酸马奶发酵过程中理化指标与细菌相的动态变化
2016-09-10布仁其其格高雅罕包艳青任秀娟魏睿元萨如拉赵一萍
布仁其其格,高雅罕,包艳青,任秀娟,魏睿元,萨如拉,赵一萍,芒 来,*
(1.内蒙古农业大学动物科学学院,内蒙古自治区蒙古马遗传资源保护及马产业工程实验室,内蒙古呼和浩特 010018;2.内蒙古医科大学基础医学院,内蒙古呼和浩特 010110)
酸马奶发酵过程中理化指标与细菌相的动态变化
布仁其其格1,2,高雅罕1,+,包艳青1,任秀娟1,魏睿元1,萨如拉1,赵一萍1,芒来1,*
(1.内蒙古农业大学动物科学学院,内蒙古自治区蒙古马遗传资源保护及马产业工程实验室,内蒙古呼和浩特 010018;2.内蒙古医科大学基础医学院,内蒙古呼和浩特 010110)
分析了酸马奶发酵过程中(0~96 h)部分理化指标以及细菌群落的变化情况。结果显示酸马奶发酵96 h内pH为下降趋势,发酵初期迅速酸化,发酵后期下降速度缓慢趋于稳定,最低达到pH3.54;总蛋白质、脂肪含量存在一定的起伏变化,72 h达到最高值;乳糖含量减少,而乳酸含量增加;葡萄糖、半乳糖、乙酸、丙酸含量在发酵过程中处于较低的水平;细菌群落结构分析结果显示酸马奶发酵过程中存在丰富的细菌多样性,并且群落结构有明显的演替变化。乳杆菌属(Lactobacillus)为优势细菌属,在酸马奶发酵过程中12~24 h呈现出上升的趋势,而24~48 h时其含量下降。研究结果表明不同发酵时间酸马奶营养价值存有差异。
酸马奶,发酵过程,理化指标,细菌群落结构
马乳具有很高的营养价值,有研究表明可作为婴幼儿代乳品[1],还具有提高免疫力[2]、抗疲劳等功效[3]。马乳经微生物发酵成为酸马奶后其成分变得更加丰富易吸收,不仅营养保健价值得到了进一步提升,还延长了马乳保存期。
酸马奶主要依靠细菌和酵母菌发酵而成。殷文正[4]通过培养基法统计得知酸马奶发酵72 h时乳酸菌可达8×109CFU/mL,酵母菌可达1×108CFU/mL。乳酸菌数比酵母菌数高一个数量级,对品质形成具有重要作用。已有研究报道了新疆地区[5]、内蒙古锡林郭勒盟[6]以及蒙古国[7]等不同地区酸马奶中乳酸菌情况。酸马奶中乳酸菌研究手段从基于传统的分离培养鉴定[8-10]发展到以分子生物学方法分类鉴定[11-12],大大丰富了对其群落结构了解。然而以往对酸马奶中细菌及理化指标的关注只局限于发酵过程中某个时间点的样品检测,需要开展对酸马奶发酵过程中细菌以及理化指标的动态变化研究。
传统发酵酸马奶主要以牧户为单位酿制,还没有酿制的统一标准。为了使研究结果更加可靠,选取具有18年酿制酸马奶经验,酸马奶品质口感评价较高的牧户,完全参照牧户家做法酿制酸马奶采集样品。采用焦磷酸测序技术以及多种检测分析手段研究不同发酵时间点酸马奶中细菌群落结构和部分理化指标,从而对酸马奶传统发酵过程中细菌及成分变化规律进行初探。
1 材料及方法
1.1材料与仪器
在内蒙古锡林郭勒盟正镶黄旗一牧户家,参照牧户家做法酿制酸马奶进行样品收集。一个体系共3.5 L,其中鲜马奶与发酵剂以13∶1混匀,共有三个平行发酵,发酵剂采用牧民家中酸马奶。每隔2.5 h捣拌300次,一天共进行5次。发酵温度在(22±2)℃,每隔12 h采集一次样品收集至96 h,液氮保存带回实验室做后续实验。
