“中科2号”海湾扇贝(Argopecten irradians)幼虫期壳长遗传力的估计
2016-10-20张守都张国范
张守都, 李 莉, 张国范
“中科2号”海湾扇贝(Argopecten irradians)幼虫期壳长遗传力的估计
张守都1, 李 莉2, 张国范2
(1. 青岛国家海洋科学研究中心, 山东青岛 266071; 2. 中国科学院海洋研究所, 山东青岛 266071)
基于全同胞组内相关法估计“中科2号”海湾扇贝()幼虫期壳长的遗传力。通过平衡巢式交配实验设计构建了7个半同胞和21个全同胞“中科2号”海湾扇贝家系。在家系幼虫期的第1天和第6天分别测定每个全同胞家系的30个幼虫个体的壳长。利用SAS 9.0 软件的一般线性模型(GLM)计算表型变量的原因方差组分, 估算壳长遗传力。分析结果显示, “中科2号”海湾扇贝幼虫1日龄和6日龄壳长的遗传力分别为0.51和0.58。检验显示两个日龄壳长遗传力的估计值均不显著(>0.05), 且6日龄雌性遗传方差组分大于雄性遗传方差组分, 说明雌性遗传方差组分随日龄增长可能存在较大母性效应。较大的加性遗传方差表明选择育种对于“中科2号”海湾扇贝的早期生长尚具有较大的改良潜力。
“中科2号”; 海湾扇贝(); 全同胞; 幼虫期; 遗传力
遗传力概念的提出距今已有70多年的历史, 它是反映亲本性状能够遗传给后代大小程度的指标, 在选择指数确定、选择反应预测、育种值估计、选择方法比较以及育种规划决策等方面, 均具有十分重要的参考作用[1]。遗传力的研究和应用在农作物、畜牧中早已广泛开展并取得了显著的研究成果[2-5]。近年来, 随着水产动物育种理论的进步, 贝类遗传力的研究也越来越多, 主要包括牡蛎、扇贝、贻贝、合浦珠母贝以及鲍鱼等的遗传力研究[6-10]。由于贝类也同其他水产动物一样, 具有高繁殖力这一特性, 在遗传力估计时多采用同胞分析进行估计。同胞分析所采用的方差组分估计方法主要包括方差分析法、最小范数二次无偏估计和极大似然法。
海湾扇贝自1982年引入中国以后[11], 因其生长速度快、养殖效益高, 深受广大养殖户的欢迎, 并在世界上首次形成了海湾扇贝养殖产业[12], 且发展成为中国海水养殖的支柱产业之一[13], 为中国沿海经济的发展做出了巨大的贡献。然而, 自从20世纪90年代以来, 海湾扇贝在整个育苗与养殖产业中逐步暴露出现一些明显的问题: 生长速度减缓、商品贝体个体小型化[14]; 育苗和养成过程中病害频发、死亡率明显升高、近交衰退严重、抗逆能力减弱[15]; 主要养殖群体的遗传多样性严重丢失[16-17]等。为此中国贝类育种工作者持续开展了海湾扇贝的品种选育工作。“中科2号”海湾扇贝就是在此背景下由中国科学院海洋研究所经过多年定向选育培育出的海湾扇贝新品种。“中科2号”海湾扇贝壳色鲜艳, 生长性状优良, 具有很好的养殖潜力[18], 掌握该新品种的遗传特性具有重要的科学和应用意义。本研究通过平衡巢式设计的全同胞组内相关法, 建立全同胞家系, 通过方差分析法, 对“中科2号”海湾扇贝幼虫期壳长的遗传力进行估计, 旨为“中科2号”海湾扇贝的推广养殖和继续选择育种提供必要的理论依据和技术参考。
1 材料与方法
1.1 亲贝的获取和培养
实验所用亲贝为中国科学院海洋研究所培育的“中科2号”海湾扇贝养殖群体, 所有亲贝均在青岛市崂东海珍品良种培育有限公司育苗车间进行性腺促熟, 培育方法参照张福绥[11]等, 整个促熟过程遵循无公害食品海湾扇贝养殖技术规范NY/T 5063— 2001。
1.2 全同胞家系的建立
