Seasonal change mediates the shift between resource and pollen limitation in Hedysarum scoparium(Fabaceae)
2015-12-19ChengChenPanQiFengHaLinZhaoXueYongZhaoLinDeLiuJiLiangLiuLiZhangJinLi
ChengChen Pan ,Qi Feng ,HaLin Zhao ,XueYong Zhao ,LinDe Liu ,JiLiang Liu ,Li Zhang ,Jin Li
1.Cold and Arid Regions Environmental and Engineering Research Institute,Chinese Academy of Sciences,Lanzhou,Gansu 730000,China
2.School of Life Sciences,Ludong University,Yantai,Shandong 264025,China
1 Introduction
A large proportion of flowers within a plant do not develop into fruits(Stephenson,1981;Sutherland and Delph,1984).The developed fruit contains few seeds with respect to the number of ovules(Fenster,1991;Campbell and Halama,1993).Limitations in pollen and abiotic resources(e.g.,soil nutrients,water,light,and climatic factors)have been presented to explain these common phenomena in the absence of herbivory or predation(Burd,1995,1998).
Resource and pollen limitations are usually tested by comparing fruit set or seed production of flowers,inflorescences,or individual plants receiving supplemental hand pollination with those of open-pollinated counterparts within a season(Asikainen and Mutikainen,2005).A plant is considered pollen-limited if fruit set or seed production are elevated by hand pollinations relative to natural pollinations.The degree of pollen limitation will affect the role of female mate choice in plants,the opportunity for selection of floral traits,or the possibility of changes in the mating system(Fenster and Ritland,1994).The main reasons for pollen limitation include limitation in pollinator visits,limited pollen availability,inefficient pollen transfer,low pollen-tube survival,and zygote death(Harder and Aizen,2010).
Conversely,a plant is considered resource-limited if seed or fruit set cannot be elevated by hand pollinations.Abiotic resource limitations include inadequate soil nutrients,water,light,and climatic factors(Willson and Burley,1983;Niesenbaum,1993;McIntosh,2002;Gaudeul and Till-Bottraud,2004).
Because of the stochastic environment of pollination and resource availability,pollen and resource limitation may vary over time(Vaughton,1991)or between and within individuals and populations of the same species(Karoly,1992;Casper and Niesenbaum,1993;Dudash,1993).Theory suggests that with sufficient environmental variation,pollen limitation might be observed at some places or times,and resource limitation at others(Campbell,1987).To date,most studies have focused on the availability of pollen and light,nutrient,or water resources(Willson and Burley,1983;Niesenbaum,1993;McIntosh,2002;Gaudeul and Till-Bottraud,2004).To our knowledge,no empirical research exists addressing the effect of seasonal change on the variation of pollen limitation and resource limitation at peak flowering.
Here we evaluate resource limitation or pollen limitation of fruit set and seed production at different stages of peak flowering for a shrub,Hedysarum scoparium(Fabaceae)in the middle reaches of the Hexi Corridor region in Gansu Province of Northwest China.In this region,the flowering season forH.scopariumlasted from late May to late September in 2009(Panet al.,2010).Peak flowering occurred from the middle of August(late summer)to the middle of September(early autumn).Thus,we were able to examine the limits of fruit set and seed production caused by seasonal change.
To estimate resource or pollen limitation of fruit set and seed production,we compared fruit set and seed production under different pollination treatments during both seasons(summer and autumn)in 2010.Our goal was to determine whether seasonal change altered the pollinator-mediated reproductive success ofH.scoparium,and further influenced the shift between pollen limitation and resource limitation.
2 Methods
2.1 Study site
This experiment was conducted in the experimental zone of the Linze Inland River Basin Research Station,Chinese Ecosystem Research Network(39°19'N,100°21'E),in the middle reaches of the Hexi Corridor region in Gansu Province of Northwest China.The study area is located at the southern edge of the Badain Jaran Desert,with an altitude ranging from 1,368 to 1,380 m a.s.l..This region is traversed with dense moving dunes as well as Gobi.This region is characterized by a temperate continental climate:dry and hot in the summer,wetter and cooler in the autumn,and cold in the winter.The average annual precipitation is 117 mm,65% of which falls in the summer;the annual mean evapotranspiration is more than 2,390 mm.The annual mean temperature is 7.6 °C,while the absolute maximum may reach 39 °C and the minimum-27 °C.The mean temperature in the growing season(late May to early October)is 20.4 °C.The plants grown here are principally woody species such asHaloxylon ammodendron,Calligonum mongolicum,Calligonum gobicum,Calligonumpotanini,Calligonumchinense,Caragana korshinskii,H.scoparium,Nitraria sphaerocarpa,andReaumuria soongorica.Except forH.ammodendron,which often grows into small arbors,the others are shrub species.