乳酸、乙酸、丙酸、丁酸、乳糖、葡萄糖和半乳糖标准品美国Sigma;超纯水;甲醇:色谱纯、磷酸-磷酸盐缓冲液:0.010 mol/L pH=2.0自配、盐酸优级纯,天津市福晨化学试剂厂;E.Z.N.A Soil DNA提取试剂盒OMEGA公司。
450-GC气相色谱仪德国布鲁克公司;Agilent1100液相色谱仪美国Agilent公司、PB-10酸度计德国赛多利斯公司;Roche GS FLX基因组测序仪瑞士罗氏公司。
1.2实验方法
1.2.1pH测定采样现场以酸度计测定酸度值。
1.2.2总蛋白质采用凯氏定氮法测定[13](GB5009.5-2010)。
1.2.3脂肪采用Rose-Gonile(哥特里-罗兹法)法测定[14]。
1.2.4乳酸、乙酸、丙酸和丁酸测定称取室温融化后摇匀的酸马奶样品1.0 g,与3 mL的HCl(1 mol/L)震荡混匀,以12000 r/min的速度离心15 min,吸取上清液经0.45 μm滤膜过滤后进Agilent1100液相色谱系统(Zorbax SB-Aq,5 μm,4.6 mm×150 mm,Agilent,USA)分析。流动相为0.010 mol/L磷酸—磷酸二氢钠(pH=2.5),流速0.5 mL/min,紫外检测波长210 nm,柱温30 ℃,进样量为20 μL。
1.2.5乳糖、葡萄糖和半乳糖测定参考文献[15]的方法,称取室温融化后摇匀的酸马奶样品0.4 g,与1.2 mL超纯水混匀,超声波处理3 min后,以105 ℃高压处理15 min,再在4 ℃下以4500 r/min速度离心10 min。吸取上清液0.45 μm膜过滤,进Agilent1100液相色谱系统(CARBOSeq CHO-620 CA 2.5 μm,4.6 mm×250 mm,Agilent,USA)分析。流动相为水,流速0.6 mL/min,柱温90 ℃,进样量为5 μL。
1.2.6细菌鉴定方法采用E.Z.N.A Soil DNA提取试剂盒,按说明提取酸马奶宏基因组DNA。1%琼脂糖凝胶电泳检测抽提的基因组 DNA。PCR扩增16S rRNA 基因序列V1-V3区,使用Roche GS FLX 基因组测序仪测序,由上海美吉生物公司完成测序工作。使用QIIME(v1.4.0)分析平台开展序列的生物信息学分析[16-17],用RDP(Ribosomal Database Project)classifier[18]对序列进行同源性比对和种属分类学鉴定。
1.3数据统计分析
数据采用SPASS19.0软件进行统计分析,p值检验数据间差异显著性。
2 结果与分析
2.1酸马奶发酵过程中理化指标变化情况
2.1.1粗蛋白质与脂肪鲜马奶粗蛋白质平均含量2.09%,如图1所示,开始发酵24 h内无明显增长,24 h以后蛋白质含量逐渐增多,到72 h达到最高值2.92%,之后含量开始减少,96 h时含量为2.57%,但是各时间点蛋白质含量变化差异不显著(p>0.05)。蛋白质在发酵过程中被蛋白酶水解形成肽、氨基酸等小分子,部分作为能量物质继续代谢成为酸马奶的风味物质。同时酸马奶发酵过程中新增蛋白质或多肽类物质,例如具有抑菌杀菌作用的细菌素[19]等。数据表明在酸马奶发酵72 h内微生物的主要能源物质并非是蛋白质,分泌含氮物质增加大于消化利用速度。72 h之后蛋白质含量降低表明,在该发酵阶段,蛋白质作为氮源消耗加大。