2012年5月上旬, 通过巢式平衡设计建立了7个父系半同胞家系和21个全同胞家系。从所培养亲贝群体中随机挑选30只性腺发育至第四期[19]的个体在空气中阴干30 min, 然后对每只扇贝注射五羟色胺[8], 并将每一只处理好的扇贝逐一各放入一个加有22℃沙滤海水的1L烧杯中待产, 大约30 min后所处理扇贝会首先排放精子, 收集质量好的精子备用, 接下来在排卵前会出现一个10 min左右的暂停期, 此时将扇贝用新鲜海水冲洗几遍后放回烧杯中, 待其进一步排放卵子。为了进一步防止自体受精, 第一次收集卵子弃之不用, 接下来每次收集卵子均用400目筛绢洗卵, 尽量减少自身精子污染机会, 然后将所收集卵子在显微镜下检查, 选取未被污染的(即未自体受精)卵子备用, 实验分别收集到28只扇贝足量的精子和未自体受精的卵子。
按照巢式实验设计, 以其中7只扇贝为父本, 其余为母本, 每只父本扇贝的精子分别和3只母本扇贝的卵子配对交配, 共构建起7个父系半同胞家系和21个全同胞家系。
1.3 幼虫培养
在受精卵孵化24 h后发育成为D形幼虫, 用300目的筛绢对D形幼虫进行选优, 幼虫的起始培养密度被设置为 10个/mL, 各实验组分别在80 L的白塑桶23℃沙滤海水中培养, 并尽可能使各组环境条件保持一致。幼虫的前期饵料为金藻, 后期为金藻、扁藻和小硅藻等。投饵量和投饵次数随幼虫增长而增加, 确保每天全换水1次。10 d左右时幼虫开始出现眼点。当30%左右的幼虫出现眼点时开始投放附着基(聚乙烯网片), 每个塑料桶中投放网片数依幼虫多少而定。
1.4 数据测量
对各个家系1日龄和6日龄幼虫的壳长在显微镜下用目微尺进行测量, 每个家系测量个体为30个。
1.5 统计分析
由于实验设计采取3水平经典巢式平衡设计, 即雄性亲本内每个雌性亲本的后代数目(1)、雄性亲本间每个雌性亲本的后代数目(2)和每个雄性亲本后代数目都相等(3), 各个亲本后代数不需要加权校正计算。各种数据通过SAS9.0软件的GLM (General Linear Model)过程实现运算, 差异的显著性都设置为<0.05。全同胞资料表型变量的方差组分见表1。

表1 表型变量组成的方差分析
注:分别为后代个体总数、雄性亲本数、雌性亲本数;: 父系半同胞方差;: 母系半同胞方差;: 全同胞个体间方差
父本和母本对后代表型性状均值影响的方差分析通过下面的统计模型进行:
Y=+α+β+e
其中,Y是第父本与第母本交配而得的第个子女的观察值,为共同均数,α为第父本的效应,β为与第父本交配的第母本的效应,e是归因于个体的不可控环境方差和遗传离差。
各部分方差组分的计算公式如下:
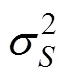
1.6 遗传力的计算及显著性检验
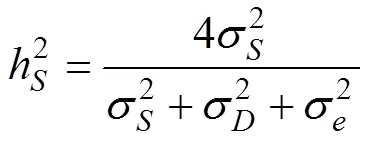
对各个计算所得遗传力进行显著性的检验:
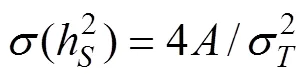

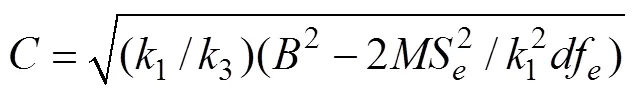
2 结果与分析
2.1 “中科2号”海湾扇贝幼虫期的壳长统计数据
“中科2号”海湾扇贝1日龄和6日龄幼虫期的壳长平均数、标准差和统计个数等见表2。

表2 “中科2号”海湾扇贝幼虫期壳长的基本统计参数
2.2 “中科2号”海湾扇贝幼虫期壳长数据的方差分析
“中科2号”海湾扇贝幼虫期壳长数据的方差分析见表3, 结果表明在幼虫期1日龄和6日龄, 父本间均表现差异显著(<0.05), 而母本间均表现差异极显著(<0.01)。

