西藏嵩草ISSR-PCR反应体系优化研究
2016-09-19胡延萍
胡延萍, 包 蕊, 王 莉, 石 琳,2, 李 毅*
( 1. 中国科学院西北高原生物研究所, 西宁 810008; 2. 中国科学院大学, 北京 100049 )
西藏嵩草ISSR-PCR反应体系优化研究
胡延萍1, 包蕊1, 王莉1, 石琳1,2, 李毅1*
( 1. 中国科学院西北高原生物研究所, 西宁 810008; 2. 中国科学院大学, 北京 100049 )
西藏嵩草(Kobresiatibetica)为莎草科嵩草属多年生草本,根状茎短,秆密集丛生。生于海拔2 550~4 950 m的河滩地、湿润草地、高山灌丛草甸,分布于甘肃、青海、四川西部、西藏东部,其根系发达,喜湿,耐寒,生活力强。西藏嵩草繁殖以营养繁殖为主、有性繁殖为辅,其茎叶茂盛,有较高的营养价值,产草量高,是青藏高原夏、秋两季的主要放牧饲草。该研究以青藏高原高寒沼泽化草甸的建群种和优势种——西藏嵩草为材料,对影响其ISSR-PCR反应的因素(Mg2+、TaqDNA聚合酶、dNTP、引物和模板DNA)进行5因素4水平的正交试验,以确定西藏嵩草ISSR分析的最佳反应体系,并筛选适宜的ISSR引物及各引物的最佳退火温度。结果表明:建立了适宜于西藏嵩草ISSR-PCR反应的最佳体系为20 μL反应液中包括10 × PCR buffer 2 μL、1.5 mmol·L-1Mg2+、1.0 UTaqDNA聚合酶、0.100 mmol·L-1dNTP、0.3 μmol·L-1引物和30~40 ng DNA模板;同时从100条ISSR引物中筛选出了扩增结果清晰、稳定的12条引物,各引物的最佳退火温度为48.0~53.2 ℃(引物不同,其最佳退火温度也不同);西藏嵩草ISSR-PCR适宜的扩增程序为首先预变性94 ℃ 5 min,然后变性94 ℃ 20 s、复性48.0~53.2 ℃ 1 min、延伸72 ℃ 80 s、38个循环,最后72 ℃延伸6 min。体系稳定性验证结果表明,该体系在西藏嵩草其他样品中所得条带清晰且多态性丰富,为后续西藏嵩草的遗传多样性分析和优良牧草种质资源筛选研究奠定了基础。该研究结果对以西藏嵩草为优势种的高寒沼泽化草甸研究及湿地生态系统的修复和保护具有重要意义。
西藏嵩草, 莎草科, ISSR-PCR, 反应体系, 正交设计
西藏嵩草(Kobresiatibetica),为莎草科(Cyperaceae)嵩草属(KobresiaWilld.)多年生草本。根状茎短。秆密集丛生,较粗壮,高5~15 cm,直径1~2 mm,有钝棱或圆柱形,具条纹,基部具浅褐色至深褐色枯死叶鞘。叶短于秆,丝状,宽约1 mm,边缘席卷。生于海拔2 550~4 950 m的河滩地、湿润草地、高山灌丛草甸,分布于甘肃、青海、四川西部、西藏东部,其根系发达,喜湿,耐寒,生活力强(中国科学院中国植物志编辑委员会, 2000; 张胜邦和卢学峰, 2012)。西藏嵩草的繁殖以营养繁殖为主,有性繁殖为辅,其茎叶茂盛、有较高的营养价值,产草量高,是青藏高原夏、秋两季的主要放牧饲草(邓德山等, 1995; 马玉寿和徐海峰, 2013)。以西藏嵩草为优势种形成的高寒沼泽化草甸,具有典型的高原湿地生态特征,是青藏高原高寒草甸类草地的组成部分,其独特的水源涵养功能,在维持青藏高原生态系统平衡及保护生物多样性等方面发挥着重要作用;其泥炭的形成与储存对降低大气中温室气体和稳定全球变化有着重要意义(周兴民, 2001; 李春秀和孙海松, 2009;田昆等, 2009)。