2.2 Species
Hedysarum scoparium(Fabaceae)is a perennial,large shrub usually located in arid and semi-arid desert regions,and it can grow well as a pioneer plant in shifting sand dunes.It is well known for its ability to live in desertified habitats(Liu and Zhao,2001).This perennial species is self-compatible but not autogamous(Panet al.,2010).It produces racemose inflorescences and nectar-secreted,hermaphroditic flowers with degraded wings that open during daylight hours.Flowering occurs from late May until late September,with flowering peaks in approximately mid-August(Panet al.,2010).During its flowering peak,H.scopariumis virtually the only dominant species in full bloom.
2.3 Experimental design
To study the effect of seasonal change on reproductive output during peak flowering,we separated peak flowering into two stages:the early stage during summer and the late stage during autumn.August 15–25 and September 5–15,2010 were selected to represent the early and late stages,respectively.For this study,one population around the meteorological station was selected(50m×50m)because the same flowering pattern was observed in other populations in this region.To estimate the degree of resource limitation and/or pollen limitation,we performed a pollen-supplementation experiment.We labelled nine plants at the same flowering stage.All plants for the experiment were similar in size(approximately 2 m height).For each plant,two pollination treatments were applied at each stage:(1)cross-pollination(C)in which the flower buds were emasculated,bagged,and hand-pollinated with pollen from individuals located at least 20 m away;and(2)control(N)in which flowers were left to be naturally pollinated by insects.Each treatment was replicated on two to four inflorescences over two to four branches per plant.To avoid the effect of the position-dependent fruit set,the whole inflorescence was pollinated.We pollinated all new receptive flowers from 11:00–15:00 every day.Each inflorescence was covered with gauze bags(20cm×40cm,0.1 mm mesh light).All the experimental inflorescences were regularly monitored until fruit production.
After maturation,we counted the number of experimental flowers that had produced fruits.These fruits were taken to the laboratory for determination of the seed number per fruit.
