Mn、Zn和Fe对醉马草内生真菌共生体麦角酰胺和麦角新碱含量的影响
2016-09-02万志文冯疆蓉李春杰
万志文,冯疆蓉,王 萍,李春杰
(兰州大学 草地农业科技学院,草地农业生态系统国家重点实验室,兰州 730020)
Mn、Zn和Fe对醉马草内生真菌共生体麦角酰胺和麦角新碱含量的影响
万志文,冯疆蓉,王萍,李春杰*
(兰州大学 草地农业科技学院,草地农业生态系统国家重点实验室,兰州 730020)
该研究通过温室砂培试验,考察了不同浓度Mn、Zn和Fe处理对醉马草内生真菌共生体麦角酰胺和麦角新碱含量的影响,以明确共生体产碱所需3种微量元素的最佳浓度范围。结果表明:(1) 在短时间(3周)处理下,共生体麦角酰胺和麦角新碱含量均在Mn2+浓度为5.0 mmol·L-1时达到最大值,且显著高于其余Mn2+浓度梯度;在较长时间(6、9 周)处理下,其麦角酰胺含量在Mn2+浓度为5.0 mmol·L-1时最大但增幅不明显,麦角新碱在对照(0.01 mmol·L-1Mn2+)处理下积累量达到最大,且显著高于其余处理。(2)共生体麦角酰胺含量在处理时间为3、6和9周时, Zn2+处理均在7.0 mmol·L-1下达到最大值,且处理3 周的含量显著高于其余处理时间;其麦角新碱在3个处理时间下均于Zn2+浓度为21.0 mmol·L-1时达到最大值。(3) 在不同处理时间下,共生体麦角酰胺的含量均在Fe浓度为6.0~12.0 mmol·L-1下达到最大值,且在3 周时的含量显著高于其余处理时间;麦角新碱含量在第3、6 周时均于Fe浓度为0 mmol·L-1下达到最大值,在第9 周时则于Fe浓度为0.03 mmol·L-1条件下达到最大值。研究认为,高浓度短时间的Mn、Zn和Fe元素处理有利于刺激醉马草内生真菌共生体2种麦角碱的积累,可通过控制3种微量元素的浓度和共生体的生长时间来提高2种麦角碱的含量,为今后能萃取更高浓度的麦角酰胺和麦角新碱作为临床药剂或生物农药提供较精确的外界条件。
醉马草;内生真菌;麦角酰胺;麦角新碱;Mn;Zn;Fe
醉马草(Achnatheruminebrians)是中国北方天然草地的烈性毒草之一,系禾本科芨芨草属(Achnatherum)多年生草本植物,在新疆、青海、内蒙古、西藏、甘肃等省(区)分布较广[1]。禾草内生真菌(fungal endophyte)是一类能够在禾草中完成全部或大部分生命周期,但不会对宿主植物致病的一类真菌[2]。一般禾草中内生真菌主要是Epichlo⊇ 属及其无性阶段的Neotyphodium属,根据最新国际真菌命名法规,现统一称为Epichlo⊇内生真菌[3]。自1994年美国学者Bruehl等[4]首次在采自中国新疆的醉马草种子中发现了内生真菌,到2000年南志标和李春杰调查发现,中国甘肃省醉马草植株的内生真菌带菌率近乎100%[5]。据报道,内生真菌提高了醉马草的抗旱性、耐盐性[6]、抗虫性[7]、抗病性[8]和对重金属胁迫的耐受性[9-11],而醉马草提供了内生真菌生长发育所需的营养物质,两者形成互惠的共生体。醉马草内生真菌共生体可产生2种重要的麦角生物碱,即麦角酰胺和麦角新碱,这2种生物碱作为一类控制伤口血流量的药物已被广泛应用于临床[12-13]。但其也是醉马草引起家畜和食草昆虫中毒的主要原因[14-16]。因此,麦角新碱和麦角酰胺既具有一定的药用价值,也可作为环保绿色的生物防治农药进一步开发利用。
植物生长发育除需要大量元素碳、氢、氧、氮、磷和钾等以外,还需要微量元素硼、钼、铜、锌和铁等。微量元素在植物体内虽然含量极少,但它是植物正常生长不可或缺的物质,对植物体内生理生化过程起着至关重要的作用。缺少微量元素后植物会出现一系列的生理病症,比如植物缺B会出现花而不实,这是因为在缺B条件下,花粉的萌发和花粉管的生长受到了抑制,无法形成正常的果实。Mn、Zn和Fe在植物体内均参与多种酶的组成、光合作用和呼吸作用等,缺乏这3种微量元素会导致植物体内代谢受阻,叶片失绿,产量下降等。但过量的Mn、Zn和Fe会对植物产生毒害,影响植物正常生长[17-21]。