鲜奶与发酵过程中不同时间点酸马奶的脂肪含量存在显著差异(p<0.05)。脂肪含量在发酵至48 h时无明显变化。随后缓慢增加,72 h时达到最高值1.38%,到96 h时酸马奶脂肪下降至1.33%,发酵过程中脂肪含量起伏变化差异不显著(p>0.05)。结果表明发酵过程中脂肪降解有限,而后期已形成的脂肪酸可能与乙醇发生酯化反应[20],使脂肪酸含量下降。在酸马奶发酵过程中所测得脂肪含量整体低于新疆地区酸马奶[21]脂肪含量,存在地区差异。
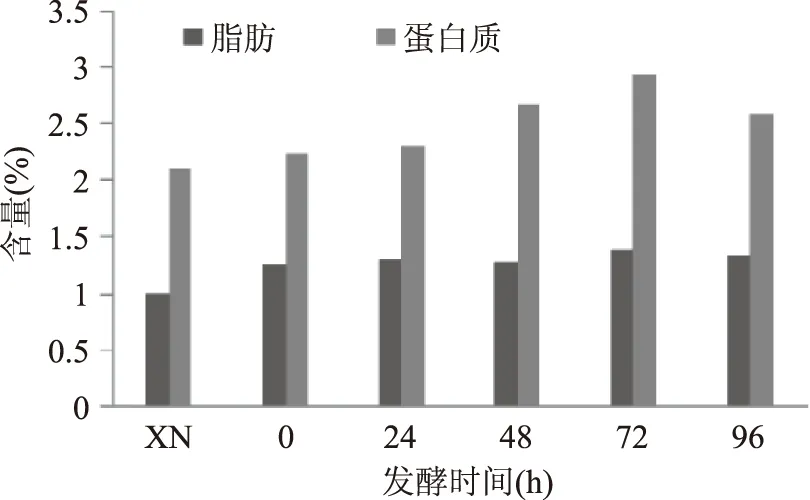
图1 酸马奶发酵过程中脂肪、蛋白质变化趋势Fig.1 The trend of fat and protein in the fermentation process of Koumiss 注:XN代表鲜奶,图2~图5同。
2.1.2乳糖、葡萄糖、半乳糖在鲜马奶中乳糖含量为20.83 g/L,随着发酵含量减少。从图2可看出前12 h几乎没有变化,12~48 h之间下降速度较快,48 h后下降速度相对缓慢,96 h时含量只有4.21 g/L。发酵48 h以后酸马奶乳糖含量变化差异显著(p<0.05)。从图3可看出鲜马奶中有少量的葡萄糖与半乳糖,加入发酵剂后含量增多,随后迅速被消耗,在后续的发酵过程中变化幅度不大,处于很低的水平,甚至60 h后未检测到葡萄糖。在发酵过程中产生的乳糖酶使乳糖分解成葡萄糖和半乳糖,进而转化为乳酸和其它有机酸。研究结果表明酸马奶发酵过程中乳糖迅速降解,48 h时分解50%,96 h时仅剩下20%。但是发酵过程中葡萄糖和半乳糖含量没有迅速增加,表明在发酵过程中葡萄糖、半乳糖利用速度大于生成速度。

图2 酸马奶发酵过程中乳糖变化趋势 Fig.2 The trend of Lactose in the fermentation process of Koumiss
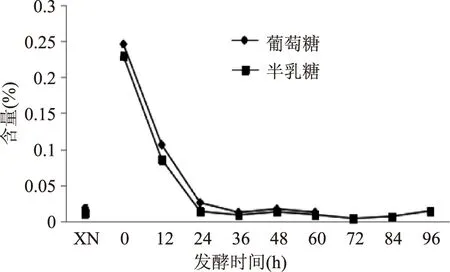
图3 酸马奶发酵过程中葡萄糖、半乳糖变化趋势 Fig.3 The trend of glucose,galactosen in the fermentation process of Koumiss
2.1.3乳酸、乙酸、丙酸、丁酸图4可知,酸马奶发酵36 h内乳酸含量增长迅速,接着出现一个短暂的负增长后又恢复增长并且趋于平稳。在96 h内乳酸含量自0.225%增至3.29%,变化差异显著(p<0.05),表明大量的乳糖被微生物酵解为乳酸。乳糖、乳酸在发酵过程中的变化趋势与殷文政[4]的研究结果一致。
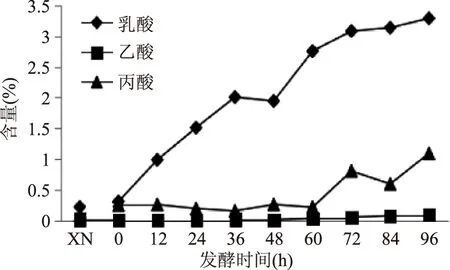
