不同生境米氏冰草种群的繁殖分配特征
2016-12-27金晓明高倩文林海俊
金晓明, 张 伟, 苟 琪, 高倩文, 林海俊
(1.呼伦贝尔学院生命与环境科学学院,内蒙古呼伦贝尔021008;2.呼伦贝尔市教育局,内蒙古呼伦贝尔021008)
不同生境米氏冰草种群的繁殖分配特征
金晓明1*, 张 伟1, 苟 琪1, 高倩文1, 林海俊2*
(1.呼伦贝尔学院生命与环境科学学院,内蒙古呼伦贝尔021008;2.呼伦贝尔市教育局,内蒙古呼伦贝尔021008)
由于米氏冰草品质好,营养丰富,适口性好,各种家畜均喜食;又因其返青早,能较早地为放牧家畜提供青饲料,因此,受到人们关注。然而近年来,草原沙漠化严重,牧草需要量供不应求,为了解米氏冰草种群在沙地的适应机制,本文采取种群繁殖生态学的方法,对米氏冰草种群在沙地植被恢复不同阶段,各构件的数量和生物量分配特性进行研究。结果表明:(1)在地面环境相对恶劣情况下,米氏冰草优先占据地下生态位空间的繁殖策略,以避免植物地上部分受到伤害,为植物的营养繁殖做好储备;(2)营养繁殖是米氏冰草的主要繁殖方式,且米氏冰草对地上分株的生物量分配比地下根茎高5.7~6.5倍;(3)随着沙化草地的植被次生演替进程,由流动沙地过渡到固定沙地阶段,米氏冰草种群由数量较少的大个体分株,变成数量较多的小个体分株,地上分株的分配比例也有所增加,而其有性繁殖的分配比率呈降低趋势。反应了米氏冰草在r-k-连续统中,实现由r-端向k-端转变的过程,并进一步揭示了无性系植物在沙地不同生境中繁殖扩散及调节适应机理,为种群繁殖生态学的研究提供理论依据。
种群;构件;营养繁殖;有性繁殖
繁殖是生物繁衍后代延续种族最基本的行为和过程,即产生新个体的过程。大多数根茎型无性系植物能够同时进行营养繁殖与有性繁殖,且这两种繁殖方式之间的权衡在不同物种间以及同一物种内部变化很大。无性系植物的营养繁殖与有性繁殖之间可能存在着权衡关系,即对营养繁殖的资源投入将会减少对有性繁殖的资源投入,而这种权衡关系可能是由环境条件、竞争力度、植物寿命和遗传等因素决定的(张玉芬等,2006)。种子和营养繁殖体分别被认为是无性系植物的有性繁殖后代和无性繁殖后代。由于种子成功地进行了遗传重组并能长距离传播,而且可通过休眠机制使植物安全地度过不良时期,因此种子后代可能有助于植物拓殖新的栖息地 (Huber等,2000;Nishitami等,1999)。无性系植物的营养繁殖习性使得个体在空间和资源利用、逃避环境风险等方面有着明显的优势(Cook等,1985)。营养繁殖后代由于母体的供养更容易安全度过幼龄期,实现成功定居,而且不必付出与有性繁殖相伴的代价(Lovett Doust等,1981)。
繁殖分配(RA)是指植物在繁殖过程中分配到繁殖部分的物质的数量,它有两种表达方式,即资源的多少和繁殖构件数量的多少 (Douglas等,1981)。植物在其生活史中,通过最佳的资源分配格局适应环境,最终目标是物种的持续生存和繁衍(钟章成等,1995)。植物繁殖分配与植物的生长、植物的生活史、植物所处的演替状态以及植物的生存环境都有极大的关系 (操国兴,2003;钟章成等,1995)。目前有关有性繁殖与营养繁殖间资源分配研究所得出的结论不一致 (Mendez,1999;Saikkonen等,1998;Westley,1993)。大致认为无性系植物的营养生长与有性繁殖方式间资源分配比例的变化可能受种群密度及基株个体大小等因素的影响。
