, 2010, 34(1): 19-25. Peng Shiming, Shi Zhaohong, Hou Junli. Comparative analysis on the genetic diversity of cultured and wild silver pomfret populations based on mtD-loop andgene[J]. Journal of Fisheres of China, 2010, 34(1): 19-25.
[8] 刘良国, 杨品红, 王文彬, 等. 洞庭青鲫与其他六个鲫鱼品系线粒体 DNA 控制区的比较分析[J]. 水生生物学报, 2010, 34(2): 378-387. Liu Liangguo, Yang Pinhong, Wang Wenbin, et al. Comparative analysis of the mitochondrial DNA control region in Dongtingking crucian carp and other six strains of carassius[J]. Acta Hydrobiologica Sinica, 2010, 34(2): 378-387.
[9] Sambrook J, Fritsch E P, Manlatis T. Molecular cloning: a laboratory manual[M]. New York: Cold Spring Harbor Laboratories Press, 1989: 31-46.
[10] 孟玮, 杨天燕, 海萨, 等. 基于线粒体基因序列的亚东鲑DNA条形码研究[J]. 水产学杂志, 2010, 23(1): 2010-2014. Meng Wei, Yang Tianyan, Hai Sa, et al. Study of DNA barcoding based on the mitochondrialGene sequences in[J]. Chinese Journal Fisheries, 2010, 23(1): 2010-2014.
[11] 宋娜, 宋林, 高天翔, 等. 斑尾复虾虎鱼群体遗传多样性比较分析[J]. 水产学报, 2011, 35(3): 321-326. Song Na, Song Lin, Gao Tianxiang, et al. Comparative analysis of genetic diversity ofbased on the mitochondrial DNA control region[J]. Journal of Fisheres of China, 2011, 35(3): 321-326.
[12] Tamura K. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Molecular Biology and Evolution, 2011, 28(10): 2731- 2739.
[13] Posada D, Crandall K A. Modeltest: testing the model of DNA substitution[J]. Bioinformatics, 1998, 14 (9): 817-818.
[14] 徐玲, 罗玉萍, 周冬根, 等. U83 Box C/D snoRNA构建果蝇科系统发生树[J]. 细胞生物学杂志, 2007, 29: 758-762. Xu Ling, Luo Yuping, Zhou Donggen, et al. U83 box C/D snoRNA as a molecular marker in the phylogeny of Drosophilidae[J]. Chinese Journal of Cell Biology, 2007, 29: 758-762.
[15] Librado P R, Dna S P. A software for comprehensive analysis of DNA polymorphism data[J]. Bioinformatics, 2009, 25: 1451-1452.
[16] 王中铎, 郭昱嵩, 颜云榕, 等. 南海大眼金枪鱼和黄鳍金枪鱼的群体遗传结构[J]. 水产学报, 2012 , 36(2): 191-201. Wang Zhongduo, Guo Yusong, Yan Yunrong, et al. Population genetics of tunas in South China Sea inferred from control regions[J]. Journal of Fisheres of China, 2012, 36(2): 191-201.
[17] 彭士明, 施兆鸿, 侯俊利, 等. 银鲳3个野生群体线粒体基因的序列差异分析[J]. 上海海洋大学学报, 2009, 18(4): 398-402. Peng Shiming, Shi Zhaohong, Hou Junli, et al. Genetic diversity of three wild silver pomfret () populations based ongene sequences[J]. Journal of Shanghai Ocean University, 2009, 18(4): 398-402.
[18] 张凤英, 马凌波, 施兆鸿, 等. 两种鲷属鱼类线粒体基因片段序列的比较[J]. 上海水产大学学报, 2006, 15(4): 403-408. Zhang Fengying, Ma Lingbo, Shi Zhaohong, et al. Studies on mitochondrial cytochrome oxidase subunit I fragments of two genera offish[J]. Journal of Shanghai Ocean University, 2006, 15(4): 403-408.
[19] 毕潇潇, 高天翔, 肖永双, 等. 4种鳕鱼线粒体16S rRNA、和基因片段序列的比较研究[J]. 南方水产, 2009, 5(3): 46-52. Bi Xiaoxiao, Gao Tianxiang, Xiao Yongshuang, et al. Sequence comparison of mtDNA 16SrRNA,andgene fragments in four species offish[J]. South China Fisheries Science, 2009, 5(3): 46-52.
[20] 窦新杰, 常玉梅, 唐然, 等. 几种雅罗鱼亚科鱼类基于mtDNA序列的亲缘关系[J]. 江苏农业学报, 2014, 30(4): 826-832. Dou Xinjie, Chang Yumei, Tang Ran, et al. Relationship of Leuciscinae fish based on mtDNA sequence[J]. Jiangsu J of Agr Sci, 2014, 30(4): 826-832.