图4 酸马奶发酵过程中乳酸、乙酸、丙酸变化趋势Fig.4 The trend of lactic acid、acetic acidand propionic acid in the fermentation process of Koumiss
酸马奶发酵过程中有低量的乙酸生成,平均0.04%,60 h后变化差异显著(p<0.05)。由于乙酸是易挥发性有机酸,所得检测含量可能低于实际产出量。酒精发酵产生的乙醇与乳酸、乙酸等发生酯化反应,也会降低其含量。
丙酸在发酵前期无变化,在60 h后有明显的增加和下降的波动变化,差异显著(p<0.05),96 h时浓度为1.10%。丁酸在各个时期都未检测到。
2.1.4pH变化对三组平行样品的酸度检测,统计分析其变化趋势,如图5所示。鲜马奶的pH接近7,加上发酵剂后pH下降,但仍然高于6。在发酵36 h内下降相对较快,变化差异显著(p<0.05),之后下降速度缓慢趋于稳定,最低达到pH=3.54,表明酸马奶后期发酵环境酸度较高。
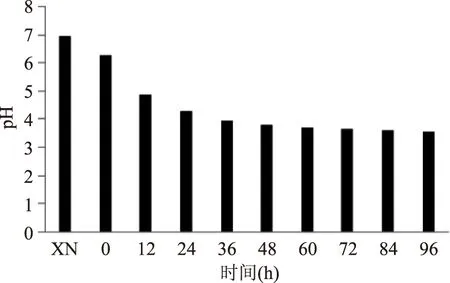
图5 酸马奶发酵过程中pH变化趋势Fig.5 The trend of pH in the fermentation process of Koumiss
2.2酸马奶发酵过程中细菌群落结构动态变化
2.2.1细菌16S rRNA基因序列分析对于酸马奶样品中细菌16S rRNA基因V1-V3区序列PCR扩增产物进行454焦磷酸高通量测序,18个样品共产生67514条高质量基因序列,每个样品平均产生3750条。经过PyNAST alignment 和100%序列鉴定聚类分析后,共得到11456条代表性序列。继而根据序列的97%相似性进行分类操作单元(Operational taxonomic units,OTU)划分及嵌合体检查,得到477个OTUs进行下一步分析。
2.2.2基于属水平的不同发酵时间点酸马奶细菌群落结构采用RDP数据库同源性序列比对与聚类相结合的方法对序列进行分类鉴定,共得到54个属,有0.25%的序列不能鉴定到属水平,表明酸马奶发酵过程中细菌具有丰富的多样性,还有发现新的菌种的可能性。
细菌群落结构分析结果显示在酸马奶传统制作过程中乳杆菌属(Lactobacillus,51.06%)为优势细菌属,乳球菌属(Lactococcus,30.29%)为次优势细菌属,其后是醋酸菌属(Acetobacter,7.13%)。殷文正[4]在其研究中也发现酸马奶发酵过程中乳杆菌属为优势细菌属。酸马奶发酵过程中乳杆菌属、乳球菌属与醋酸菌属动态变化情况如图6所示,在发酵起始12 h后乳杆菌属的繁殖速度加快,此时环境中酸度值能够满足其启动生长,24 h后其含量又呈现出下降的趋势,可能是菌群大量繁殖后出现了养分竞争而抑制了繁殖速度。与此相比,发酵起始后酸马奶中乳球菌属繁殖速度较快,至12 h后随着乳杆菌属数量增多其繁殖速度开始下降,当乳杆菌属自24 h开始数量下降时其繁殖速度又升高。72 h后乳杆菌属数量增加,而乳球菌属数量减少,在发酵过程中可能两者之间存在某种相互作用;酸马奶发酵起始所含醋酸菌属数量在发酵过程中减少,在48 h后其数量明显增多,72 h后又开始下降。乳球菌属与醋酸菌属在发酵24 h后发酵过程中的变化趋势相似。