生物量、能量、氮、磷、钾、微量元素及热量等常作为度量指标测定繁殖分配及繁殖投资,但由于采用的指标不同,所得到的繁殖分配差异较大。近20年来,对繁殖、繁殖分配进行了大量研究,不过至今仍未找到一种客观统一的度量标准,因为生物量、能量、矿质元素在植物体内的分配格局并非一致,而经常是等级式的,这是问题的关键所在(Romero等,1996)。
从植物的生活史上,无性系禾本科植物返青后先进入营养生长阶段,抽穗期至成熟期由于有性繁殖体的形成及逐渐生长,使其同时进行有性繁殖和营养繁殖,并减少对营养繁殖的养分投入,成熟期以后植物不再向有性繁殖体提供养分,使营养繁殖的投入增加。成熟期对无性系植物的资源分配进行研究,可以很好地了解植物有性繁殖与营养繁殖之间的权衡问题。
呼伦贝尔草地是世界上保存最完好的天然草地之一。但在人为活动的影响下,使草地逆向演替,向沙漠化方向发展,变成固定沙地、半流动沙地和流动沙地状态(卢欣石等,2000)。本文以呼伦贝尔沙化草地植被次生演替过程中的两个不同阶段,即流动沙地阶段和固定沙地阶段的单优势种米氏冰草为研究对象,采用种群繁殖生态学的野外调查方法,探讨无性系植物对各构件的资源及数量的分配特征,以揭示米氏冰草种群在异质生境下的繁殖适应机制。
1 材料与方法
1.1 研究区自然概况 研究区位于中国呼伦贝尔草原境内。地理位置为东经11°44′75″~118°54′76″,北纬49°08′87″~49°11′48″,海拔588.4~618.0m。具温带大陆干旱半干旱过渡气候特征。年均气温-1.5℃,降水量230~300mm(70%集中于夏秋季),而蒸发量为降水量的4~6倍;年均风速4.5m/s,最大风速为20m/s,全年8级以上大风日数平均30 d以上。地带性植被为以贝加尔针茅和羊草为优势种的群落。沙漠化草地的植被主要以草本植物种为主,其中有米氏冰草、东北牡蒿、扁蓄豆、狗尾草、百里香、猪毛菜、沙蓬、沙米等。另外,还零星分布着少量的灌木,如差不嘎蒿和小叶锦鸡儿等。
1.2 样地选择及研究方法 本研究是在以米氏冰草为优势种的流动沙地(P1)和固定沙地(P2)内进行,分别代表草地沙漠化演替过程中的两个不同阶段。
2015年8月1日,即米氏冰草的成熟期,对流动沙地和固定沙地的米氏冰草种群进行调查。采用单位面积挖出的取样方法,面积为25 cm× 25 cm,深度为30 cm,将四周的土挖开使样方成土柱状,连同地上部分一并装入样袋中,尽量保持土柱的完整性。每样地10个重复,编号后分置。注意按不同密度取样。回室内用水轻轻冲洗,保持所有地上和地下部分的自然联系,以便于鉴别。
按每个样方对生殖分株、营养分株和苗等构件进行计数,并测量根茎长度,然后按相同构件放在一起,80℃烘干至衡重后测其干重。这里把拔节并抽穗的分株视为生殖分株,把拔节但未抽穗的分株视为营养分株,把由芽发育而形成且露出地面并变绿,但未拔节的营养繁殖体均视为苗。
1.3 数据处理 数据采用Excel软件和SPSS l6.0软件处理。将调查数量指标均换算成1m2的数据指标,对米氏冰草无性系的各构件,包括根茎、生殖分株、营养分株和苗等指标,采用ANOVA的Duncan法进行方差分析,取各样方的平均值(M)及标准差(SD),对流动沙地和固定沙地的米氏冰草种群各构件性状进行差异显著性比较。各构件分配的计算方法如下:
总分株数量(或生物量)=生殖分株数量(或生物量)+营养分株数量(或生物量);
总生物量=总分株生物量+根茎生物量;
总分株生物量比例/%=总分株生物量/总生物量×100;
根茎生物量比例/%=根茎生物量/总生物量× 100;