微量元素除了参与上述各种生理生化反应,还在植物体内次生产物的代谢过程中具有重要的作用。如硼和铜均能与高等植物体内类黄酮反应生成络合物,使类黄酮在植物体内的存在更加稳定[22]。生物碱是常见的次生代谢物,其含量与生物因子(如基因型、病虫害、食草动物、 微生物、人工干扰等)和非生物因子(如光照、温度、土 壤、水分等)有关[23-25]。据报道,禾草内生真菌中的生物碱与大量元素有关,如 内生真菌麦角类生物碱的合成与P元素有关[26];土壤中有效P能够影响带菌高羊茅中麦角生物碱的含量[27];N和Ca也与新西兰草甸羊茅中生物碱含量有关[28];N、P、NP组合及色氨酸对醉马草麦角酰胺和麦角新碱含量也有一定的促进作用[29]。但关于微量元素对麦角生物碱合成代谢影响的研究目前尚无报道。王萍等[30]对Mn、Zn和Fe胁迫下醉马草的生长和生理指标进行了探讨,但未对麦角新碱和麦角酰胺进行进一步的检测,因此本试验拟在不同浓度的Mn、Zn和Fe处理下考察醉马草内生真菌共生体的产碱情况,通过高效液相色谱检测其麦角新碱和麦角酰胺的含量变化,以明确醉马草内生真菌共生体生物碱积累所需3种微量元素的最佳浓度范围,为提取更高浓度的麦角酰胺和麦角新碱探索最佳条件。
1 材料和方法
1.1试验材料
醉马草种子于2011年采自甘肃天祝打柴沟(102°52′ E,37°12′ N)并播种于兰州大学榆中校区,2012年10月收获的种子5℃保存于农业部牧草与草坪草种子质量监督检验中心(兰州)种子储藏室,2013年3月建立带菌(E+)和不带菌(E-)醉马草小区,2014年7月将收获的醉马草种子用于本试验。3种微量金属元素分别以MnCl24·H2O、ZnSO4·7H2O和Fe·citrate·3H2O分析纯试剂的形式添加。
1.2材料培养与处理
挑选籽粒饱满、表面健康的带菌醉马草种子于2装有300 g混合培养基质的(蛭石∶珍珠岩=3∶1)聚乙烯花盆(口径15 cm,底径10 cm,深12 cm)中,在兰州大学榆中校区智能温室条件下[光周期12 h光照,温度(20±1) ℃,光照强度120 μmol·m-2·s-1]进行培养。每周定量浇灌300 mL Hoagland营养液。待醉马草幼苗生长到第4周龄后开始处理,处理液元素种类和处理水平见表1。Mn和Zn元素浓度梯度的设置参考苏加义等[31]的方法,Fe元素浓度梯度的设置参考高大文等[32]的方法。以浇灌完全营养液作为对照,其他各浓度梯度作为处理,每个浓度梯度(共计5个浓度梯度)3次重复。每周浇灌蒸馏水和处理液各1次,每次浇灌量为300 mL。试验各处理梯度采取完全随机区组摆放。在处理第3周、6周和9周后对醉马草内生真菌共生体麦角酰胺和麦角新碱含量进行测定。
1.3醉马草内生真菌共生体幼苗麦角生物碱的提取及其含量的测定
将待测醉马草幼苗在-20 ℃的冰箱冰冻5 h,并在冷冻干燥机中-60 ℃干燥24 h后,取出速于研钵中研磨成粉末。称取50 mg样品装于1.5 mL的eppendorf离心管中;加入1 mL提取液(CHCl3∶MeOH∶NH4OH=75∶25∶2), 颠倒混匀,室温黑暗条件下过夜。 后经15 ℃、10 000 r·min-1条件下离心5 min,于通风厨中黑暗条件下自然挥发干燥。每个离心管加入0.75 mL MeOH∶CCl4(1∶2)和0.25 mL 25 mmol·L-1酒石酸,颠倒混匀后,于离心机(Beckman, Germany)中在21 ℃、12 000 r·min-1条件下离心5 min,吸取上清液0.25 mL经0.22 μm孔径的有机相过滤塾过滤至1.5 mL棕色色谱瓶,待用[33]。用Agilent 1 100 series高效液相色谱系统,ZORBAX-XDB C18色谱柱,流动相流速1 mL·min-1进行检测。流动相A为0.1 mol·L-1NH4OAc,B为乙腈/0.1 mol·L-1NH4OAc(3/1)。按照95% A液3 min、70% A液22 min、65% A液5 min、95% A液10 min流动相配比与时间,检测波长312 nm,发射波长427 nm[34]。