表3 “中科2号”海湾扇贝幼虫期表型变量组成的方差分析
注: *表示差异显著(< 0.05), **表示差异极显著(< 0.01)
2.3 “中科2号”海湾扇贝幼虫期壳长的遗传力估计
各个原因方差组分构成见表4, 其中在6日龄雌性亲本的方差要大于雄性亲本的方差, 说明同1日龄相比较, 在此时已经体现出较为明显的母性效应。
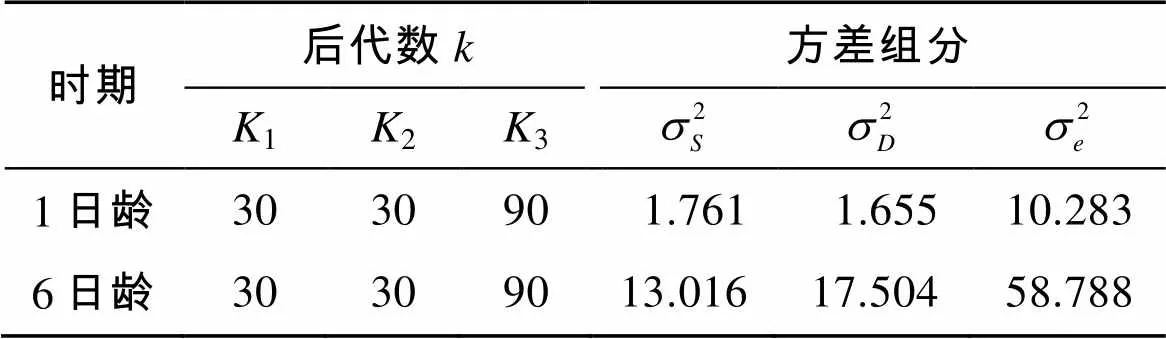
表4 “中科2号”海湾扇贝幼虫期方差组分估计值
根据表4的各个方差组分估计值, 计算了“中科2号”海湾扇贝幼虫期1日龄和6日龄壳长的遗传力并进行了检验(表5)。其中1日龄壳长的遗传力为0.499~0.514, 6日龄壳长的遗传力为0.583~0.784。
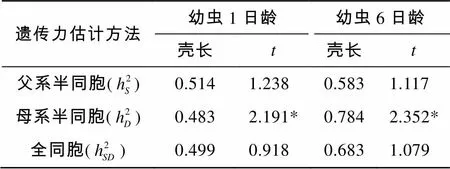
表5 “中科2号”海湾扇贝幼虫期壳长遗传力及其t检验
注: *表示差异显著(< 0.05)
对所计算各个遗传力按上述公式进行了检验结果表明: 以母系半同胞资料估计的遗传力均达到显著水平(<0.05), 而以父系半同胞资料和全同胞资料估计的遗传均未达到显著水平(>0.05)。
3 讨论
遗传力是选择育种中最常用到的指标, 遗传力的大小能直接反映可供选择的加性遗传方差占总体方差的比例, 即可以预测选择育种的效果并指导采取合适的选择育种的方法。当某个群体遗传力大于0.4时一般将其划分为高遗传力, 则其性状选择有较大的可靠性, 适合用于个体或群体表型选择法进行选择育种, 而当遗传力低于0.2时将其划分为低遗传力, 认为该性状选择效果较差, 适合用于家系选择或提高选择的强度等[20]。在很多贝类的选择育种项目中均已经开展了遗传力的研究。Loose[21]估计美洲牡蛎()7日龄、14日龄和21日龄幼虫壳长的半同胞遗传力分别为0.44、0.40和0.55。Lannan等[22]利用11~15个全同胞家系估计了18月龄长牡蛎壳高、总重、壳重、肉重的广义遗传力, 依次为0.15、0.33、0.32、0.37。Toro 等[23]报道了欧洲牡蛎()24个家系生长率的遗传力估计值从0.112到0.243。Mallet等[24]利用同胞分析技术估计了加拿大贻贝()幼虫壳长的遗传力为0.11。邓岳文[10]利用巢式平衡设计估计皱纹盘鲍10日龄、20日龄和30日龄壳长的半同胞遗传力从0.232 到0.365、全同胞遗传力从0.175 到0.747、平均遗传力从0.202 到0.489; 壳宽的半同胞遗传力从0.181到0.316、全同胞遗传力从0.104到0.604、平均遗传力从0.142到0.428。王庆志等[25]估计了长牡蛎5到25日龄幼虫壳长及壳高的狭义遗传力, 其遗传力估计值分别为从0.139到0.814 和从0.161到0.771。郑怀平[26]利用巢式平衡设计估计了“中科红”海湾扇贝在1日龄、10日龄和40日龄壳长的半同胞遗传力从0.439到0.639, 全同胞遗传力从0.325到0.432, 半同胞与全同胞的平均遗传力从0.413到0.536; 壳高的半同胞遗传力从0.433到0.539, 全同胞遗传力从0.268到0.433, 半同胞与全同胞的平均遗传力从0.404到0.433。梁冰冰[27]等采用部分因子设计构建了23个全同胞家系, 应用单性状及两性状动物模型对文蛤稚贝阶段的生长性状进行遗传力分析, 结果表明, 附着变态期文蛤壳长和壳高的遗传力估计值为0.11和0.34, 遗传力研究已经成为贝类遗传育种一项必不可少的基本工作。