生殖分株数量(或生物量)比例/%=生殖分株数量(或生物量)/总分株数量(或生物量)×100;
营养分株数量(或生物量)比例/%=营养分株数量(或生物量)/总分株数量(或生物量)×100。
2 结果与分析
2.1 各构件的数量特征 不同生境米氏冰草种群各构件的数量特征见表1。平均值代表种群水平的数量指标,标准差反映样本内的绝对变异度,最大值和最小值是样本的数值范围。固定沙地总分株数量、营养分株数量分别比流动沙地高1.9倍、2.6倍,且二者均达到显著水平;流动沙地生殖分株数量、根茎长度分别比固定沙地高1.3倍、1.1倍。表明米氏冰草在地面环境相对恶劣情况下,采取优先占据地下生态位空间的繁殖策略,以避免植物地上部分受到伤害,为植物的营养繁殖做好储备。
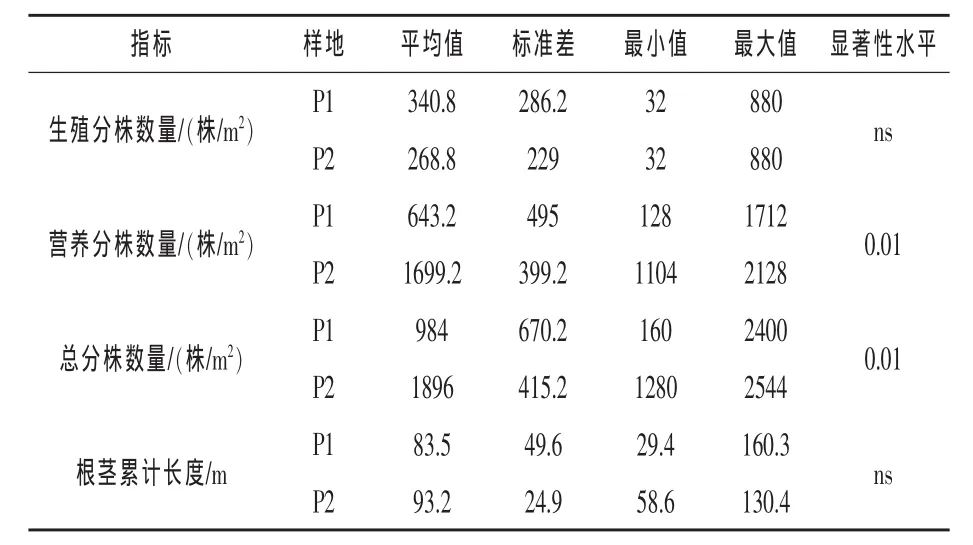
表1 不同生境米氏冰草各构件数量性状比较(样方大小:1m×1m)
2.2 各构件的生物量特征 不同生境米氏冰草种群各构件生物量特征见表2。流动沙地总分株生物量和生殖分株生物量分别比固定沙地高1.1倍、2.8倍;固定沙地营养分株生物量比流动沙地高2.2倍,且米氏冰草的生殖分株生物量和营养分株生物量分别在两个样地之间均达到差异显著性水平;两样地的根茎生物量相同;流动沙地生殖单株生产力和营养单株生产力分别比固定沙地高1.6倍、1.1倍。

表2 不同生境米氏冰草各构件生物量性状比较(样方大小:1m×1m)
2.3 各构件的数量及生物量分配 米氏冰草种群的分株数量及生物量分配比例见图1a和1b。流动沙地的生殖分株数量比例显著大于固定沙地,是固定沙地的3倍;固定沙地的营养分株比例显著大于流动沙地,是流动沙地的1.4倍。两个样地的地上及地下构件的生物量分配比例上,地上分株生物量均显著大于地下根茎生物量,其中流动沙地和固定沙地的分株生物量分别是根茎生物量的5.7倍和6.5倍。流动沙地的生殖分株生物量分配比率显著高于固定沙地,是固定沙地的2.6倍;而固定沙地的营养分株生物量分配比率显著高于流动沙地,是流动沙地的2.1倍。
3 讨论与结论
许多无性系植物由无性分株和连接分株的间隔子、根茎和匍匐茎等构成,具有独特的无性构件(Dong等,1994;Slade等,1987)。无性系植物可能通过间隔子长度和生物量分配等选择性地放置分株和储藏养分,缓解资源的时空异质性对其生长和繁殖的不利影响 (Bart等,2001;De Kroon等,1995;Dong等。1994)。流动沙地与固定沙地相比,风沙较大,地面环境较恶劣,不利于植物地上部分的生长。本研究中,流动沙地的米氏冰草与固定沙地的相比,减少了单位根茎长度上的分株数量。即米氏冰草在地面环境相对恶劣情况下,采取优先占据地下生态位空间的繁殖策略,以避免植物地上部分受到伤害,为植物的营养繁殖做好储备。这与欧活血丹在低养条件下,匍匐茎节间增长类似(Slade等,1987)。

图1 不同生境米氏冰草各构件数量及生物量的分配
生物量分配格局决定着植物在异质生境中获取资源的能力(Porter等,1990),从而影响其在群落中的竞争力(Tilman,1988;Grime等,1979)。本研究中,米氏冰草种群在生物量资源分配上,地上构件是地下构件5.7~6.5倍,且固定沙地的地上构件分配比例略高于流动沙地。这与其他的研究结果类似。在养分或水资源贫瘠的情况下,植物把更多的生物量分配到根部,从而有利于对养分或水资源的获取(King,2003;Mǔller等,2000);而在光照资源比较低的情况下,植物把生物量更多地分配到地上茎、叶部分,有利于对光资源的获取。在固定沙地,由于地面环境相对较好,风沙活动明显减少,植物地上和地下构件的密度均较高,对资源的竞争也激烈,尤其对沙地植被的限制性生态因子—水分的竞争,使土壤水分含量相对降低。因此米氏冰草采取优先发展地上分株的策略,充分利用植物的地上部分,竞争光和水分,为植物体积累更多的养分。
