海南4个野生地理群体长肋日月贝AFLP分析
2016-12-21朱彩艳江世贵杨其彬张殿昌苏天凤
朱彩艳,江世贵,杨其彬,2,张殿昌,苏天凤,王 雨,2
(1中国水产科学研究院南海水产研究所,广州 510300;2中国水产科学研究院南海水产研究所热带水产研究开发中心,海南 三亚 572018)
海南4个野生地理群体长肋日月贝AFLP分析
朱彩艳1,江世贵1,杨其彬1,2,张殿昌1,苏天凤1,王 雨1,2
(1中国水产科学研究院南海水产研究所,广州 510300;2中国水产科学研究院南海水产研究所热带水产研究开发中心,海南 三亚 572018)
为了解长肋日月贝(Amusiumpleuronectes)种群遗传结构,本研究应用6对引物,采用AFLP分子标记技术对中国海南4个野生群体(共134个个体)的长肋日月贝进行遗传结构分析.结果显示,6对引物组合共扩增出203条扩增带,平均每对引物组合产生33.8条扩增带;扩增带中的多态性位点共97条,多态位点所占比例为47.78%.统计分析了4个野生群体的多态性位点、群体Nei氏遗传多样性指数和Shannon’s 信息指数等,并进行了UPGMA聚类分析.结果表明,4个群体的群体Nei氏遗传多样性指数在0.0957~0.1 135之间,Shannon’s 信息指数均在0.1 493~0.1 769之间,4个群体的遗传多样性均偏低.陵水群体(LS)和三亚群体(SY)的多态性位点的比率均为40.89%,潭门群体(TM)的多态性位点比率最低,为34.4 8%;4个不同群体间遗传距离都不大,SY与白马井群体(BMJ)间遗传距离最大,为0.0162,TM与BMJ间的遗传距离最小,为0.0049;没有发现群体特异性条带.分析了各群体内部的个体间遗传距离,结果表明,BMJ、TM、LS和SY群体内部的遗传距离分别为0.1 217、0.1 065、0.1 212和0.1 179.AMOVA分析结果显示,种群间的分化系数(Fst)为0.06834,基因流水平(Nm)为3.4 08.表明野生长肋日月贝的遗传变异主要来自群体内部,群体间的遗传分化程度不大.另外,在长肋日月贝的种质资源保护和良种繁育方面提出了建议.
长肋日月贝;AFLP;遗传多样性
0 引言
长肋日月贝(Amusiumpleuronectes)是扇贝科(Pectinidae)的一种经济贝类,是中国南方海区特有的优质食用贝类[1].其主要优点在于个体较大,生长迅速,壳薄、闭壳肌发达.相对于壳重、生殖腺重等其他重量性状,长肋日月贝的闭壳肌重与体重的相关关系最大,其对体重有极显著的正相关关系[2].其出肉率高,甚至都超过了海湾扇贝(Argopectensirradias)优质品种“中科红”[3].近年来长肋日月贝的资源量越来越少[4],已经引起人们的重视.国外对于长肋日月贝的研究主要集中在繁殖发育生物学等方面[5-6].有学者研究了泰国长肋日月贝的线粒体DNA的多样性[7],为长肋日月贝的育种提出了建议.近年来,我国已经在长肋日月贝的人工繁殖技术取得了成功[4],但是很少有长肋日月贝种群遗传结构方面的研究,制约了长肋日月贝的种质资源保护和良种繁育.
AFLP是一种高效分子标记技术,已广泛用于水产生物的遗传育种研究[8-10],可为水产生物的种质资源保护和优良性状品种的选育提供有力的技术指导.佟广香等利用AFLP技术分析了我国野生哲罗鱼(Huchotaimen)种质资源的遗传多样性,认为增殖放流可以提高哲罗鱼的资源量,增加其遗传多样性[11].潘洁等也利用AFLP技术分析了3个栉孔扇贝(Chlamysfarreri)群体的遗传多样性,指出可以通过引种提高我国栉孔扇贝的遗传多样性[12].本研究采集了中国海南岛的东部、西部和南部总共4个野生群体的长肋日月贝,利用AFLP分子标记技术对其种群遗传结构进行了分析.这对于摸清长肋日月贝野生种群的遗传结构有重要的意义,对于长肋日月贝的种质资源保护和今后大规模开发提供有用的理论参考.