1.4数据处理
所有数据均用Excel录入,并作图。采用SPSS 16.0统计软件进行差异显著性分析,用Duncan法进行多重比较。
2 结果与分析
2.1Mn处理对醉马草内生真菌共生体麦角生物碱含量的影响
2.1.1麦角酰胺含量随着处理时间的延长,醉马草内生真菌共生体麦角酰胺含量在不同浓度的Mn2+处理条件下均呈先降低后增加的趋势,并均在处理第3 周时达到最大值,且大多显著(P<0.05)高于其余处理时间(表2)。在相同处理时间内,共生体麦角酰胺含量随着Mn2+处理浓度的升高先增加后降低,且各处理时间均在Mn2+处理浓度为5.0 mmol·L-1时达到最大值,分别比对照(CK)增加了43.8%、26.5%和8.1%,而其它各个处理均与对照无显著性差异(P>0.05)。
2.1.2麦角新碱含量由表2可知,随着处理时间的增加,醉马草内生真菌共生体麦角新碱含量在对照和0.0 mmol·L-1Mn2+处理下呈逐渐增加的趋势,在处理第9 周时达到最大值,此时分别是处理第3 周时的10.06倍和2.76倍(P<0.05);在2.5 mmol·L-1Mn2+处理下,共生体麦角新碱含量随着时间的延长呈缓慢下降趋势,但没有显著变化(P>0.05);在5.0和7.5 mmol·L-1Mn2+处理下,共生体麦角新碱含量随着处理时间的增加呈先降低后上升的趋势,但仍以处理3周时显著较高。在处理第3周时,共生体麦角新碱含量以5.0 mmol·L-1Mn2+处理最高,且显著高于其余Mn2+浓度梯度(P<0.05),是对照的4.53倍;而在处理第6、9周时,共生体麦角新碱含量均在对照下达到最大值,且显著高于其余处理(P<0.05)。
以上结果说明,较高浓度的Mn2+对醉马草内生真菌共生体麦角酰胺的积累具有一定的促进作用,且在短时间内效果更好;同样,短时间的高浓度Mn2+处理可以刺激共生体产生麦角新碱,而长时间的高浓度处理则不利于麦角新碱的积累。

表1 盆栽试验处理元素种类和处理水平[28]

表2 不同Mn处理下醉马草幼苗麦角酰胺和麦角新碱含量的变化
注:表中数据为平均值±标准误(n=5);同列中的不同小写字母表示同期处理间差异显著(P<0.05),而同行不同大写字母表示处理时间之间差异显著(P<0.05)。下同
Note: Data are present as mean ± SE(n=5); Different normal letters within the same column mean significant difference among treatments at 0.05 level, while different capital letters within the same row mean significant difference among stages at 0.05 level. The same as below.
2.2Zn2+处理对醉马草幼苗麦角生物碱含量的影响
2.2.1麦角酰胺含量表3显示,随着Zn2+处理时间的延长,醉马草内生真菌共生体麦角酰胺含量在0 ~14.0 mmol·L-1Zn2+处理下呈先降低后增加的趋势,并在处理第3 周时达到最大值,且显著(P<0.05)高于其余处理时间;而在21.0 mmol·L-1Zn2+浓度时,共生体麦角酰胺含量呈逐渐下降的趋势,且处理第3 周时的含量显著(P<0.05)高于其余处理时间。在相同处理时间内,醉马草内生真菌共生体麦角酰胺含量随着Zn2+处理浓度的升高大多呈先增加后降低的趋势;在处理第3、6和9周时,共生体麦角酰胺含量均在7.0 mmol·L-1Zn2+处理下达到最大值,分别比对照显著增加了84.0%、33.1%和44.5%;在处理第9 周时,21.0 mmol·L-1Zn2+浓度处理的麦角酰胺含量显著(P<0.05)低于其余的浓度处理,与对照相比显著减少了28.5%。