许多无性系植物同时具有有性繁殖和无性繁殖的特性,并竞争相同的资源(Abrahamson,1975)。在资源受限的情况下,对有性繁殖投入的增加是以减少无性繁殖的投入为代价的(Tilman,1988)。本研究中,流动沙地和固定沙地米氏冰草的营养分株数量分别是生殖分株数量的2倍和9倍,即营养繁殖是米氏冰草的主要繁殖方式。这与大多数根茎型禾草一致 (张晓娜等,2010)。 但米氏冰草在不同生境之间对有性繁殖和营养繁殖的分配比率有所差异。流动沙地米氏冰草的有性繁殖比例高于固定沙地,其生殖分株生物量是营养分株生物量的2倍。即在环境条件较恶劣的流动沙地,有利于米氏冰草的有性繁殖。米氏冰草通过有性繁殖形成更多种子,并散布到距离母体较远的地方,为种群寻找适应其生存的新栖息地提高机会。这与R-对策者的理论是相吻合的(MacArthur等,1967)。许多干旱地区的植物具有相似特性,如在环境恶劣的滩地,四合木种群的生殖值、地上年净生物量的生殖分配比生境条件好的丘地要高(徐庆等,2001)。生活在干旱地区的一年生植物—抱茎独行菜的生殖配置为50.36%,高于一次结实的草本植物的生殖投入(魏岩等,2003)。生活在荒漠中的旱麦草属4种短命植物的生殖分配高达55%~60%,远高于大多数一年生和多年生植物(吕玲等,2005)。一般生殖分株所消耗的能量大于营养分株。
在固定沙地,米氏冰草减少生殖分株分配比率,而增加营养分株分配比率,是其采取的一种低能耗的繁殖策略,即把用于生殖分配的能量转移到输出较多营养分株上。有研究认为,在生长受限条件下,无性系植物趋向于把更多的生物量分配给营养繁殖,减少有性繁殖(Eriksson,1997;Cook,1985;Abrahamson,1980)。米氏冰草在固定沙地,其营养分株生物量是生殖分株生物量的3倍,其营养分株数量是生殖分株数量的8.5倍。表明,在固定沙地,米氏冰草的生存对种群密度具有依赖性,是稳定环境的维护者。这与K-对策者的理论是相吻合的(MacArthur等,1967)。然而,植物对有性繁殖和营养繁殖的分配变化是复杂的,如遗传特性 (Hartnett,1990)、植物个体大小(Schmid等,1995;Hartnett,1990)、植物年龄 (Lopez等,2001)及种群年龄(Escarré等,1994)等都会影响种群的繁殖分配。本研究中,在流动沙地,米氏冰草的分株个体均大于固定沙地,尤其表现在生殖分株上,流动沙地的米氏冰草个体比固定沙地的高1.6倍。流动沙地的米氏冰草输出少量的大个体分株,可能是对较强风沙活动的一种适应机制。其中一部分分株被沙埋后形成了根茎,而一部分未被沙埋的较大个体的分株可吸收较充足的养分,以保证正常的生长和繁殖。但米氏冰草的生殖分配与个体大小是否有关,还有待进一步研究。
随着沙化草地的次生演替进程,由流动沙地过渡到固定沙地阶段,米氏冰草地上与地下构件的分配,以及有性与无性繁殖的分配变化,既反映了米氏冰草在r-k-连续统中,实现由r-端向k-端转变的过程,又反映了米氏冰草对资源的不同利用方式。
[1]操国兴.植物种群的生殖分配[J].四川林业科技,2003,24(2):25~29.