本研究中估计的“中科2号”海湾扇贝1日龄和6日龄的狭义遗传力分别为从0.483到0.514和从0.583到0.784。该遗传力估计相对其他贝类遗传力研究结果偏大, Davis等[28]发现受过选择的美洲牡蛎()幼虫壳长的半同胞遗传力为0.44, 而未选择群体为0.14。虽然本研究未设置未受选择群体作为对照, 但导致本研究中遗传力较大的原因亦有可能是由长期选择引起。本研究结果中(表5)基于1日龄父系半同胞资料估计的遗传力(0.514)要大于基于1日龄全同胞资料估计的遗传力(0.499)。而基于6日龄父系半同胞资料估计的遗传力(0.583)小于基于6日龄全同胞资料估计的遗传力(0.683)。以上结果表明基于半同胞资料和全同胞估计的资料估计的遗传力结果并不完全一致。一般认为, 基于全同胞资料估计的遗传力, 由于包含显性效应和母性效应, 较真值偏大。但是本研究在1日龄基于父系半同胞资料估计的遗传力反而大于基于母系半同胞资料估计的遗传力, 可能与该群体经过多年选育而亲本间有一定的亲缘关系, 不同父系间全同胞个体的一致性增加, 组间变异或下降, 导致基于全同胞资料所估计遗传力偏低。Kanis 等[29]发现虹鳟()母系半同胞遗传力受到母性效应的显著影响。在本研究中6日龄的母系半同胞遗传力相比1日龄母系半同胞遗传力有明显增大趋势且大于基于6日龄父系半同胞资料估计的遗传力, 这跟海湾扇贝中一开始并不表现出较强母性效应相一致[30], 而据此推断在后期遗传力的值还会随着母性效应的变化而变化。虽然本研究采用完全平衡巢式设计和父系半同胞组内相关分析法, 克服了用全同胞资料估计遗传力偏大的缺点, 但该研究结果的母性半同胞遗传力值还是受到母性效应的明显影响。检验显示在“中科2号”海湾扇贝1日龄和6日龄母系半同胞遗传力均达到显著水平(<0.05), 而父系半同胞遗传力虽未达到显著水平(>0.05), 但因未受母性效应影响仍较母系半同胞遗传力为准, 所以本研究中认为父系半同胞估计的遗传力是“中科2号”海湾扇贝幼虫期壳长遗传力的无偏估计。
本研究结果表明, 基于“中科2号”海湾扇贝幼虫期壳长的加性遗传方差较大, 个体选择育种对于“中科2号”海湾扇贝幼虫期的生长仍具有较大的改良潜力。
致谢: 感谢青岛崂东海珍品良种培育有限公司为本研究提供了实验所用的场地以及所有设施。
[1] 盛志廉, 陈瑶生. 数量遗传学[M]. 北京: 科学出版社, 2001. Sheng Zhilian, Chen Yaoshen. Quantitative Genetics[M]. Beijing: Science Press, 2001.
[2] 吴景锋. 我国玉米杂交种发展的主要历程、差距和对策[J]. 玉米科学, 1995, 3(1): 1-5. Wu Jingfeng. Main development stages, disparity and development polices on maize hybrids in China[J]. J Maize Sci, 1995, 3(1): 1-5.
[3] Kallo. Vegetable breeding[M]. Florida: CRC Press, 1988, 2: 176 -179.
[4] Wehner T C. Estimates of heritabilities and variance component for low-temperature germination ability in cucumber[J]. J Amer Soc Hort Sci, 1984, 109(5): 664- 666.