Students' t-tests were performed to determine the difference in temperature,relative humidity,fruit set,and seed production of the C and N treatments between the two stages.Data from individual flowers belonging to the same treatment and plant were averaged.Weather data were provided by the Linze Inland River Basin Research Station,Chinese Ecosystem Research Network.
3 Results
As shown in Figure 1,the temperature was significantly higher in the early stage peak flowering than in the late stage,and concomitant relative humidity had an opposite trend.In the early stage peak flowering,experimental pollen supplementation did not increase the net reproductive rate of the plants(Figure 2),based on comparisons of the performance of supplemented flowers with those of control(N)flowers.In the late stage peak flowering,experimental pollen supplementation significantly increased the net reproductive rate of the plants(Figure 2).
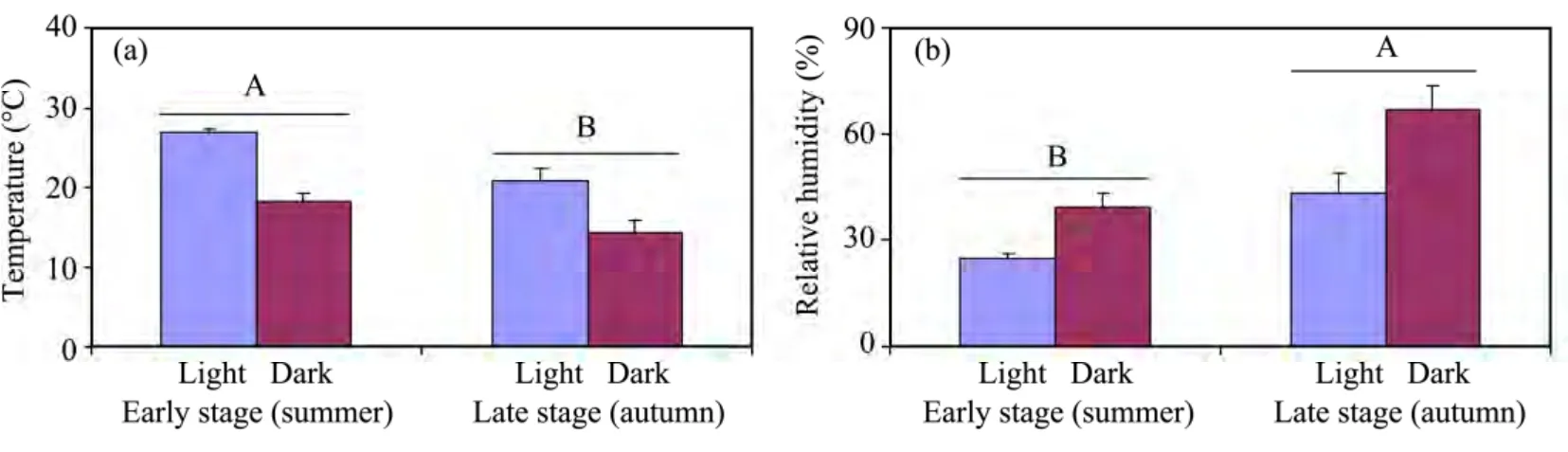
Figure 1 The means of temperature(a)and relative humidity(b)of light and dark in a day during the early and late stage peak flowerings.Vertical bars denote standard errors.Different letters(A,B)indicate significant differences at α <0.05
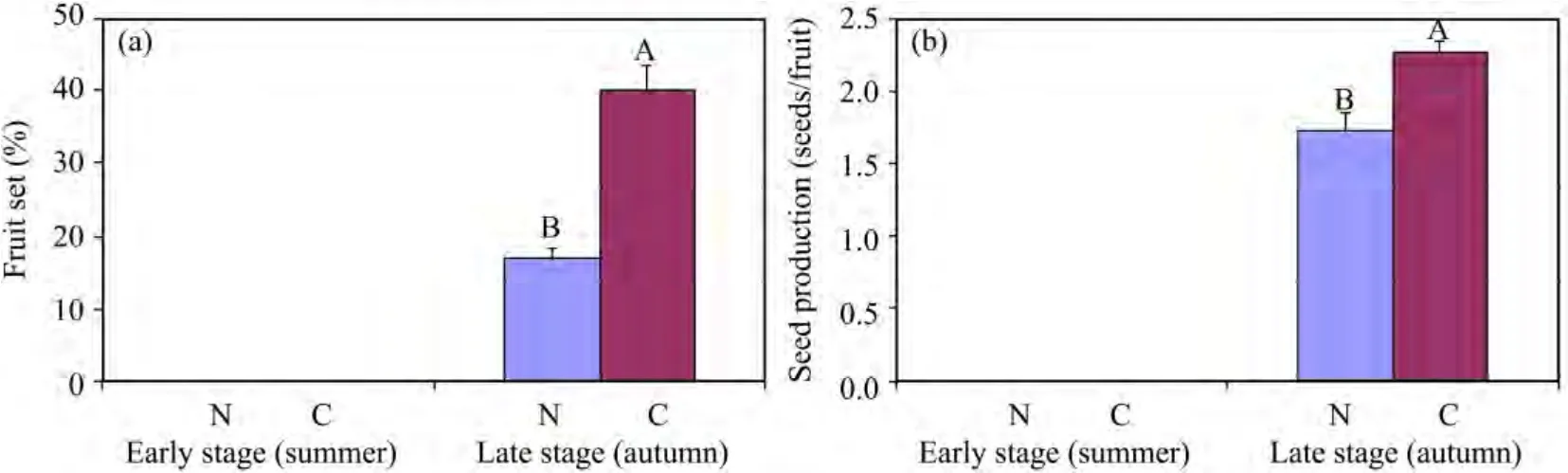
Figure 2 The mean fruit set(a)and seed production(b)under different pollination treatments at the early-and late-stage peak flowerings in H.scoparium.Vertical bars denote standard errors.Pollination treatments:N,control;C,cross-pollination.Different letters(A,B)indicate significant differences at α <0.05
4 Discussion
In the present study,pollen and resource limitations were examined by comparing fruit set and seed production in natural-and hand-pollinated flowers.The responses of reproduction to increased pollen loads suggested that the reproductive success ofH.scopariumwas resource limited in the early stage peak flowering(in fact,H.scopariumwas resource limited during the whole summer),whereas it was pollen limited in the late stage peak flowering.This accords with the findings of Larson and Barrett(2000)and Zimmerman and Aide(1989).The shift between resource limitation and pollen limitation was related to the seasonal variation in temperature and humidity.Differences in air temperature and humidity between both stages(summer and autumn)were major and were clearly associated with the variation in pollen and resource limitation.This phenomenon was observed in otherH.scopariumpopulations in this region in 2009.