1 材料和方法
1.1 材料
实验用4个野生长肋日月贝群体均来自中国海南岛,分别为白马井群体(BMJ)、潭门群体(TM)、陵水群体(LS)和三亚群体(SY). 4个群体采样时间均为2008年3月,样品规格相近,壳长平均7.8±1.2 cm,壳高平均8.0±1.3 cm.每个群体随机选取31~37个个体(表1),采闭壳肌组织于-80℃冷冻保存.

表1 用于实验分析的4个群体
1.2 基因组DNA提取
使用Tiangen公司生产的试剂盒,参照说明书进行DNA的提取.将获得的DNA标准化为50 ng/μl,-20 ℃下保存备用.
1.3 AFLP分析方法
1.3.1 基因组DNA双酶切
试验中用于AFLP分析的内切酶是EcoR I和MseI,均为NEB公司生产.反应体系为300ng DNA中加入10 U EcoR I酶和2 U Mse I酶,酶切反应在37 ℃下进行4.5 h.
1.3.2 接头的制备及连接
接头序列和扩增引物序列见表2. EcoR I的两条接头和Mse I的两条接头在使用前分别两两混合,95 ℃变性5 min,室温下自然冷却,以形成双链结构,供连接使用.连接反应在1 U T4 DNA连接酶(TAKARA公司生产)的催化下,于16℃反应18 h.

表2 AFLP分析用的接头与引物序列
1.3.3 PCR扩增
预扩增反应体系为,10× rTaq buffer 2.5 μl,连接产物4μl,EcoRI和MseI预扩增引物各0.2 μl(250 ng/μl), dNTPs(2mM each) 3μl,rTaq 酶 (5U/μl )0.3 μl,补足ddH2O至25μl;反应中反应条件为, 94℃ 预变性3 min, 94℃ 变性30 s, 56℃ 退火30 s,72℃ 延伸30 s,循环24次,72℃终止延伸10 min, 4℃ 保存.将预扩增产物稀释5倍作为选择性扩增的模板.选择性扩增反应体系为, 10× rTaq buffer 2.0μl,稀释后的预扩增产物1μl,EcoRI和MseI选择性扩增引物各2.0μl(100ng/μl), dNTPs(2mM each) 1.5 μl, rTaq 酶 (5U/μl )0.1 μl,补足ddH2O至20μl;反应条件为94℃预变性3 min, 94℃变性30 s, 65℃ 退火30 s, 72℃延伸1 min,循环9次,每个循环退火温度降低0.9℃;94℃ 变性30 s, 56℃ 退火30s, 72℃ 延伸1 min,循环24次, 72℃终止延伸10 min, 4℃ 保存. 2%琼脂糖电泳,120 V,15 min检测PCR产物.
1.3.4 聚丙烯酰胺凝胶电泳
取5μl选扩产物与等体积的2×上样缓冲液充分混匀,95℃变性5 min后,迅速置于冰上.点样2μl,在6%变性聚丙烯酰胺凝胶上电泳,仪器为BioRad3000型电泳仪,电泳前进行30 min预电泳,之后120W恒功率电泳80 min.银染法染色,室温下自然干燥,数码相机拍照并保存图像.
1.4 数据分析
根据所获得的AFLP聚丙烯酰胺凝胶电泳胶图,统计多样性位点比率,并采用POPGENE3.2 软件计算该种群内部平均观测等位基因数、平均有效等位基因数、平均基因多样性指数以及Shannon’s多样性信息指数,分析种群内遗传距离.然后根据遗传距离表,采用MEGA软件构建遗传距离聚类图,方法为不加权成对算术平均法[9].对4个群体的数据用Arlequin软件进行分子方差分析(AMOVA分析).
2 结果
2.1 AFLP扩增多态性

图1 E6-M6引物组合在三亚群体中扩增(最右一列为marker)
6对引物组合的扩增片段大小主要在50~400bp左右(图1).使用的6对引物组合共产生了203条扩增带,平均每对引物组合产生33.8条扩增带,扩增带中的多态性位点97条,多态位点占扩增带比例为47.78%.多态位点百分率最高的一对引物组合是E8-M8(64.9%).见表3.