加拿大蒙特利尔大学Zietkiewicz et al(1994)创建了基于PCR的分子标记Inter-simple Sequence Repeats (ISSR)。该标记操作简单、灵敏度高、稳定性好而且多态性丰富(Nagaoka & Ogihara, 1997; Devarumath et al, 2002)。鉴于ISSR的优点,现已在牧草遗传结构分析、基因作图与定位、亲缘关系及种质资源筛选与育种等方面应用广泛(Li et al, 2013; 李钰莹和董宽虎, 2014)。本研究设计正交试验,优化影响青藏高原高寒草甸优势种西藏嵩草ISSR的主要因子(如TaqDNA polymerase、Mg2+、dNTP、引物和DNA模板),旨在建立适合西藏嵩草ISSR的稳定性及重复性好,多态性丰富的最佳反应体系,为进一步开展ISSR标记在西藏嵩草遗传多样性及种质资源筛选等方面的研究提供理论依据与遗传基础。
1 材料与方法
1.1 材料
2013年7月赴青海省海北藏族自治州祁连县境内的石棉矿(38°240′43.8″ N, 99°16′37.1″ E)地区采集西藏嵩草叶片材料,经中国科学院西北高原生物研究所卢学峰研究员鉴定为西藏嵩草(Kobresiatibetica),凭证标本放置于青藏高原生物标本馆(HNWP,标本号2013003)。
1.2 试剂
dNTP、TaqDNA 聚合酶、100 bp和200 bp DNA molecular marker购于TaKaRa公司。所用引物(Set#9,由加拿大哥伦比亚大学University of British Columbia提供)委托上海生工合成。
1.3 方法
1.3.1 提取DNA与质量检测基因组DNA提取采用Doyle JJ & Doyle JL(1987)改进的CTAB法。NanoDrop 2000c微量紫外分光光度计(Thermo Scientific)检测DNA的纯度和浓度,其OD260/280值均在2.04~2.12之间,浓度在95~450 ng·μL-1之间,满足ISSR-PCR对DNA模板质量的要求。
1.3.2 ISSR-PCR反应体系的优化对TaqDNA 聚合酶、Mg2+、dNTP、Primer及DNA等进行5个因素4种不同梯度的L16(45)正交试验筛选(表1)。优化试验选用UBC855(其序列为5′-ACACACACACACACACYT-3′)引物。每个体系还含有2 μL的10×PCR buffer,用超纯水补至20 μL。PCR扩增反应在C1000 TouchTMThermal Cycler型 (Bio-Rad) PCR仪上进行。扩增程序为预变性94 ℃ 5 min;变性94 ℃ 20 s,复性50 ℃ 1 min,延伸72 ℃ 80 s, 38 Cycles;72 ℃ 6 min延伸;并于4 ℃保存。PCR产物于水平板琼脂糖凝胶(浓度为1.2%)上电泳,使用凝胶成像系统(ChemiDocTMMP, Bio-Rad)拍照的方法记录实验结果。
1.3.3 引物筛选及其最佳复性温度优化在优化后的PCR体系上,从100条引物中筛选出扩增条带丰富且清晰、稳定性和多态性好的引物。以引物UBC855(Tm=52 ℃)为例,在其Tm值上下浮动4 ℃,设置8个温度梯度(48.0、48.5、49.6、51.2、53.1、54.7、55.6、56.0 ℃),其它反应程序同前文正交试验的扩增程序,筛选出该引物的最佳退火温度。
1.3.4 ISSR-PCR体系的稳定性验证用引物UBC855对西藏嵩草不同材料进行ISSR-PCR扩增,进行ISSR-PCR扩增体系的稳定性检测试验。
1.4 统计与分析
采用正交设计直观分析方法评分实验结果,按照扩增条带数量丰富度、清晰度及背景深浅的标准进行打分,条带数量丰富、清晰且背景低的为16分,最差的为1分。用DPS 7.05软件进行数据统计分析(唐启义等, 2006)。
2 结果与分析
2.1 西藏嵩草正交设计数据分析
图1是西藏嵩草ISSR-PCR正交实验的结果,处理12扩增片段清晰,重复性好,亮度适中。16个处理的直观打分结果如表1。

表 1 西藏嵩草正交试验 (5因素4水平)