[5] 吴珍芳, 陈文广. 猪胴体和肉质性状遗传力及其遗传相关[J]. 华北农学报, 2000, 15(增刊): 46-50. Wu Zhenfang, Chen Wenguang. Heritabilities and genetic correlations of pig carcassand meat quality traits[J]. Acta Agri Boreali-Sinica, 2000, 15(Suppl.): 46-50.
[6] Lannan J E. Estimating heritability and predicting response to selection for the Pacific oyster,[J]. Proc Natl Shellfish Assoc, 1972, 62: 62-66.
[7] Stromgren T, Nielsen M V. Heritability of growth in larvae and juveniles of[J]. Aquaculture, 1989, 80: 1-6.
[8] Crenshaw J W, Heffernan P B, Walker R L. Heritability of growth rate in the southern Bay scallop,[J]. J Shellfish Res, 1991, 10: 55-63.
[9] 何毛贤, 姜卫国. 合浦珠母贝遗传育种研究进展[J]. 海洋湖沼通报, 2000, 1: 75-82. He Maoxian, Jiang Weiguo. The advance of genetics and breeding in(D.)[J]. Trans Oceanol Limnol, 2000, 1: 75-82.
[10] 邓岳文. 皱纹盘鲍数量性状遗传与育种[D]. 青岛: 中国科学院海洋研究所, 2005. Deng Yuewen. Studies on quantitative genetics and breeding of Pacific abalone[D]. Qingdao: IOCAS, 2005.
[11] 张福绥, 何义朝, 刘强生, 等. 海湾扇贝的引种、育苗与试养[J]. 海洋与湖沼, 1986, 17(5): 367-374. Zhang Fusui, He Yichao, Liu Qiangsheng, et al. A report on the introduction, spat-rearing and experimental culture of bay scallopLamarck[J]. Oceanol Limnol Sin, 1986, 17(5): 367-374.
[12] 张福绥. 中国海湾扇贝养殖业的发展[J]. 海洋科学, 1992, 16(4): 1-4. Zhang Fusui. The development of bay scallop aquaculture industry in China[J]. Marine Sciences, 1992, 16(4): 1-4.
[13] 张福绥. 海湾扇贝引进中国10周年[J]. 齐鲁渔业, 1993, 5: 9-12. Zhang Fusui. The 10th anniversary for introduction bay scallop to China[J]. Shandong Fisheries, 1993, 5: 9-12.
[14] 张福绥, 何义朝, 齐铃欣, 等. 海湾扇贝引种复状研究[J]. 海洋与湖沼, 1997, 28(2): 146-152. Zhang Fusui, He Yichao, Qi Lingxin, et al. Studies on the restoration of cultured bay scallopthrough reintroduction of broodstock[J]. Oceanol Limnol Sin, 1997, 28(2): 146-152.
[15] 张国范, 李霞, 薛真福. 我国养殖贝类大规模死亡的原因分析及防治对策[J]. 中国水产, 1999, 9: 34-39. Zhang Guofan, Li Xia, Xue Zhenfu. Cause analysis and strategies of reoperation for large-scale death of cultured shellfish in China[J]. China Fisheries, 1999, 9: 34-39.
[16] Blake S G, Blake N J, Oesterling M J, et al. Genetic divergence and loss of diversity in two cultured populations of the bay scallop,(Lamark, 1819)[J]. J Shellfish Res, 1997, 16: 55–58.
[17] Wang L L, Zhang H, Song L S, et al. Loss of allele diversity in introduced populations of the hermaphroditic bay scallop[J]. Aquaculture, 2007, 271: 252-259.
[18] 黄太寿, 李巍. 2011年水产新品种推广指南[M]. 北京: 中国农业出版社, 2001. Huang Taishou, Li Wei. Guidelines for new aquatic varieties extension in 2011[M]. Beijing: China Agriculture Press, 2001.
[19] Sastry A N. Reproduction of the bay scallopLamarck. Influence of temperature on maturation and spawning[J]. Biol Bull, 1963, 441: 146-153.
[20] 赵存发, 高佃平, 李金泉, 等. 内蒙古白绒山羊体重性状遗传力的估计[J]. 畜牧与饲料科学, 2010, 6: 73-74.Zhao Cunfa, Gao Dianping, Li Jinquan, et al. Heritability of body weight for white cashmere goats in inner Mongolia[J]. Ani Husb Feed Sci, 2010, 6: 73-74.
[21] Loose E. Influence of heredity on larvae and spat growth in[J]. Proc World Maricult. Soc, 1978, 9: 101-108.