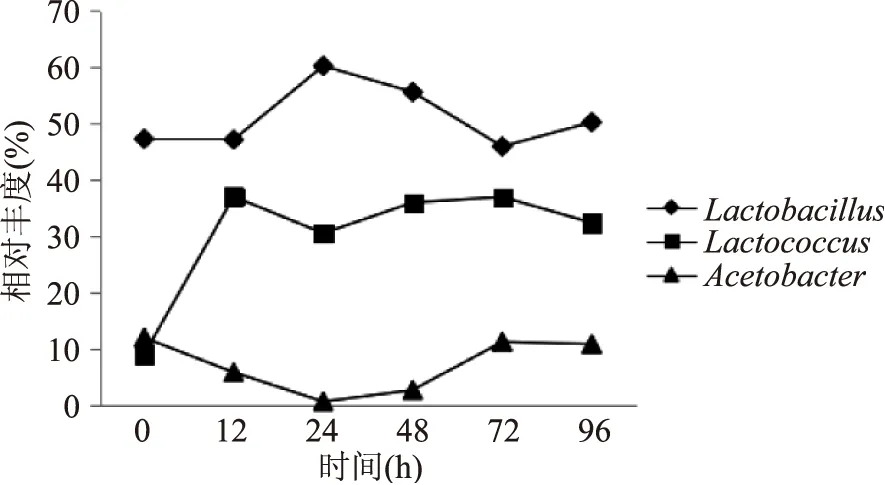
图6 基于属水平的不同发酵时间点酸马奶主要细菌属丰度变化Fig.6 The trend of main bacterial content based on genus levels at different time in the fermentation process of Koumiss
3 结论
酸马奶发酵过程中细菌相呈现多样性,其中优势细菌属为乳杆菌属,其次为乳球菌属。发酵过程中不同细菌属之间存在明显的演替变化以及相互作用。
发酵过程中乳糖含量减少,而乳酸含量增多,两者变化非常明显。葡萄糖、半乳糖、乙酸、丙酸含量较低,无明显动态变化。蛋白质、脂肪含量较高,存在一定的动态变化。
总而言之,研究结果表明不同发酵时间酸马奶营养价值存有差异。作为酸马奶发酵过程研究初探所得结果有限,还需开展氨基酸、脂肪酸、风味物质、乙醇、真菌微生物等在发酵过程中的状态研究,为高品质的酸马奶开发提供依据。
[1]刘亚东.马奶营养价值评定及在早产儿配方乳中的应用[D]. 哈尔滨:东北农业大学,2012.
[2]Miraglia N,Martin-Rosset W. Nutrition and feeding of the broodmare[M]. Wageningen,The Netherlands:Wageningen Academic Publishers,2006:133-147.
[3]刘洪元,安建钢,王海英,等. 鲜马奶缓解体力疲劳的研究[J]. 中国乳品工业,2010,38(2):24-27.
[4]殷文政,乌尼,钱建伟.锡林郭勒牧区马奶酒生物活性动态的研究[J]. 内蒙古农业大学学报,2002,23(1):9-16.
[5]孙天松,王俊国,张列兵.中国新疆地区酸马奶中乳酸菌生物多样性研究[J]. 微生物学通报,2007,34(3):451-455.
[6]乌日娜. 内蒙古传统酸马奶中乳杆菌的分离鉴定及16S rDNA 序列同源性分析[D]. 呼和浩特:内蒙古农业大学,2005.
[7]孟和毕力格,乌日娜,王立平. 不同地区酸马奶中乳杆菌的分离及其生物学特性的研究[J].中国乳品工业,2004,32(11):6-11.
[8]Ying A N,Adachi Y,Ogawa Y. Classification of lactic acid bacteria isolated from chigee and mare milk collected in Inner Mongolia[J]. Animal Science Journal,2004,75(3):245-252.
[9]熊素玉,姚新奎,谭小海,等. 酸马奶中乳酸菌的分离、纯化与鉴定[J]. 新疆农业科学,2007,44(5):696-701.
[10]霍小琰,李少英,郭荣荣. 酸马奶中乳酸菌的鉴定及生物学特性的研究[J]. 微生物学通报,2012,39(7):940-948.
[11]刘芳,都立辉,杜鹏,等.内蒙古酸马奶中乳酸菌多样性的研究(英文)[J]. 食品科学,2008,29(2):218-224.