表2为方差分析统计结果,据F值,各因素对西藏嵩草ISSR-PCR的影响程度由强到弱依次为Mg2+、TaqDNA聚合酶、引物、模板DNA、dNTP,且每个因素的影响均达到了极显著水平(P<0.01)。因此,为确定各个因素的最佳水平,需对5个因素分别进行因素内水平间的Duncan多重比较(表3)。
Mg2+是影响西藏嵩草ISSR-PCR反应最大的因素。当Mg2+浓度在0.9~1.5 mmol·L-1之间时,随Mg2+浓度增加,其评分结果均值呈增大趋势;当Mg2+浓度达1.8 mmol·L-1时,结果均值有减小趋势(图2)。Mg2+浓度为0.9~1.2 mmol·L-1,扩增条带丰度低且淡弱;使用1.5~1.8 mmol·L-1的Mg2+时,所得条带丰度高且清晰明亮,特别是1.5 mmol·L-1时,所得条带是实验中最清晰明亮的(处理12)。因此,1.5 mmol·L-1是Mg2+最适宜的浓度。
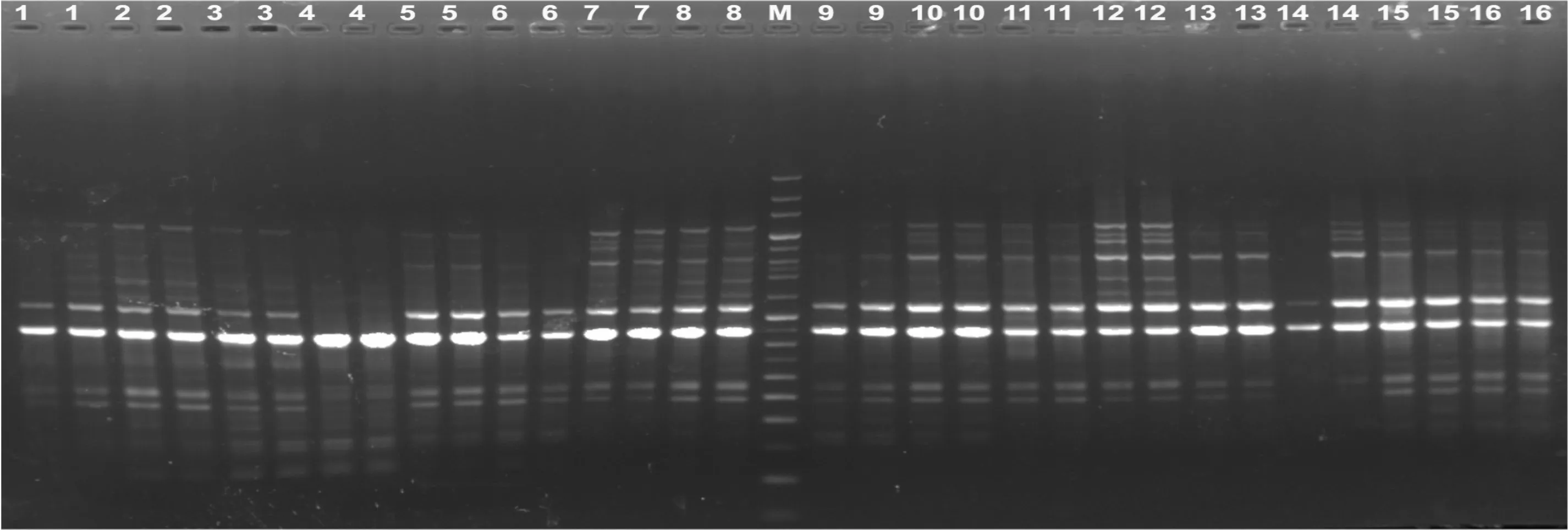
图 1 西藏嵩草ISSR-PCR正交设计电泳结果(引物UBC855) 1-16. 处理编号(表1); M. 100 bp+200 bp标准分子量参照物。Fig. 1 Electrophoresis results of ISSR-PCR in Kobresia tibetica with orthogonal design (Primer UBC855) 1-16. Treatment numbers as showed in Table 1; M. 100 bp+200 bp molecular ladder.

表 2 不同因素对西藏嵩草ISSR-PCR影响的方差分析
注: **差异极显著(P<0.01)。
Note: **Highly significant differences(P<0.01).

图 2 Mg2+浓度与评分结果均值间的关系 Fig. 2 Relationship between Mg2+ concentration and mean grading results
TaqDNA聚合酶对西藏嵩草ISSR反应的影响仅次于Mg2+(表2),评分结果均值随酶量的增加呈增大趋势(图3)。当TaqDNA聚合酶量为0.4~0.6 U时,扩增条带少;TaqDNA聚合酶量为0.8~1.0 U时,所得条带逐渐增多。由Duncan比较(表3)结果可知,仅水平3与4差异不显著,其它水平间均差异显著。综上结果及酶量的经济性原则,确定水平4(1.0 U·20 μL-1)是TaqDNA聚合酶的最佳用量。
对西藏嵩草ISSR-PCR影响较大的另一因素是引物浓度,引物浓度低时,扩增效率低;引物浓度偏高,则会导致错配、非特异性扩增及引物二聚体生成等可能性(图4),对不同分子量大小片段的扩增,引物浓度具有一定的选择性。由图1可知,引物浓度为0.3~0.4 μmol·L-1时,小于400 bp的片段未出现,而大于1 200 bp的片段被清晰明显扩增出来;当浓度增加到0.6 μmol·L-1时(处理4、6、9、15),大于1 200 bp的条带少且弱,出现了小于400 bp的片段。说明低浓度引物对大片段条带的扩增有利,高浓度引物则有利于小片段条带扩增。多重比较表明,水平1与水平2和3差异不显著,水平1与水平4差异显著。确定引物的适宜浓度为0.3 μmol·L-1。
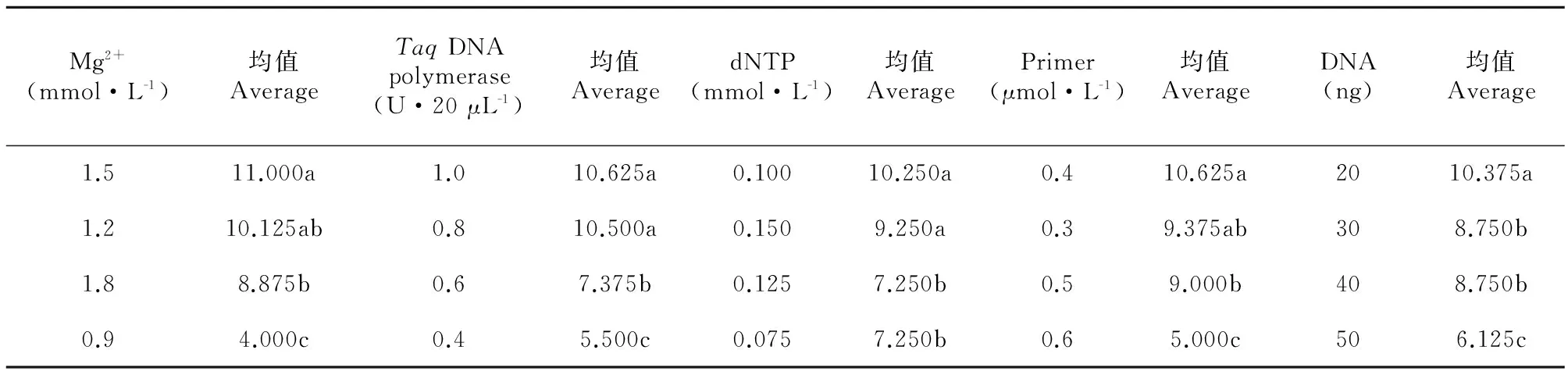
表 3 ISSR-PCR各因素水平间Duncan比较
注: 0.05显著水平,不同字母表示处理间差异显著,字母相同则表示不存在显著差异。
Note: Data with different letters differ significantly while with the same letters not significantly at 0.05 level.