2.2.2麦角新碱含量从表3还可看出,随着Zn2+处理时间的延长,醉马草内生真菌共生体麦角新碱含量在不同浓度的Zn2+处理下均逐渐增加,并在处理第9 周时达到最大值,且大多显著(P<0.05)高于其余处理时间。醉马草内生真菌共生体麦角新碱含量在处理第3周时随着Zn2+处理浓度的升高而增加,而在处理第6、9 周时则随着Zn2+处理浓度增加呈先降低后升高的趋势;在3个处理时间下,共生体麦角新碱含量均在21.0 mmol·L-1Zn2+处理下达到最大值,分别比对照增加了3.66倍、1.55倍和1.29倍。
以上结果说明,短时间内醉马草内生真菌共生体产生麦角酰胺的最适宜Zn2+浓度为7 mmol·L-1,过高或过低浓度均会对其产生不利影响,而长时间内Zn2+处理对麦角酰胺的积累效果不佳;但是,长时间高浓度的Zn2+处理有利于共生体麦角新碱的积累。
2.3Fe处理对醉马草幼苗麦角生物碱含量的影响
2.3.1麦角酰胺含量随着Fe处理时间的延长,醉马草内生真菌共生体麦角酰胺含量在不同浓度的Fe处理下呈先降低后增加的趋势,并均在处理第3 周时达到最大值,并大多显著高于其余处理时间(表4)。在相同处理时间内,共生体麦角酰胺含量随着Fe处理浓度的升高呈先增加后降低的趋势;在处理第3、6 周时,共生体麦角酰胺的含量均在Fe浓度为6.0~12.0 mmol·L-1下达到最大值,分别比对照显著增加了大约109.3%和204.2%;在处理第9 周时,共生体麦角酰胺含量在Fe浓度为6.0 mmol·L-1处理下达到最大值,与同期对照相比显著增加了113.9%。

表3 不同Zn处理下醉马草幼苗麦角酰胺和麦角新碱含量的变化
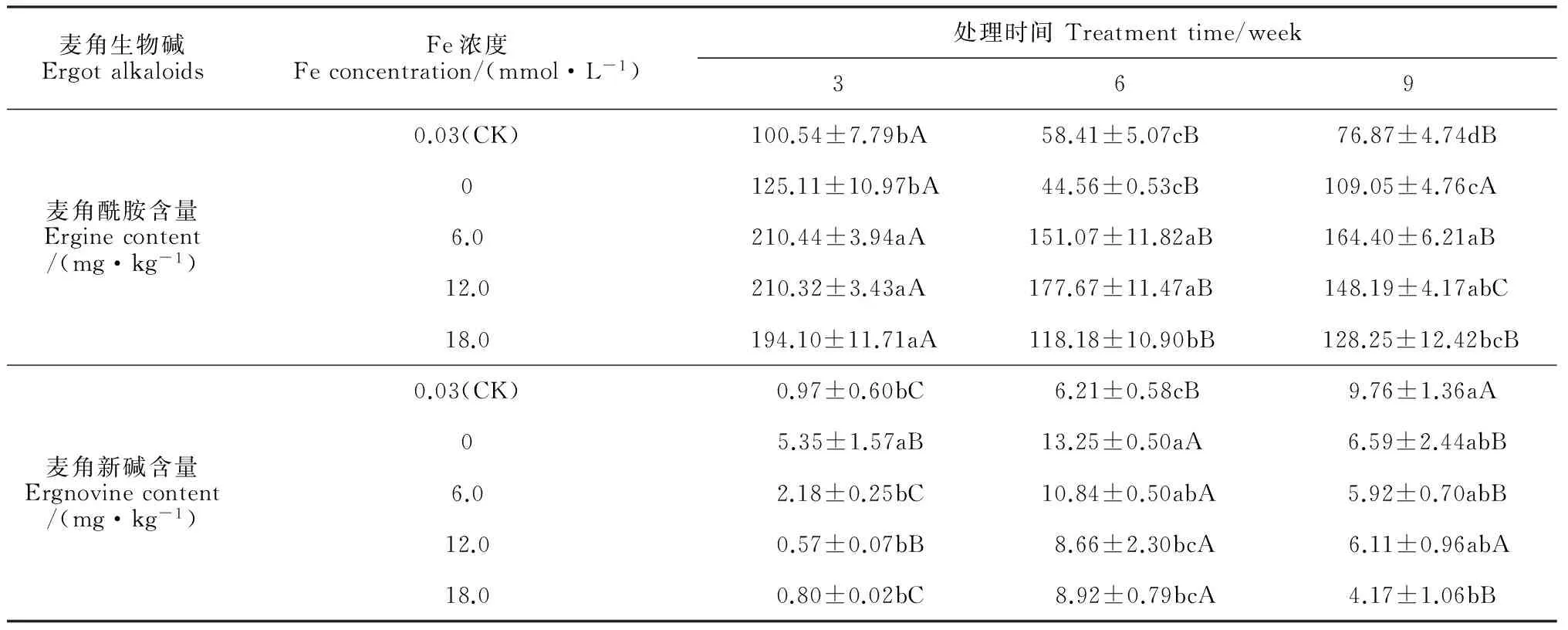
表4 不同Fe处理下醉马草幼苗麦角酰胺和麦角新碱含量的变化
2.3.2麦角新碱含量表4显示,随着处理时间的延长,醉马草内生真菌共生体麦角新碱含量在对照组表现为逐渐增加,而在其他Fe浓度处理下均表现为先增加后减小的趋势,各浓度处理第3周的含量均明显低于第6、9周。在处理第3、6 周时,共生体麦角新碱含量均在Fe浓度为0 mmol·L-1处理下达到最大值,与对照相比分别显著增加了5.52和2.13倍;而在处理第9 周时,麦角新碱含量在Fe浓度为0.03 mmol·L-1条件(CK)下达到最大值。
从以上分析结果可知,短时间高浓度的Fe对醉马草内生真菌共生体麦角酰胺的产生具有较强促进作用;而共生体麦角新碱产生对于Fe的需求量较少,但长时间的生长需要补充一定量Fe元素。
3 讨 论
麦角碱生物合成首先是合成麦角灵环。由乙酰辅酶A生成的甲戊二羟酸(MVA)是生物碱合成的起始物,进而形成焦磷酸二甲烯丙酯,在二甲烯丙基色氨酸合酶催化下与L-色氨酸发生环合得到二甲烯丙基色氨酸(DMAT)。DMAT在甲基转移酶催化下发生N-甲基化,之后脱羧并环合形成裸麦角碱,裸麦角碱为麦角灵环生物合成的起始物质[27,35]。而二甲烯丙基色氨酸合酶(dimethylallytrytophan synthase,DMATS) 已被证明是整个麦角生物碱合成过程中的限速酶[36],色氨酸既是共生体中麦角碱生物合成的前体也是该关键酶的诱导物。