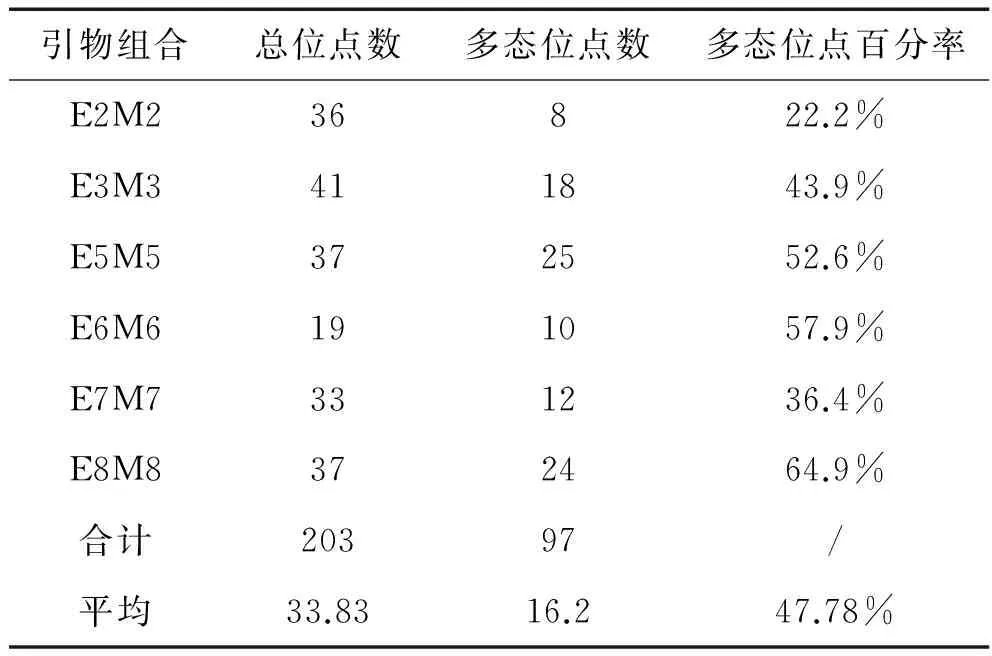
表3 六对AFLP引物组合的扩增
2.2 各群体遗传结构和遗传多样性
长肋日月贝4个群体内的遗传多样性参数见表4.结果表明,4个群体的群体Nei氏遗传多样性指数在0.0957~0.1 135之间,Shannon’s信息指数均在0.1 493~0.1 769之间,4个群体的遗传多样性均偏低.
陵水群体(LS)和三亚群体(SY)的多态性位点的比率均为40.89%,潭门群体(TM)的多态性位点比率最低,为34.4 8%.研究中没有发现群体特异性条带.

表4 长肋日月贝4个群体内遗传多样性参数
2.3 群体间遗传相似度和遗传距离
统计结果表明,BMJ群体和SY群体的遗传距离最大,为0.0162,遗传相似度为0.9840; BMJ群体和TM群体的遗传距离最小,为0.0049,遗传相似度为0.9952(表5).而LS和SY群体的遗传相似度为0.9925,遗传距离也较小.
对各群体内部的个体间遗传距离进行了分析,结果表明,BMJ、TM、LS和SY群体内部的遗传距离分别为0.1 217、0.1 065、0.1 212和0.1 179,均高于群体间的遗传距离.这表明野生长肋日月贝的遗传变异主要来自群体内部,群体间的遗传分化程度不大.

表5 长肋日月贝4个群体的群体间相似度和遗传距离矩阵
注:对角线以上数值为遗传相似度;对角线以下数值为遗传距离.

图2 长肋日月贝4个群体的UPGMA聚类图(标尺为遗传距离)
4个群体的聚类分析见图2.从图2中也可以看出,BMJ和TM群体的遗传距离最小,聚在一起;LS和SY群体聚在一起,表明遗传相似度较高.总的看来,4个群体的分化不明显,群体间的遗传相似度均较高,其差异明显处于种内地理群水平.
对4个群体的数据进行AMOVA分析,结果显示,种群间的分化系数(Fst)为0.06834,基因流水平(Nm)为3.4 08.同样说明长肋日月贝4个群体的变异主要来自群体内部,群体间的基因交流较为频繁,未形成明显分化.
3 讨论与结论
培育贝类新品种、新品系和保护现有的优良种质资源是目前摆在我国贝类育种方面的重要课题.海南近年来海域污染对生物多样性也造成了一定的影响[13].了解贝类新品种的遗传多样性状况,种群遗传结构,有助于建立清晰的亲体管理策略,做到对现有的种质资源进行合理、有效、可持续性地开发利用.