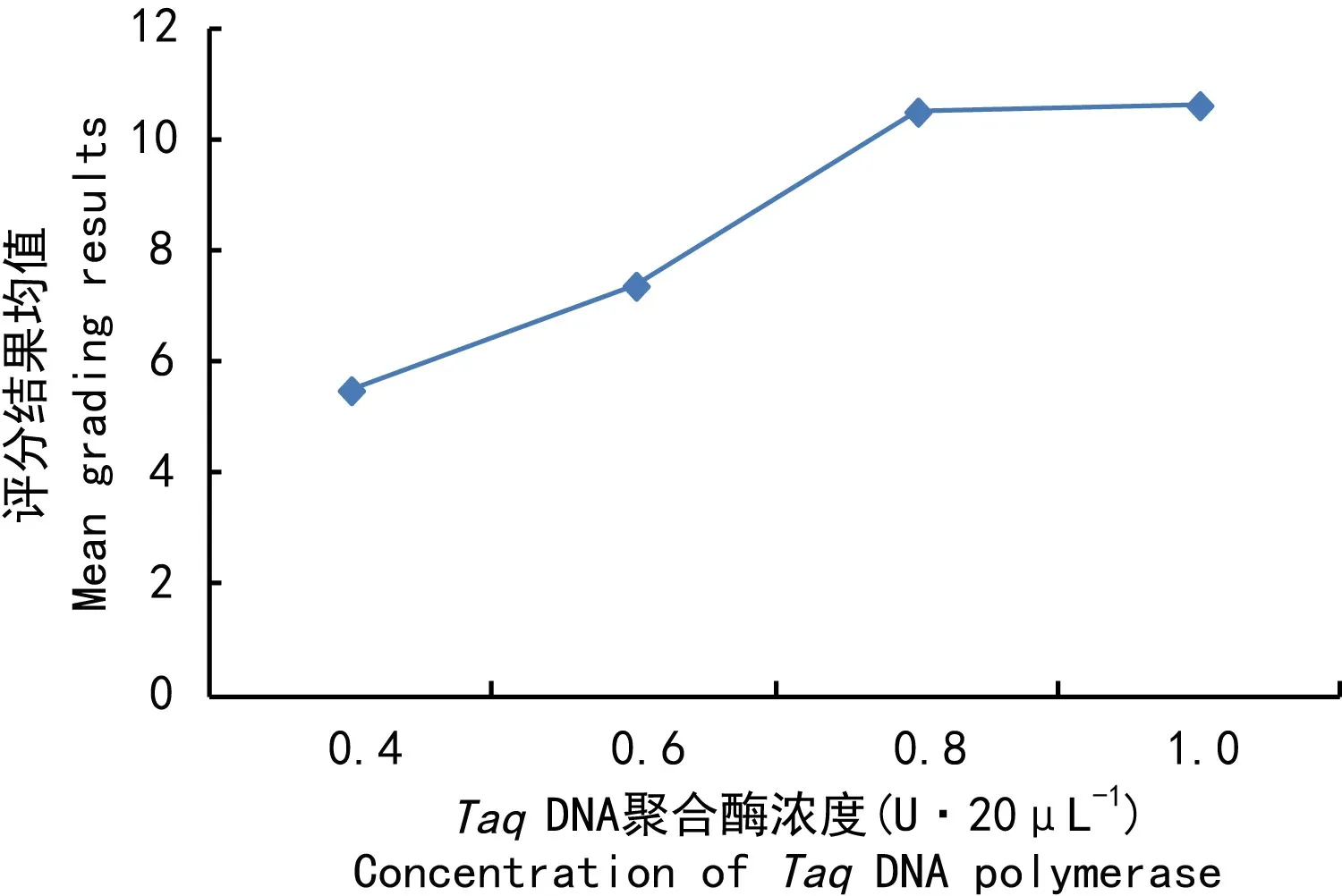
图 3 Taq DNA聚合酶浓度与评分结果均值的关系Fig. 3 Relationship between the concentration of Taq DNA polymerase and the mean grading results

图 4 引物浓度值与评分结果平均值的关系Fig. 4 Relationship between primer concentration and mean grading results
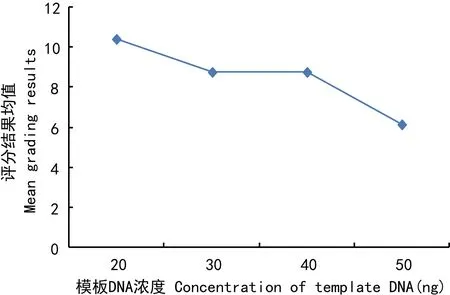
图 5 模板DNA浓度对评分结果平均值的影响Fig. 5 Influence of the concentration of template DNA on the mean grading results
评分结果均值随模板DNA用量的增加而减少(图5)。当DNA用量为20 ng时(处理1、8、10、15),因模板量太少,扩增的稳定性差。仅水平2与水平3差异不显著,水平1与其它3个水平差异显著,水平4与水平2、3差异也显著。因此,模板DNA的用量范围在30~40 ng均可。
dNTP对ISSR-PCR反应的影响最小(表3),评分均值结果波动不剧烈(图6)。当dNTP为0.075 mmol时(处理1、6、11、16),扩增条带少且不清晰;当dNTP大于0.100 mmol时,扩增条带逐渐增多。Duncan分析表明,水平1与水平3、水平2与水平4差异不显著;水平1、3与水平2、4差异显著。所以,选取dNTP的最佳浓度为0.100 mmol。
比较上述不同分析方法数据处理结果,本研究中每个因子(Mg2+、TaqDNA polymerase、 dNTP、primer和DNA template)的最适浓度组合是处理12,即20 μL反应体系中含有1.5 mmol·L-1Mg2+、1.0 U·20 μL-1TaqDNA聚合酶、0.100 mmol·L-1dNTP、0.3 μmol·L-1primer和40 ng 模板DNA。
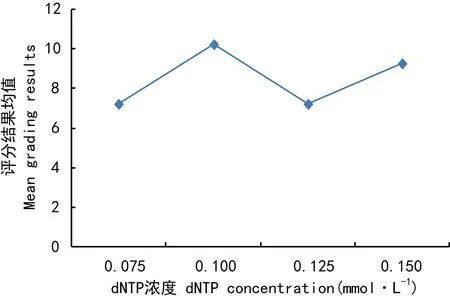
图 6 dNTP浓度对评分结果均值的影响Fig. 6 Influence of dNTP concentration on mean grading results
2.2 引物筛选及其最佳退火温度
图7为退火温度试验(UBC855引物)结果,退火温度对ISSR-PCR结果影响明显。当退火温度低于51.2 ℃时,小于800 bp的片段较少,1 200~1 400 bp的片段多,整体条带亮度较弱。当退火温度高于53.1 ℃时,400~800 bp的片段增多且逐渐清晰,大于1 600 bp的片段亮度增强,但1 200~1 400 bp的片段逐渐消失。退火温度为53.1 ℃时,所得扩增条带丰富且清晰稳定,因此引物UBC855的最佳退火温度为53.1 ℃。据此方法,筛选出12条理想的引物,其序列及最佳退火温度见表4。