[2]卢欣石,何琪.内蒙古草原带防沙治沙现状、分区和对策[J].中国农业资源与区划,2000,21(4):58~62.
[3]吕玲,谭敦炎.旱麦草属4种短命植物的结实特性及生殖包装[J].新疆农业大学学报,2005,28(3):21~25.
[4]魏岩,周桂玲,谭敦炎.短命植物抱茎独行菜的生殖对策分析[J].新疆农业大学学报,2003,26(3):37~39.
[5]徐庆,刘世荣,臧润国,等.中国特有植物四合木种群的生殖生态特征—种群生殖值及生殖分配研究[J].林业科学,2001,37(2):36~41.
[6]张晓娜,哈达朝鲁,潘庆民.刈割干扰下内蒙古草原两种丛生禾草繁殖策略的适应性调节[J].植物生态学报,2010,34(3):253~262.
[7]张玉芬,张大勇.克隆植物的无性与有性繁殖对策[J].植物生态学报,2006,30(1):174~183.
[8]钟章成.植物种群的繁殖对策[J].生态学杂志,1995,14(1):37~42.
[9]Abraham son W G.Demography and vegetative reproduction.In:Solbrig OT(ed)Demography and evolution in plant populations(Botanical monograph 15)[J].Blackwell,Oxford,1980,18:89~106.
[10]Abrahamson W G.Reproductive strategies in dewberries[J].Ecology,1975,56:721~726.
[11]Bart F.Long-term disadvantages of selective root placement:root proliferation and shoot biomass of two perennial species in a 2-yesr experiment[J]. Journal of Ecology,2001,89,711~722.
[12]Cook R E.Grow th and development in clonal plant population.In:Jackson JBC,Buss LW,Cook RE eds.Population Biology and Evolution of Clonal Organisms[J].Yale University Press,New Haven,1985,45:259~296.
[13]De Kroon H,Hutchings M J.M orphological plasticity in clonal plants:the foraging concept reconsidered[J].Journal of Ecology,1995,83:143~152.
[14]Dong M,de Kroon H.Plasticity in morphology and biomass allocation in Cynodon dactylon,a grass species forming stolons and rhizomes[J].O ikos,1994,70:99~106.
[15]Dong M.Foraging Through Morphological Responses in Clonal Herbs[J]. PhD dissertation,Utrecht University Press,Utrecht,1994,49~83.
[16]Douglas D A.The balance between vegetative and sexual reproductions of Mimulus primuloides at different altitude in California[J].J.Ecol.,1981,69 (1):295~31.
[17]Eriksson O.Clonal life historiesand the evolution of seed recruitment.In:Kroon H de,Groenendael Jvan.The ecology and evolution of clonal plants[J]. Leiden:Backhuys,1997,211~226.
[18]Escarré,et al.An experimental study of the role of seedling density and neighbor relatedness in the persistence of Rumex acetosella in an old-field succession[J].Canadian Journal of Bot,1994,72:1273~1281.
[19]Grime JP.Plant Strategies and Vegetation Processes[J].Wiley:Chichester,1979:39~48.
[20]Hartnett D C.Size-dependent allocation to sexual and vegetative reproduction in four clonal composites[J].Oecologia,1990,84:254~259.
[21]Huber H,During H J.No long_term costs of meristem allocation to flowering in stoloniferous Trifolium species[J].Evolutionary Ecology,2000,14:731 ~748.