早在1948年Tsui[37]发现缺Zn的番茄植株中色氨酸含量显著下降,1951年Nason等[38]观察到链孢霉在缺Zn条件下色氨酸含量减少,并且认为是由色氨酸合成酶活性下降所致。后来学者对缺Zn玉米幼苗进行代谢物测定时也发现色氨酸含量下降的现象[39-40],Zn被证实是色氨酸合成所必需的元素。因此,Zn可以通过影响色氨酸的合成间接地影响麦角生物碱的合成。除此之外,Zn还可以通过参与醉马草内生真菌共生体的生理生化过程影响其生长,进而影响产碱。王萍等[30]试验表明,当Mn、Zn和Fe处理浓度低于或高于对照时,醉马草内生真菌共生体株高、分蘖和生物量均呈下降趋势;而随着3种微量元素浓度的升高,脯氨酸(Pro)、丙二醛(MDA) 过氧化物酶(POD)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)含量呈上升趋势。本试验结果发现,不同浓度的Zn和不同处理时间对醉马草内生真菌共生体2种麦角碱的影响也不同,短时间处理下最适宜麦角酰胺积累的Zn浓度为7 mmol·L-1,长时间Zn处理的效果不如短时间处理明显,而长时间高浓度的Zn处理则有利于共生体麦角新碱的积累。
Mn直接参与植物光合作用中电子传递系统的氧化还原过程及PSⅡ系统中水的光解,并对维持叶绿体膜正常的结构有重要的作用[41]。Mn含量的多少直接影响植物的生长状况,过量的Mn会导致叶片输导组织坏死,蛋白质合成受阻,而Mn不足则会通过作用于光合作用等生理过程减缓生命进程[42]。本试验研究表明,在整个Mn处理过程中,醉马草内生真菌共生体麦角酰胺的含量均在Mn浓度为5.0 mmol·L-1下达到最大值,且在短时间内增幅最大,同样共生体麦角新碱含量在处理第3 周、Mn浓度为5.0 mmol·L-1下达到最大值,但超过3 周后,其含量在较低浓度的对照(0.01 mmol·L-1)条件下显著高于其他Mn处理浓度,说明短时间内的较高浓度Mn对麦角新碱产生的刺激效果最佳,这可能是由于长时间的高浓度Mn处理对醉马草内生真菌的生长有一定的抑制作用,进而影响其产碱。
Fe除了参与植物体内光合作用、氧化还原反应和电子传递之外,还是与氮代谢有关的固氮酶、硝酸还原酶和亚硝酸还原酶等的组成成分[43-44]。麦角生物碱是一类含氮化合物,因此,Fe既可以通过直接影响醉马草内生真菌共生体的生长来影响其产碱,也可能直接参与麦角碱的合成。在本试验中,当Fe的处理浓度为6.0~12.0 mmol·L-1时,最适宜醉马草内生真菌共生体麦角酰胺的积累,而麦角新碱在短时间内对Fe的需求量较少,但随着生长时间的延长,需要适当补充一定量较低浓度(0.03 mmol·L-1)的Fe元素。
综上所述,本研究首次在不同浓度Mn、Zn和Fe的处理下探讨了醉马草内生真菌共生体麦角酰胺和麦角新碱含量的变化规律,总体来说,高浓度短时间的 3 种微量元素处理有利于刺激2种麦角碱的积累,但其刺激醉马草内生真菌共生体的产碱机理还有待进一步研究。
[1]史志诚. 中国草地重要有毒植物[M]. 北京: 中国农业出版社, 1997: 166-176.
[2]SIEGEL M R, LATCHG C M, JOHNSO M C. Fungal endophytes of grasses[J].AnnualReviewofPhytopathology, 1987, 25: 293-315.
[3]LEUCHTMANN A, BACON C W, SCHARDL C L,etal. Nomenclatural realignment ofNeotyphodiumspecies with genusEpichlo⊇[J].Mycologia, 2014, 106(2): 202-215.
[4]BRUEHL G W, KAISER W J, KLEIN R E. An endophyte ofAchnatheruminebrians, an intoxicating grass of northwest China[J].Mycologia,1994,86(6):773-776.