图 7 引物(UBC855)退火温度试验 1-8泳道的复性温度分别为48.0、48.5、49.6、51.2、53.1、54.7、55.6、56.0 ℃; M. 200 bp标准分子量参照物。Fig. 7 Annealing temperature experiment of primer UBC855 Renaturation temperatures of lanes 1-8 were 48.0, 48.5, 49.6, 51.2, 53.1, 54.7, 55.6, 56.0 ℃ respectively; M. 200 bp molecular marker.
2.3 ISSR-PCR优化体系稳定性检测
用引物UBC855和UBC846对西藏嵩草不同个体进行扩增,验证优化体系的稳定性。从图8中可以看出,扩增产物电泳条带丰度高且清晰明亮。由此可见,优化后的ISSR反应体系是稳定可靠的,可用于西藏嵩草种质资源遗传结构分析。
3 讨论与结论
ISSR-PCR反应扩增结果受反应体系各组成成分、反应条件和实验材料等诸多因素的影响(胡延萍等, 2007; 李猛等, 2013)。对不同物种而言,其ISSR-PCR反应的最佳条件各异,并且由于试验设计的因素水平不同,各因素水平的影响也有差异。为确保ISSR分析结果的正确性及可重复性,须在实验前事先对体系条件进行优化筛选。本研究中,西藏嵩草ISSR-PCR扩增反应的影响因素为Mg2+>TaqDNA聚合酶>引物>模板DNA>dNTP。Mg2+对西藏嵩草ISSR-PCR反应的影响最为显著,这与蚬壳花椒(李猛等, 2013)和紫云英(孙清信等, 2012)中的研究结果一致。所以,确定合适的Mg2+浓度对西藏嵩草ISSR实验来说是非常重要的工作。反应液中处于游离态的Mg2+不仅会影响反应过程中TaqDNA聚合酶的活性,还可进一步结合反应液中的DNA template、dNTP和primer,从而会改变primer与template的结合效率、template与PCR产物的解链温度以及产物的特异性(胡延萍等, 2010)。因此,本研究适宜的Mg2+浓度为1.5 mmol·L-1。TaqDNA聚合酶对西藏嵩草ISSR-PCR的影响居第二位,仅次于Mg2+,TaqDNA聚合酶浓度过低会使PCR扩增片段少且条带弱,浓度过高不仅成本高而且会引起非特异性扩增。本研究以1.0 U为合适用量。引物是ISSR-PCR特异性反应的关键,引物浓度过低仅能扩增出大片段的条带,小片段的条带不能有效扩增;浓度过高则会引起错配和非特异性扩增,且会形成引物二聚体(胡延萍等,2014),本研究中适宜的引物浓度是0.3 μmol·L-1。模板DNA是ISSR-PCR的研究对象,在保证其质量的前提下,ISSR对模板DNA用量的要求范围通常较广,本试验中模板DNA的用量为30~40 ng。dNTP是对西藏嵩草ISSR-PCR反应影响最小的因素。本研究表明,dNTP浓度为0.100 mmol·L-1时,扩增的结果最理想。