王雨等分析了野生长肋日月贝形态性状和重量性状的内在关系,结果表明,壳高与长肋日月贝的体重和闭壳肌重关系密切.在日月贝的人工育苗或者育种时,不需要测量壳长、壳宽、体重等其他形态和重量指标,只要选择壳高较大的个体,就可以快捷方便挑选出具有较大闭壳肌等主要食用部分的优良性状个体,便于亲本的选择[3].
目前,我国长肋日月贝的人工繁殖尚未大规模开展,天然捕捞到的长肋日月贝可以认定为野生种.本研究利用AFLP分子标记技术,采用6对引物,从多态位点、Nei氏遗传多样性指数、Shannon’s信息指数、遗传距离和遗传相似度等方面对中国海南4个野生群体进行了遗传多样性分析.结果表明,多态位点百分率最高的是LS和SY群体(均为40.89%),多态位点百分率最低的是TM群体(34.4 8%).研究中没有发现群体特异性条带.4 个野生群体之间的遗传相似度均高于0.98,说明4个群体之间的基因流动比较频繁,特别是SY和LS群体以及BMJ和TM群体之间的遗传相似度超过0.99,说明他们群体之间的基因流动更加频繁.
Hare等(1996)在对美洲牡蛎(Crassostreavirginica)的研究中发现,幼虫浮游生活时间较长的贝类,不同地理群体间会有一定幅度的基因流动[14].长肋日月贝从D形幼虫发育至眼点期幼虫需要11~15 d,从眼点期到附着完毕最多10 d[4],也就是说,长肋日月贝的浮游期最长可达25 d.而海南岛受到季风等的影响,周围海流复杂而多变,部分浮游幼体可随海流产生地理漂移,因而造成基因的流动,这可能是造成群体间遗传相似性较高的原因之一.
长肋日月贝4个群体的群体Nei氏遗传多样性指数在0.0957~0.1 135之间,Shannon’s信息指数均在0.1 493~0.1 769之间,4个群体的遗传多样性均偏低.据调查,近年来长肋日月贝的资源量越来越少[4],这可能是导致各群体遗传多样性偏低的另一个原因.较低的遗传多样性,使得种群结构趋向单一化,物种的适应能力也降低,大大增加种群面临灭绝的危险.
杂交是育种中最为简便、有效的途径之一.一般地说,不同群体之间,由于地理距离较远或者存在某些天然屏障,种群之间的某些基因存在表达差异.因此,不同种群之间进行物种杂交可以获得较大的杂种优势.有研究表明,3个野生种群的马氏珠母贝(Pinctadamartensii)在繁育过程中,杂交子一代的遗传多样性高于各种群自繁子一代[15].本研究表明,长肋日月贝的群体间遗传距离小于群体内部的遗传距离,因此,今后在长肋日月贝的育种方面,可以采用分子标记和优良性状的筛选(如壳高等)相结合的方法进行优质苗种的繁育.首先利用分子标记技术掌握不同地理群体之间个体的遗传多样性水平,进而尝试利用不同地理群体间具有优良性状的个体进行杂交,以提高后代的遗传多样性,获得较大的杂种优势.
今后,还需要对国内外长肋日月贝种质资源的遗传多样性进行评估,以期找出遗传距离较大的种群,进行引种杂交,提高本地种群的遗传多样性.另外,加强长肋日月贝的育种技术研究,并建立长肋日月贝的原种场和种质库,也是保护种质资源的重要措施.一方面可以维持长肋日月贝的遗传多样性,另外还能利用分子生物学手段选择杂合度较高的个体进行苗种繁殖,避免群体自繁引起遗传多样性的降低.
[1]王祯瑞.中国动物志[M].北京:中国科学出版社, 2002.
[2]王雨,叶乐,陈旭等.海南野生长肋日月贝形态性状与重量性状的通径分析[J].安徽农业科学, 2009, 37(8):3570-3572.
[3]朱彩艳,王雨,张殿昌,等.长肋日月贝的营养成分分析及评价[J]. 海洋科学, 2011,35(3):87-91
[4]王雨,叶乐,杨其彬,等.长肋日月贝个体发生观察及人工育苗初步试验[J].南方水产, 2009, 5(1):36-41.
[5]Belda C A, Del Norte A G C. Notes on the Induced Spawning and Larval Rearing of the Asian Moon Scallop,Amusiumpleuronectes(Linne), in the laboratory[J]. Aquaculture, 1988(72):173-179.