[5]NAN Z B, LI C J.Neotyphodiumin native grasses in China and observations on endophyte/host interaction[C]// PAUL V H, DAPPRICH P D. Proceedings of the 4th InternationalNeotyphodium/Grass Interactions Symposium. Soest, Germany, 2000, P41-50.
[6]ZHANG X X, LI C J, NAN Z B. Effects of salt and drought stress on alkaloid production in endophyte-infected drunken horse grass (Achnatheruminebrians)[J].BiochemicalSystematicsandEcology,2011, 39(4-6):471-476.
[7]张兴旭, 陈娜, 李春杰, 等. 禾谷缢管蚜与内生真菌互作对醉马草幼苗的生理影响[J]. 草地学报, 2008, 16(3): 244-261.
ZHANG X X, CHEN N, LI C J,etal. Physiological effects ofRhopalosiphumpadionAchnatheruminebriansKeng infected by endophyte[J].ActaAgrestiaSinica, 2008, 16(3): 244-261.
[8]柳莉,郭长辉,吕卉,等. 内生真菌对醉马草白粉病抗性的影响[J]. 草业学报, 2015, 24(11): 66-71.
LIU L, GUO C H, LÜ H,etal. Effects of theEpichlo⊇gansuensisendophyte on the disease resistance of drunken horse grass to powdery mildew[J].ActaPrataculturaeSinica, 2015, 24(11): 66-71.
[9]张兴旭, 南志标,李春杰. 内生真菌提高禾草耐重金属胁迫的研究进展[J]. 草业科学, 2014, 31(8): 1 466-1 474.
ZHANG X X,NAN Z B, LI C J. Research progress of improved resistance of the grass to the heavy metal tress by endophyte[J].PrataculturalScience, 2014, 31(8): 1 466-1 474.
[10]ZHANG X X, LI C J, NAN Z B. Effects of cadmium stress on seed germination and seedling growth ofElymusdahuricusinfected with theNeotyphodiumendophyte[J].ScienceChina-LifeSciences, 2012, 55(9): 793-799.
[11]ZHANG X X, LI C J, NAN Z B. Effects of cadmium stress on growth and anti-oxidative systems inAchnatheruminebrianssymbiotic withNeotyphodiumgansuense[J].JournalofHazardousMaterials, 2010, 175(1-3): 703-709.
[12]YOUNG C A, SCHARDL C L, PANACCIONE D G,etal. Genetics, genomics and evolution of ergot alkaloid diversity[J].Toxins, 2015,7(4): 1 273-1 302.
[13]MCCOLLOUGH S F, PIPER E, LOHNSON Z B,etal. Effect of the fescue alkaloids on peripheral blood flow, serum prolactin, and rectal terperatures in steers[J].JournalofAnimalScience,1994, 72(Suppl.2): 31.
[14]SCHARDL C L, PANACCIONE D G, TUDZYNSKI P. Ergot alkaloids-biology and molecular biology[J].Thealkaloids:ChemistryandBiology, 2006, 63: 45-86.
[15]ZHANG X X, LI CH J, NAN ZH B,etal.Neotyphodiumendophyte increasesAchnatheruminebrians(drunken horse grass) resistance to herbivores and seed predators[J].WeedResearch, 2012, 52(1): 70-78.
[16]李春杰, 南志标, 张昌吉,等. 醉马草内生真菌对家兔的影响[J].中国农业科技导报,2009,11(2):84-90.
LI C J, NAN Z B, ZHANG C J,etal. Effects of drunken horse grass infected with endophyte on Chinese rabbit[J].JournalofAgriculturalScienceandTechnology, 2009,11(2):84-90.
[17]鲁璐, 吴瑜.3种微量元素对小麦生长发育及产量和品质的影响研究进展[J].应用与环境生物学报, 2010, 16(3): 435-439.
LU L, WU Y. Advance in study of effects of three trace elements on wheat[J].ChineseJournalofApplied&EnvironmentalBiology, 2010, 16(3): 435-439.
[18]SMITH B N. Iron in higherplants: storage and metabolicrole[J].PlantNutri., 1984, 7: 759-766.
[19]李元芳, 陈婉华. 微量元素及其在牧草生产上的应用[J].中国草地,1991,2:70-73.
LI Y F, CHEN W H. Trace elements and their application to the pasture production[J].GrasslandofChina,1991, 2:70-73.
[20]吴妤, 禹文雅, 李奕松. 缺铁胁迫对草莓幼苗光合特性及细胞器铁含量的影响[J]. 植物营养与肥料学报, 2013, 19(4): 918-925.
WU Y, YU W Y, LI Y S. Effect of iron deficiency stress on phptpsynthetic characteristics and organelle iron content of straberry seedlings[J].JournalofPlantNutritionandFertilizer, 2013, 19(4): 918-925.
[21]刘芷宇, 唐永良, 罗质超. 主要作物营养失调症状图谱[M]. 北京: 中国农业出版, 1982: 47-48.