[22]King D A.Allocation of above-ground grow th is related to light in temperate deciduous saplings[J].Functional Ecology,2003,17:482~488.
[23]Lopez F,et al.Age changes in the vegetative vs.reproductive allocation by module demographic strategies in a perennial plant [J].Plant Ecology,2001,157:13~21.
[24]Lovett Doust L L.Population dynamics and local specialization in a clonal plant Tanunculus repens[J].The dynamics of ramets in contrasting habitats. Journal of Ecology,1981,69(3):743~755.
[25]MacArthur R H,Wilson E O.The theory of island biogeography[M]. Princeton:Princeton University Press,1967:66~71.
[26]Mendez M.Effects of sexual reproduction on grow th and vegetative propagation in the perennial Geophyte Arum italicum (Araceae)[J].Plant Biology,1999,1(7):115~120.
[27]Mǔller I,Schmid B,Weiner J.The efects of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants.Perspectives in Plant Ecology[J].Evolution and Systematics,2000,3(2):115~127.
[28]Nishitami S,Takada T,Kachi N.Optimal resource allocation to seeds and vegetative propagules under density-dependent regulation in Syneilesis palmate (Compositae)[J].Plant Ecology,1999,141,179~189.
[29]Porter H,Retakes C,Lambers H.Carbon and nitrogen economy of twenty-four wild species differing in relative grow th rate.Plant Physiology[J]. 1990,94(11):621~627.
[30]Romero JM,et al.Allocation of biomass and mineral elements in Melilotus segetalis:effects of NaCl Salinity and plant age[J].New phytol,1996,132 (4):565~573.
[31]Saikkonen K,Koivunen S,Vuorisalo T,et al.Interactive effects of pollination and heavy metals on resource allocation in Potentilla anserina L[J].Ecology,1998,79,1620~1629.
[32]Schmid B,Bazzaz F A.Size dependency of sexual reproduction and of clonal growth in two perennial plants[J].Canadian Journal of Botany,1995,73 (4):1831~1837.
[33]Slade A J,Hutchings M J.An analysis of the costs and benefits of physiological integration between ramets in the clonal perennial herb Glechoma hederacea[J].Oecologia,1987,73(8):425~431.
[34]Slade A J,Hutchings M J.The efects of nutrient availability on foraging in the clonal herh Glechoma hederacea[J].Journal of Ecology,1987,75(11):98~112.
[35]Tilman D.Plant strategies and the dynamics and structure of plant communities[M].Princeton:Princeton University Press,1988:138~142.
[36]Westley L C.The effect of inflorescence bud removal on tuber production in Helianthus tuberosus L[J].(Asteraceae).Ecology,1993,74(6):2136~2144.■
Because of the good quality,rich nutrition,good palatability and green early for grazing livestock,Agropyron michnoi are fed for livestock.So there is a increasingly popular with people.In recent years,owing to the threat of desertification,and demand of grass is in short supply.In order to understand the adaptation mechanisms of A.michnoi populations in the sandy land,use the method of population reproduction ecology,we studied the characteristics of number and biomass allocation ofmodulars at different growth areas in sandy land.The results showed that:(1)A.michnoi preferentially occupy the space of underground niche when it was poor in ground conditions,to avoiding the damaged of stems and leaves,and prepareing for vegetative propagation.(2)Vegetative reproduction was the main way of breeding and the biomass allocation to the aboveground tillers was 5.7~6.5 times higher than the underground rizomes.(3)With the process of secondary succession of vegetation community in desertification grassland,A.michnoi changed from the small number of larger individual ramets into the large number of smaller individual ramets,and the distribution ratio to the aboveground ramets was also increased,and the distribution of its sexual reproduction ratio was decreasing.At the same time,it was revealed the propagation and adjustment mechanism of clonal plants in different habitats in the sandy land,which provided a theoretical basis for the population reproductive ecological research.
population;modular;vegetative propagation;sexual reproduction
10.15906/j.cnki.cn11-2975/s.20162305
S816
A
1004-3314(2016)23-0020-05
国家自然科学基金项目(31560657);内蒙古自然科学基金项目(2014MS0304);内蒙古自治区应用技术研究与开发资金计划项目(2013);呼伦贝尔重点科技专项(2014)
*