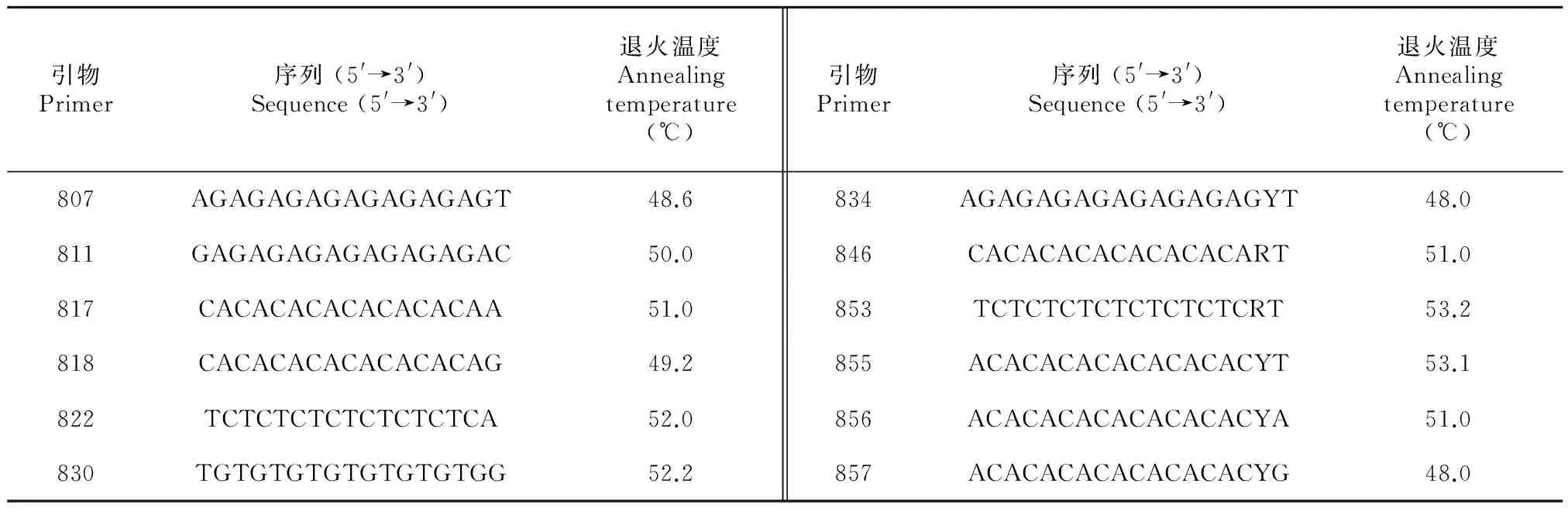
表 4 12条引物碱基组成及其最适复性温度
注: R = (A, G); Y = (C, T)。
Note: R=(A, G); Y = (C, T).
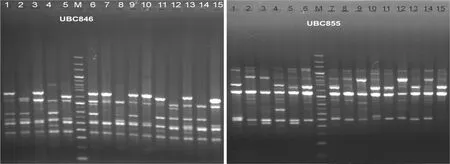
图 8 引物UBC846和UBC855对西藏嵩草不同个体ISSR-PCR扩增的结果 M. 100 bp+200 bp标准分子量参照物。Fig. 8 Electrophoresis results of different individuals in Kobresia tibetica by UBC846 and UBC855 M. 100 bp+200 bp molecular marker.
西藏嵩草ISSR-PCR不同引物的退火温度各异,其范围为48.0~53.2 ℃。不同的物种,不同的引物其碱基组成和序列长短不同,引物的退火温度也不同。包蕊等(2014)对华扁穗草ISSR分析时,引物UBC834的退火温度为55.4 ℃,在唐古特大黄中UBC834的退火温度为53 ℃(Hu et al, 2014),而本研究中,该引物的最适退火温度为48 ℃。因此,在ISSR-PCR反应中,对于不同的引物,须根据其熔点各自筛选其适宜的退火温度,不能统一而论。
正交试验设计可以对ISSR-PCR反应体系中的多个因素同时进行筛选,能够快速获得满意的水平组合,从而避免单因素试验繁琐且费时的缺点。本研究通过正交试验设计,优化西藏嵩草ISSR中每个因子的最适浓度,并采用直观分析和方差分析的方法,筛选出其最佳反应体系。即在20 μL反应体系中包括10 × PCR buffer 2 μL、1.5 mmol·L-1Mg2+、1.0 U·20 μL-1TaqDNA聚合酶、0.100 mmol·L-1dNTP、0.3 μmol·L-1primer和30~40 ng DNA模板。筛选出具有理想扩增结果的12条引物,并进一步确定了这12条引物对应的最适退火温度。利用筛选出的最佳反应条件对西藏嵩草不同个体进行验证,所得电泳图谱清晰稳定,能够迅速有效地检测出不同个体之间的差异。西藏嵩草是青藏高原高寒沼泽草甸的优势种,利用ISSR分子标记研究西藏嵩草种质资源遗传多样性和遗传结构,对青藏高原退化草甸的修复和保护具有重要的意义。
BAO R, HU YP, WANG L, et al, 2014. Optimization of ISSR-PCR system and primer screening forBlysmussinocompressus[J]. Chin J Grassl, 36(6): 46-52. [包蕊, 胡延萍, 王莉, 等, 2014. 华扁穗草ISSR-PCR反应体系优化及引物筛选 [J]. 中国草地学报, 36(6): 46-52.]