[22]张正仁, 宋长铣. 微量元素在植物生命活动中的作用[J]. 南京大学学报(自然科学版), 1991, 27(3): 530-539.
ZHANG Z R, SONG C X. The action of micronutrient splayed in plant life[J].JournalofNanjingUniversity, 1991, 27(3): 530-539.
[23]魏宝阳, 曹亮, 李顺祥, 等. 内生菌与药用植物的关系及对次生代谢产物的影响[J]. 中国农学通报, 2011, 27(19): 83-88.
WEI B Y, CAO L, LI S X,etal. The role of endophytes in medical plants and the effect of endophytes on secondary metabolites[J].ChineseAgriculturalScienceBulletin, 2011, 27(19): 83-88.
[24]高嘉卉, 南志标.禾草内生真菌生物碱的研究进展[J].生态学报, 2007, 27(6): 2 532-2 546.
GAO J H, NAN Z B. A review of bioprotective alkaloids of grass fungal endopyte symbioses[J].ActaEcologicaSinica, 2007, 27(6): 2 532-2 546.
[25]代乐英, 黄玺, 李春杰, 等. 麦角生物碱在醉马草内生真菌共生体中的空间分布[J]. 草业学报, 2010, 19(6): 215-221.
DAI L Y, HUANG X, LI C J,etal. Spatial variation of ergot alkaloids in drunken horse grass infected byNeotyphodiumgansuense[J].ActaPrataculturaeSinica, 2010, 19(6): 215-221.
[26]MALINOWSKI D P, BELESKY D P, Hill N Setal. Infilence of phosphorus on the growth and ergot alkaloid content ofNeotyphodiumcoenophialum-infected tall fescue (FestucaarundinaceaSchreb.)[J].PlantandSoil, 1998, 198: 53-61.
[27]张颖, 韩建国. 内生真菌对苇状羊茅和多年生黑麦草影响的研究进展[J]. 草业科学, 2004, (7):59-65.
ZHANG Y, HAN J G. Reviewe about effect of endophytic fungi onFestucaarundinaceaandLoliumperenne[J]PrataculturalScience, 2004, (7):59-65.
[28]田发益, 李晓忠, 何冰梅. 新西兰草甸羊茅中不同处理的黑麦草碱含量分析[J].草业科学, 2009, 26(3): 348-523.
TIAN F Y, LI X Z, HE B M. Loline content of meadow fescue from New Zealand under different treatments[J].PrataculturalScience, 2009, 26(3): 348-523.
[29]胡春霞.氮、磷和色氨酸对醉马草内生真菌共生体生长及麦角生物碱含量的影响[D].兰州:兰州大学, 2013:33-44.
[30]王萍,张兴旭,赵晓静,等. 重金属胁迫对醉马草生长及生理生化指标的影响[J]. 草业科学,2014,31(6):1 080-1 086.
WANG P, ZHANG X X, ZHAO X J,etal. Effects of three kinds of metal ions on growth,physio-biochemical response ofAchnatheruminebrians[J].PrataculturalScience, 2014,31(6):1 080-1 086.
[31]苏加义, 赵红梅. 微量元素对黑麦草产量和品质的影响[J]. 草业科学, 2008, 25(10): 66-69.
SU J Y, ZHAO H M. Effect of different trace element on yield and quality of ryegrass[J].PrataculturalScience, 2008, 25(10): 66-69.
[32]高大文, 文湘华, 周晓燕, 等. 微量元素对白腐真菌的生长影响和抑制酵母菌效果的研究[J]. 环境科学, 2006, 27(8): 1 623-1 626.
GAO D W, WEN X H, ZHOU X Y,etal. Effect of trace element on the growth of white rot fungus and suppressing yeast in liquid medium[J].EnvironmentalScience, 2006, 27(8): 1 623-1 626.
[33]胡春霞, 李秀璋, 方爱国, 等. 色氨酸对醉马草内生真菌共生体麦角生物碱含量的影响[J]. 草业科学, 2014, 31(5): 844-849.
HU C X, LI X Z, FANG A G,etal. Effects of tryptophan on ergot alkaloids concentrations ofAchnatheruminebrians-Neotyphodiumgansuenseassociation[J].PrataculturalScience, 2014, 31(5): 844-849.
[34]李春杰. 醉马草-内生真菌共生体生物学与生态学特性的研究[D]. 兰州: 兰州大学, 2005.
[35]胡晨曦, 杨金玲, 程克棣, 等. 麦角碱生物合成途径中酶学及相关基因研究进展[J]. 西北植物学报, 2005, 25(4): 819-828.
HU C X, YANG J L, CHENG K D,etal. Enzymes and corresponding genes in ergot alkaloid biosynthetic pathways[J].ActaBotanicaBoreali-OccidentaliaSinica, 2005, 25(4): 819-828.
[36]GERHARDS N, NEUBAUER L, TUDZYNSKI P,etal. Biosynthetic pathways of ergot alkaloids[J].Toxins, 2014, 6(12): 3 281-3 295.