[12]Hao Y,Zhao L,Zhang H.Identification of the bacterial biodiversity in koumiss by denaturing gradient gel electrophoresis and species-specific polymerase chain reaction[J]. Journal of Dairy Science,2010,93(5):1926-1933.
[13]GB 5009.5—85,食品中蛋白质的测定方法,中国标准出版社,1988.
[14]黄伟坤. 食品检验与分析[M]. 北京:中国轻工业出版社,1999,421-422.
[15]梁龙,牟光庆,陈历俊,等. 高效液相色谱法测定混菌发酵乳中乳糖,葡萄糖及半乳糖含量的变化[J]. 食品工业科技,2013,34(13):292-295.
[16]Lee OO,Wang Y,Yang J,et al. Pyrosequencing reveals highly diverse and species specific microbial communities in sponges from the Red Sea[J]. ISME J,2011,5(4):650-664.
[17]Kuczynski J,Stombaugh J,Walters W A,et al. Using QIIME to Analyze 16S rRNA Gene Sequences from Microbial Communities[J]. Curr Protoc Microbiol,2012,Chapter 1,Unit1E 5.
[18]Cole J R,Chai B,Farris R J,et al. The ribosomal database project(RDP-II):introd-ucing my RDP space and quality controlled public data[J]. Nucleic Acids Res,2007,35(Database issue):169-172.
[19]孙健. 酸马奶酒中抗菌物质的分离及抗菌作用的研究[D]. 呼和浩特:内蒙古农业大学,2003.
[20]崔玉琦,王雅炜,武瑞霞,等. 酵母菌在发酵乳制品中的应用研究[J]. 食品科技,2014(10):53-56.
[21]孙天松. 新疆维吾尔自治区酸马奶化学组成分与微生物学分析[J]. 中国乳品工业,2005,33(10):9-13.
Analysis of physicochemical parameters and bacteria community structure in the Koumiss’s fermentation process
Burenqiqige1,2,GAO Ya-han1,+,BAO Yan-qing1,REN Xiu-juan1,WEI Rui-yuan1,Sarula1,ZHAO Yi-ping1,Dugarjaviin Manglai1,*
(1.College of Animal Science,Inner Mongolia Agricultural University;Inner Mongolia Mongolian horse genetic resources protection and industrial engineering laboratory,Hohhot 010018,China;2.Department of Basic Medical,Inner Mongolia Medical University,Hohhot 010110,China)
To analyze the physicochemical parameters and bacteria community structure in the Koumiss’s fermentation process(0~96 h). The main findings were introduced in following part. During the fermentation process,the pH value of Koumiss was declined at high speed in early period and then the decent speed was slowed down and finally reached to the minimum value(pH=3.54). The total contents of protein and fats were fluctuant changed,and achieved to the maximum values at 72 h. The content of lactose was decreased,while the lactic acid was increased in the process of fermentation. The contents of glucose,galactose,acetic acid,and propionic acid were at low level during fermentation. The results of bacterial community structure analysis showed that there were abundant bacterial diversity and their dynamic community structures changed obviously in the Koumiss fermentation process.Lactobacilluswas the dominant bacterial genus. Its content showed a rising trend during 12 h to 24 h,while decling at 24 h to 48 h. The result showed that there were differences in nutritional value of Koumiss at different fermentation period.
Koumiss;fermentation process;physicochemical parameters;bacterial community structure
2016-01-29+为并列第一作者
布仁其其格(1982-),女,博士研究生,讲师,研究方向:分子数量遗传学,E-mail:qiqigejin@163.com。
高雅罕(1986-),女,博士研究生,研究方向:分子数量遗传学,E-mail:gaoyahan_999@126.com。
芒来(1962-),男,博士,教授,研究方向为:分子数量遗传学与马的育种,E-mail:dmanglai@163.com。
国家科学技术部对俄科技合作专项(2011DFR30860);内蒙古自治区科技厅重点实验室建设项目(20130902);内蒙古自治区科技厅应用技术研究与开发项目(20140172);呼和浩特市科技计划项目(2014-农-16)。
TS252.56
A
1002-0306(2016)11-0118-04
10.13386/j.issn1002-0306.2016.11.016