DELECTIS FLORA REPUBLICAE POPULARIS SINICAE AGENDAE ACADEMIAE SINICAE EDITA, 2000. Flora Deipublicae Popularis Sinicae [M]. Beijing: Science Press, 12: 33-34. [中国科学院中国植物志编辑委员会, 2000. 中国植物志 [M]. 北京: 科学出版社, 12: 33-34.]
DENG DS, LIU JQ, DENG ZF, et al, 1995. A preliminary study on the reproductive characters ofKobresiaon Qinghai-Xizang Plateau [J]. J Qinghai Norm Univ:Nat Sci Ed, 21(2): 24-29. [邓德山, 刘建全, 邓自发, 等, 1995. 青藏高原嵩草属植物繁殖系统生物学特性初探 [J]. 青海师范大学学报·自然科学版, 21(2): 24-29.]
DEVARUMATH RM, NANDY S, RANI V, et al, 2002. RAPD, ISSR and RFLP fingerprints as useful markers to evaluate genetic integrity of micropropagated plants of three diploid and triploid elite tea clones representingCamelliasinensis(China type) andC.assamicassp.assamica(Assam-India type) [J]. Plant Cell Rep, 21(2): 166-173.
DOYLE JJ, DOYLE JL, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue [J]. Phytochem Bull, 19: 11-15.
HU YP, WANG L, XIE XL, et al, 2014. Genetic variation in cultivated rhubarb (RheumtanguticumMaxim. ex Balf.) and the relationship with their wild relatives in China revealed by ISSR markers [J]. Plant Syst Evol, 300: 2 217-2 227.
HU YP, WANG L, BAO R, et al, 2014. Optimizing ISSR-PCR system ofCarexatrofuscausing orthogonal design [J]. Acta Agr Sin, 22(4): 911-914. [胡延萍, 王莉, 包蕊, 等, 2014. 正交设计优化黑褐苔草ISSR-PCR反应体系 [J]. 草地学报, 22(4): 911-914.]
HU YP, XIE XL, WANG L, et al, 2010. Establishment and optimization of ISSR-PCR reaction conditions forRheumtanguticum[J]. Guihaia, 30(1): 112-116. [胡延萍, 谢小龙, 王莉, 等, 2010. 唐古特大黄ISSR-PCR反应条件的优化 [J]. 广西植物, 30(1): 112-116.]
LI CX, SUN HS, 2009.Kobresiatibeticaswamping grassland resources and its protection in Qinghai [J]. Prat Sci, 26(9): 77-83. [李春秀, 孙海松, 2009. 青海省西藏嵩草沼泽化草甸资源及保护 [J]. 草业科学, 26(9): 77-83.]
LI HY, LI ZY, CAI LY, et al, 2013. Analysis of genetic diversity of Ruthenia Medic (MedicagoruthenicaL. Trautv.) in Inner Mongolia using ISSR and SSR markers [J]. Gen Resourc Crop Evol, 60: 1 687-1 694.
LI M, WANG P, SUN JK, et al, 2013. Establishment and optimization of ISSR-PCR reaction system onZanthoxylumdissitum[J]. Guihaia, 33(2): 185-190. [李猛, 王平, 孙吉康, 等, 2013. 蚬壳花椒ISSR-PCR反应体系的建立及优化 [J]. 广西植物, 33(2): 185-190.]
LI YY, DONG KH, 2014. ISSR analysis of genetic diversity of germplasm resources ofBothriochloaischaemumin Shanxi [J]. Acta Prat Sin, 23(1): 217-222. [李钰莹, 董宽虎, 2014. 山西白羊草种质资源遗传多样性的ISSR分析 [J]. 草业学报, 23(1): 217-222.]
MA YS, XU HF, 2013. Forage plants of three-river source region [M]. Beijing: Science Press. [马玉寿, 徐海峰, 2013. 三江源区饲用植物志 [M]. 北京: 科学出版社.]
NAGAOKA T, OGIHARA Y, 1997. Applicability of inter-simple sequence repeat polymorphisms in wheat for use as DNA markers in comparison to RFLP and RAPD markers [J]. Theor Appl Gen, 94: 597-602.
SUN QX, CHEN J, ZHANG H, et al, 2012. Optimization of ISSR’s primer and PCR reaction system forAstragalussinicusL. [J]. J Plant Gen Resour, 13(5): 870-878. [孙清信, 陈坚, 张辉, 等, 2012. 紫云英ISSR引物的筛选及PCR反应体系的优化 [J]. 植物遗传资源学报, 13(5): 870-878.]
TANG QY, FENG MG, 2006. DPS©data processing system-experimental design, statistical analysis and modeling [M]. Beijing: Science Press. [唐启义, 冯明光, 2006. DPS©数据处理系统-实验设计、统计分析及模型优化 [M]. 北京: 科学出版社.]
TIAN K, GUO HJ, YANG YM, 2009. The characteristic of ecological structure and research and practice of functional zoning in the conservation area of plateau wetland [M]. Beijing: Science Press. [田昆, 郭辉军, 杨宇明, 2009. 高原湿地保护区生态结构特征及功能分区研究与实践 [M]. 北京: 科学出版社.]