[37]TSUI C. The role of zinc in auxin synthesis in the tomato plan[J].AmericanJournalofBotany, 1948: 172-179.
[38]NASON A, KAPLAN N O, COLOWICK S P. Changes in enzymatic constitution in zincdeficientNeurospora[J].JournalofBiologicalChemistry, 1951, 188: 397-406.
[39]高质, 林葆, 周卫. 锌素营养对春玉米内源激素与氧自由基代谢的影响[J]. 植物营养与肥料学报, 2001, 7(4): 424-428.
GAO Z, LIN B, ZHOU W. Effects of znic nutrition on endogenous hormone and free radicals in spring maize[J].PlantNutritionandFertilizerScience, 2001, 7(4): 424-428.
[40]王景安, 柴娜, 梁晓华. 缺锌对不同基因型玉米激素代谢的影响[J]. 华北农学报, 2010, 25(3): 148-152.
WANG J A, CHAI N, LIANG X H. Effects of the hormone metabolism of different maize genotypes under zinc deficiency[J].ActaAgriculturaeBoreali-Sinica, 2010, 25(3): 148-152.
[41]任立民, 刘鹏. 锰毒及植物耐性机理研究进展[J]. 生态学报, 2007, 27(1):357-367.
REN L M, LIU P. Reviewe of Manganese toxicity & the mechanisms of plant tolerance[J].ActaEcologicaSinica, 2007, 27(1):357-367.
[42]CAO G, LIANG M Z. Manganese-trace element necessary for plants at the balance growth system[J].SoilFert., 2004, 1: 2-3.
[43]江振然, 梅兴国, 吴奇君.营养物质和前体对紫杉醇生物合成的影响[J]. 海峡药学, 2002, 14(4): 36-38.
JIANG Z R, MEI X G, WU Q J. Effect of nutrient and precursors on the taxol biosynthesis in cell of suspension of T-cuspidata[J].StraitPharmaceuticalJournal, 2002, 14(4): 36-38.
[44]黄益宗, 朱永官, 黄凤堂, 等. 镉和铁及其交互作用对植物生长的影响[J].生态环境, 2004, 13(3): 406-409.
HUANG Y Z, ZHU Y G, HUANG F T,etal. Effects of cadmium and iron and their interactions on plants growth: a review[J].EcologyandEnvironment, 2004, 13(3): 406-409.
(编辑:裴阿卫)
Ergot Alkaloids Content of Symbiont ofEpichlo⊇gansuensis-Achnatheruminebriansunder Different Mn, Zn and Fe Conditions
WAN Zhiwen, FENG Jiangrong, WANG Ping, LI Chunjie*
(Key Laboratory of Grassland Farming Systems, College of Pastoral Agriculture Science and Technology , Lanzhou University, Lanzhou 730020, China)
A study was conducted to investigate the effects of different concentrations of Mn, Zn and Fe on variations of ergot alkaloids contentin symbiont ofEpichlo⊇gansuensis-Achnatheruminebrians. The results showed that: (1) The contents of ergine and ergonovine under Mn2+5.0 mmol·L-1had a peak value at the 3rd week,which had significant(P<0.05) difference with other Mn2+treatments, but the contents of ergonovine under 0.01 mmol·L-1had a peak value at the 6th and 9th week. (2) The contents of ergine had maximum under Zn2+7.0 mmol·L-1at the 3rd , 6th and 9th week , and the contents of ergine was significantly (P<0.05) higher at the 3rd week than those at the 6th and 9th week. (3) The contents of ergonovine had maximum under Fe2+21.0 mmol·L-1at the 3rd, 6th and 9th week. The contents of ergine had a peak value under 6.0~12.0 mmol·L-1at the 3rd, 6th and 9th week , and the contents of ergine was significantly (P<0.05) higher at the 3rd week than those at the 6th and 9th week. The contents of ergonovine had maximum under Fe2+0.0 mmol·L-1at 3rd and 6th week, but under 0.03 mmol·L-1had a peak value at 9th week. So,high concentrations of Mn, Zn and Fe elements is helpful to stimulate the drunken horse grass infected by endophytic fungi to accumulate tow ergot alkaloid for a short time treatment.
Achnatheruminebrians; endophyte; ergine; ergonovine; Mn;Zn;Fe
1000-4025(2016)07-1427-08
10.7606/j.issn.1000-4025.2016.07.1427
2016-04-11;修改稿收到日期:2016-06-06
国家“973”项目(2014CB138702);国家自然科学基金(31372366);教育部创新团队发展计划(IRT13019)
万志文(1992-),女,在读硕士研究生,主要从事禾草-内生真菌共生体研究。E-mail:wanzhw14@lzu.edu.cn
李春杰,博士,教授,主要从事禾草内生真菌共生体及草类植物病理学研究。E-mail:chunjie@lzu.edu.cn
Q945.79
A