[6]Rice M A, Rheault R B, Perez M S, Perez V S. Experimental Culture and Particle Filtration by Asian Moon Scallops,Amusiumpleuronectes[J]. Asian Fisheries Science, 1994(7):179-185.
[7]Mahidol C, Na-Nakorn U, Sukmanomon S, et al. Mitochondrial DNA Diversity of the Asian Moon Scallop,Amusiumpleuronectes(Pectinidae), in Thailand[J]. Marine Biotechnology, 2007(9):352-359.
[8]赫崇波,丛林林,葛陇利,等.文蛤养殖群体和野生群体遗传多样性的AFLP分析[J].中国水产科学, 2008, 15(2):215-221.
[9]张东,隋正红,易恒,等.青岛裙带菜孢子体野生种群遗传多样性的AFLP分析[J].中国海洋大学学报, 2009, 39(4):664-668.
[10]许志强,葛家春,曹廷,等.江苏3个秀丽白虾野生群体遗传多样性的AFLP分析[J].江苏农业学报, 2009, 25(1):160-164.
[11]佟广香,匡友谊,尹家胜.野生哲罗鱼种质资源遗传多样性的AFLP分析[J].中国水产科学, 2009, 16(6):833-841.
[12]潘洁,包振民,赵洋,等.栉孔扇贝不同地理群体的遗传多样性分析[J].高技术通讯, 2002(12):78-82.
[13]贺立静,周述波,李晓梅,等.海南近岸海域污染现状及对生物多样性影响[J].琼州学院学报,2014, 21(5):99-103.
[14]Hare M P and Avise J C.. Molecular Genetic Analysis of a Stepped Multilocus Cline in the American Oyster (Crassostreavirginica) [J]. Evolution, 1996, 50(6):2305-2315.
[15]吕林兰,杜晓东,王嫣,等.马氏珠母贝3个野生种群及种群间杂交后代遗传多样性的ISSR分析[J].水生生物学报, 2008, 32(1):26-32.
(编校:何军民)
Genetic Diversity of Four Hainan WildAmusiumpleuronectesPopulations Revealed by AFLP Markers
ZHU Cai-yan1, JIANG Shi-gui1, YANG Qi-bin1,2, ZHAGN Dian-chang1, SU Tian-feng1, WANG Yu1,2
(1 South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, China;2 Tropical Fishery R&D Center, South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences,Sanya Hainan, 572018, China)
The genetic diversity ofAmusiumpleuronectesin China is not clear at present. By using AFLP markers, the current research investigated the genetic diversity of four wild populations (Baimajing (BMJ), Tanmen (TM), Lingshui (LS), Sanya (SY)) ofAmusiumpleuronectesin Hainan Province of China. A total of 203 bands were detected with 6 primers combinations, and there were 97 polymorphic loci among them (percentage:47.78%). The Nei’s gene diversity index and Shannon’s information index of the 4 populations were analyzed. Results showed that, Nei’s gene diversity index and Shannon’s information index of the 4 populations are among 0.0957~0.1 135 and 0.1 493~0.1 769 respectively, indicating low genetic diversity of the 4 populations. The ratio of polymorphic loci is high in LS and SY populations (both were 40.89%) and low in TM population (34.4 8%). The genetic distances between the 4 populations are not more than 0.0162 (max:SY-BMJ 0.0162; min:TM-BMJ 0.0049). No population-specific bands were investigated. The genetic distances within populations in BMJ, TM, LS and SY populations are 0.1 217, 0.1 065, 0.1 212 and 0.1 179, respectively. Result of AMOVA analysis showed Fst=0.06834 and Nm=3.4 08. It indicated that the genetic differentiations between populations are lower than those within populations. Some suggestions in germplasm conservation and breeding were discussed in this paper.
Amusiumpleuronectes; AFLP; genetic diversity
2016-03-09
海南省社会发展科技专项(2011SF014);国家科技基础平台建设项目(2005DKA30470-007);中央级公益性科研院所基本科研业务费专项资助项目(2007TS06)
朱彩艳(1979-),女,山东省青岛市人,中国水产科学院南海水产研究所助理研究员,硕士,研究方向为水产动物遗传育种研究.
王雨(1967-),女,湖南省衡阳市人,中国水产科学院南海水产研究所副研究员,研究方向为海水养殖与生态学.
S917
A
1008-6722(2016) 02-0099-06
10.1 3307/j.issn.1 008-6722.2 016.02.2 0