[21] Abinash P. Geographic variation within a tandemly repeated mitochondrial DNA D-loop region of a North American freshwater fish,[J]. Gene, 2014, 538(1): 63-68.
[22] 严加坤, 杨爱国, 周丽青, 等. 基于线粒体16S rRNA基因研究5个栉江珧野生群体的遗传多样性[J]. 海洋科学, 2013, 37(2): 36-42. Yan Jiakun, Yang Aiguo, Zhou Liqing, et al. The genetic diversity of five populations ofin China based on mitochondrial 16S rRNA gene[J]. Marine Sciences, 2013, 37(2): 36-42.
[23] 于亚男, 宋超, 侯俊利, 等. 基于线粒体基因部分序列的长江口虾虎鱼科鱼类系统分类[J]. 淡水渔业, 2014, 44(5): 3-8. Yu Yanan, Song Chao, Hou Junli, et al. System classification for Gobiidae derived from the Yangtze River based on mitochondrialgene partial sequence[J]. Freshwater Fisheries, 2014, 44(5): 3-8.
[24] Liu J X, Gao T X, Yokogawa K, et al. Differential population structuring and demographic history of two closely related fish species, Japanese sea bass () and spotted sea bass () in Northwestern Pacific[J]. Molecular Phylogenetics and Evolution, 2006, 39(3): 799-811.
[25] Liu J X, Gao T X, Zhuang Z M, et al. Late Pleistocene divergence and subsequent population expansion of two closely related fish species, Japanese anchovy () and Australian anchovy ()[J]. Molecular Phylogenetics and Evolution, 2006, 40(3): 712-723.
[26] Xiao Y S, Zhang Y, Gao T X, et al. Genetic diversity in the mtDNA control region and population structure in the small yellow croaker[J]. Environmental Biology of Fishes, 2009, 85(4): 303- 314.
Genetic diversity ofpopulations from western and eastern Leizhou Peninsula based on partial sequences of mitochondrialgene and D-loop
LIAO Jian, LONG Shui-sheng, HE Liang, GUO Yu-song, LIU Chu-wu, LIU Li, WANG Zhong-duo
(Key Laboratory of Aquaculture in South China Sea for Aquatic Economic Animal of Guangdong higher Education Institutes, Fisheries College, Guangdong Ocean University, Zhanjiang 524025, China)
We compared twopopulations collected from the eastern and western Leizhou Peninsula based ongene and D-loop region.The partial sequences ofgene from 44 individuals (21 from eastern Jiulongshan population and 23 from western Gaoqiao population) and D-loop region from 53 individuals (25 from JLS and 28 from GQ) were cloned. Based ongene, the results were as follows: there were 29 variable sites in 619 bpsequences. The frequencies of T, C, A, and G were 27.8%, 31.0%, 23.7%, and 17.5%, respectively, with more A + T (51.5%) than C + G (48.5%). The haplotype diversity of Jiulongshan population (0.695) was higher than that of Gaoqiao population (0.324). Genetic differentiation analysis indicated that the eastern and western populations were differentiated significantly (= 0.591, 0.01 << 0.05). The correlative values of,,,, andwere 5.37, 0.147, 0.042, 2.547, and 0.004, respectively. However, much more variable sites (133 bp) were observed in D-loop regions with 563 bp length. In addition, there were higher frequencies of A + T (67.8%) than, and the haplotype diversity of Jiulongshan population (0.797) was higher than that of Gaoqiao population (0.661). Geneticdifferentiation of the eastern and western populations was highly significant (= 0.970,< 0.001). The values of,,,, andwere 0.157, 0.315, 3.506, and 0.006, respectively. Analysis of molecular variance revealed that the percentage of variation within populations was higher than that among populations. Cluster analysis showed the presence of clustering phenomenon, which gathered by the collected sites of sample based ongene and D-loop region, but D-loops were more apparent. In summary, a significant differentiation occurred between eastern and western Leizhou Peninsula.
gene; D-loop region;; population of Leizhou Peninsula; genetic differentiation
S917
A
1000-3096(2017)02-0103-08
10.11759//hykx20160319001
2016-03-19;
2016-04-25
国家自然科学基金项目(31201996); 广东省高等学校优秀青年教师培养计划项目(Yq2013089); 广东海洋大学优秀学位论文培育项目(201605)
廖健(1990-), 男, 湖南衡阳人, 硕士研究生, 从事南海经济鱼类发育生物学研究, 电话: 13078298801, E-mail: liaojiancc@ foxmail.com; 王中铎, 通信作者, E-mail: aduofa@gmail.com
Mar. 19, 2016
[National Natural Science Foundation of China, No.31201996; Project of Outstanding Young Teacher Training Program in Colleges and Universities of Guangdong province, No. Yq2013089; Excellent Thesis Breeding Program of Guangdong Ocean University, No. 201605]
(本文编辑: 谭雪静)