[22] Lannan J E. Estimating heritability and predicting response to selection for the Pacific oyster[J]. Proc Natl Shellfish Assoc, 1972, 62: 62-66.
[23] Toro J E, Newkirk G F. Divergent selection for growth rate in the European oyster: response to selection and estimation of genetic parameters[J]. Mar Ecol Prog Ser, 1990, 62; 219-227.
[24] Mallet A L, Freeman K R, Dickie L M. The genetics of production characters in the blue mussel. I. A preliminary analysis[J]. Aquaculture, 1986, 57: 133-140.
[25] 王庆志, 李琪, 刘士凯, 等. 长牡蛎幼体生长性状的遗传力及其相关性分析[J]. 中国水产科学, 2009, 16(5): 736-743. Wang Qingzhi, Li Qi, Liu Shikai, et al. Estimates of heritabilities and genetic correlations for growth inlarve[J]. J Fishery Sci China, 2009, 16(5): 736-743.
[26] 郑怀平. 海湾扇贝两个养殖群体数量性状及壳色遗传学研究[D]. 青岛: 中国科学院海洋研究所, 2007. Zheng Huaiping. Genetic studies of quantitative traits and shell colors in two hatchery stocks of the bay scallop,[D]. Qingdao: IOCAS, 2007.
[27] 梁冰冰, 王鸿霞. 文蛤早期发育阶段生长形状遗传参数的估计[J]. 海洋科学, 2016, 40(3): 40-46. Liang Bingbing, Wang Hongxia. Genetic parameter estimation for growth traits in the early life history of[J]. Marine Sciences, 2016, 40(3): 40-46.
[28] Davis C V. Estimation of narrow-sense heritability for larval and juvenile growth traits in selected and unselected sub-lines of eastern oyster,[J]. J Shellfish Res, 2000, 19(1): 613.
[29] Kanis G, Refstie T, Gjedrem T. A genetic analysis of eg, alevin and fry mortality in salmon (), sea trout () and rainbow trout ()[J]. Aquaculture, 1976, 8: 259-268.
[30] Zhang H B, Liu X, Zhang G F, Wang C D. Growth and survival of reciprocal crosses between two bay scallops,Say andLamarck[J]. Aquaculture, 2007, 272: 88-93.
Heritability of larval shell length for Zhongke No. 2 bay scallop ()
ZHANG Shou-du1, LI Li2, ZHANG Guo-fan2
(1. National Oceanographic Center, Qingdao 266071, China; 2. Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China)Received:Apr.6, 2015
; Zhongke No. 2; full-sib; larvae; heritability
In this study, we estimated the heritability of the shell length of the Zhongke No. 2 bay scallop,by performing an intra-group correlation of full-sibs at 1 day and 6 days after hatching. Each sire was mated with three dams in a balanced nested design, and 7 half-sib groups and 21 full-sib families were produced in total. We calculated the causal components of the phenotypic variance using the general linear model (GLM) procedure and SAS 9.0 software. The results showed that the heritabilities of shell length of 1 day and 6 days were 0.51 and 0.58, respectively. The heritabilities of both these stages failed to reach their prominent level bycheckout. Maternal genetic variance was significantly greater than paternal genetic variance in 6 days, which indicates the existence of greater maternal effect. The estimated heritability indicated great additive variation in growth at the larval stage. This additive variation suggests a reasonable potential for the improvement of larval growth of the Zhongke No. 2 bay scallop for further selective breeding.
S917.3
A
1000-3096(2016)07-0054-06
10.11759/hykx20130406002
2015-04-06;
2015-09-24;
国家高技术研究发展计划(863计划, 2012AA10A410); 国家重点基础研究发展计划资助(973计划项目, 2010CB126401); 贝类产业技术体系(CARS-48); 泰山学者及泰山攀登计划
[Foundation: National High Technology Research and Development Program(863 Program, no. 2012AA10A410); National Basic Research Program of China (973 Program, no. 2010CB126401); Modern Agro-industry Technology Research System (CARS-48); Taishan Scholars Climbing Program of Shandong and Oversea Taishan Scholar Program of Shandong]
张守都(1984-), 男, 山东日照人, 博士研究生, 主要从事贝类遗传育种研究, 电话: 0532-82898712, E-mail: shouduzhang@163.com;张国范,通信作者, E-mail: gfzhang@qdio.ac.cn
(本文编辑: 张培新)