ZHANG SB, LU XF, 2012. Seed plants of Langcang River in Qinghai [M]. Xining: Qinghai Nationalities Publishing House: 362. [张胜邦, 卢学峰, 2012. 青海澜沧江源种子植物 [M]. 西宁: 青海民族出版社: 362.]
ZHOU XM, 2001. Kobresia meadow of China [M]. Beijing: Science Press. [周兴民, 2001. 中国嵩草草甸 [M]. 北京: 科学出版社.]ZIETKIEWICZ E, RAFALSKI A, LABUDA D, 1994. Genome fingerprinting by simple sequence repeats (SSR)-anchored polymerase chain reaction amplification [J]. Genomics, 20: 176-183.
Optimization of ISSR-PCR system onKobresiatibetica
HU Yan-Ping1, BAO Rui1, WANG Li1, SHI Lin1, 2, LI Yi1*
( 1.NorthwestInstituteofPlateauBiology,ChineseAcademyofSciences, Xining 810008,China; 2.UniversityofChineseAcademyofSciences, Beijing 100049, China )
Kobresiatibetica, belonging to the genus ofKobresiaand the family of Cyperaceae, is a perennial herb. The rhizomes ofK.tibeticaare short and culms are densely tufted. It is mainly distributed in Qinghai, Gansu, west of Sichuan provinces and east of Tibet Autonomous Region at altitudes ranging from 2 550 m to 4 950 m and can be found in alpine swampy meadows, weedy plains, marshes and riversides. The root system ofK.tibeticais developed. Besides, it is hygrophilous, resistant to cold and strong in vitality. For the reproduction ofK.tibetica, vegetative propagation is prior to sexual propagation.K.tibeticaplants are of highly ecological and economic importance. The stem and leaf ofK.tibeticaare luxuriant.K.tibeticacontains high levels of crude proteins and fats and is preferred by livestock over other plants found on the plateau. The production ofK.tibeticagrass is of high yield. As a consequence, it is the major grazing forage grass of summer and autumn in Qinghai-Tibet Plateau. Furthermore,K.tibeticais the constructive and dominant species of alpine swamp meadows in Qinghai-Tibet Plateau.K. meadows are also highly involved in carbon storage and serve as important carbon sinks in China. Therefore,K.tibeticawas chosen as the research object in this study. In order to establish the suitable Inter Simple Sequence Repeats Polymerase Chain Reaction (ISSR-PCR) system inK.tibetica, factors which affect the ISSR-PCR amplification, such as magnesium ion (Mg2+),TaqDNA polymerase, dNTP, primer and DNA template, were optimized and selected by orthogonal designed experiment of five factors in four levels. In addition, suitable ISSR primers and the annealing temperature of selected primers were screened from 100 ISSR primers by gradient PCR. As a result, the optimal reactions of ISSR-PCR inK.tibeticawere performed in a 20 μL volume containing 2 μL 10× PCR buffer, 1.5 mmol·L-1Mg2+, 1.0 UTaqDNA polymerase, 0.100 mmol·L-1dNTP, 0.3 μmol·L-1primer and 30-40 ng template DNA. Out of 100 ISSR primers screened, twelve primers were selected with clear and steady bands and suitable annealing temperature of each primer was 48.0-53.2 ℃, depending on primers used. The appropriate program of ISSR-PCR forK.tibeticawas as follows: an initial step of 5 min at 94 ℃ followed by 38 cycles of denaturing at 94 ℃ for 20 s, annealing at appropriate annealing temperature 48.0-53.2 ℃ (different primers with different annealing temperatures) for 1 min and extending at 72 ℃ for 80 s, ending with a final extension of 6 min at 72 ℃. The stability of the ISSR-PCR reaction system indicated that clear and rich polymorphism bands were obtained in different individuals ofK.tibeticawith the optimal system. The establishment of ISSR-PCR system was beneficial to the analysis of genetic diversity and genetic structure ofK.tibeticagermplasms. This study will provide the information for further study of screening high quality forage. Furthermore, it is of great significance for the study ofKobresia-swamp meadows in Qinghai-Tibet Plateau as well as the restoration and conservation of the wetland ecosystem.
Kobresiatibetica, Cyperaceae, ISSR-PCR, reaction sytem, orthogonal design
10.11931/guihaia.gxzw201411003胡延萍, 包蕊, 王莉, 等. 西藏嵩草ISSR-PCR反应体系优化研究 [J]. 广西植物, 2016, 36(8):915-922
HU YP, BAO R, WANG L, et al. Optimization of ISSR-PCR system onKobresiatibetica[J]. Guihaia, 2016, 36(8):915-922
2014-11-04
2015-05-07
国家自然科学基金(31200245);国家科技支撑计划项目(2012BAC08B04);中国科学院青年创新促进会项目(2014386);青海省国际科技合作项目(2015-HZ-808) [Supported by the National Natural Science Foundation of China(31200245); National Key Technology R & D Program(2012BAC08B04); Youth Innovation Promotion Committee of Chinese Academy of Sciences(2014386); International Cooperation Program for Science and Technology of Qinghai Province(2015-HZ-808)]。
胡延萍(1981-),女,山东定陶人,博士,副研究员,主要从事植物生物技术研究,(E-mail)yphu@nwipb.cas.cn。
李毅,研究员,博士生导师,主要从事植物遗传资源与繁育研究,(E-mail)liyi@nwipb.cas.cn。
Q945
A
1000-3142(2016)08-0915-08