Most previous discussions of resource limitation have focused on the fertilizer or water regimes(Ayre and Whelan,1989;Asikainen and Mutikainen,2005;Brookeset al.,2008),with little mention of varying weather conditions.Varying weather conditions,such as variation in temperature and precipitation during the flowering season,are likely to cause variation in pollination success(Dudash and Fenster,1997;Bakeret al.,2000).The climate in the Hexi Corridor region is characterized by seasonal fluctuations in humidity and temperature.In this environment,high temperature and drought stress occurring during the summer of our study period led to almost no fruiting(percentage of fruit set was 0.14%±0.09% for untagged flowers during the summer in 2010).
There are at least three possible ways by which almost no fruiting occurs during summer.First,high temperature and drought stress may affect membrane integrity and germinability of the pollen(Heslop-Harrison,1987).The results of Younget al.(2004)showed that developing pollen had lower germination rates in high temperature conditions.ForH.scopariumin our study,the pollen was of complete fertility(data not shown).According to Younget al.(2004),reduced pollen germinability and disruption of membrane integrity are probably the major contributors to reduced reproductive success in high temperature conditions.Second,successful fertilization requires the maternal guidance of the pollen tube through the maternal sporophytic tissue,to the embryo sac(Rotmanet al.,2003).In high temperature stress conditions,pollen tube growth may begin but cannot be attracted by ovules,so no fertilization takes place(Turner,1993).Third,the impacts of high temperature and drought stress on micropyle penetration,fertilization,and post-fertilization cannot be ruled out.Therefore,the observed resource limitation during summer in our study may be explained by the combined variations in membrane integrity and germinability of the pollen,pollen tube growth,and micropyle penetration during high temperature and drought stress.
Although the fruit set ofH.scopariumapparently became resource limited during the summer,the subsequent flower production was not decreased in the late stage peak flowering.The results of our experimental pollen supplementation showed that the net reproductive rate of the plants increased significantly.These indicated that reproduction ofH.scopariumin this stage was pollen limited.The pollen limitation can be strongly attributed to pollinator visitation frequency(Panet al.,2012).However,pollinator visitation frequency alone may not completely explain the results of pollen limitation.Other factors such as limited pollen availability,inefficient pollen transfer,low pollen-tube survival,and zygote death may also lead to pollen limitation(Harder and Aizen,2010).Though pollen limited in the late stage peak flowering,this fruiting strategy is a safe strategy forH.scopariumto avoid the effects of summer heat and drought to ensure seed production in the Hexi Corridor region,and may facilitate population persistence.
The climate of the Hexi Corridor region is characterized by four distinct seasons:spring,summer,autumn,and winter.The contrast between summer and autumn is striking.Summer is hot and droughty,while autumn is cool and relatively humid.Rainfall during the summer accounts for more than 65% of the annual precipitation.However,the air humidity remains low due to the high evaporation.When the first substantial autumn rain actually occurs,summer ends and autumn begins.There are significant concomitant temperature decreases and relative humidity increases.Most plant species in this region,such asElaeagnus angustifoliaandN.sphaerocarpa,flower in spring and summer,and bear ripe fruit in summer and autumn.ForH.scoparium,flowering occurs from late May until late September.However,before the first autumn rains occurred,we observed almost no fruit and seed during the summer.This suggests that resource limitation of fruit set in summer was a likely response of the plants.In 2009 and 2010,the first autumn rainfalls came on August 25 and September 1,respectively,after which most of the fruit set occurred.
5 Conclusions
The reproductive success ofH.scopariumwas resource limited during summer,whereas it was pollen limited in autumn.The shift between resource limitation and pollen limitation was induced by the fluctuations of temperature and humidity from summer to autumn.The shift timing depended on the dates of the first autumn rainfalls in 2009 and 2010.The first autumn rainfall came on August 25,2009 and on September 1,2010,after which most of the fruit set ofH.scopariumoccurred.The adaptation of pollination to seasonal environmental changes was reported by Dafni(1996),and our study found that the reproductive phenology ofH.scopariumalso showed adaptation to the typical seasonal fluctuations of the climate in the Hexi Corridor region.This selection avoids the effects of summer heat and drought that are considered a limiting factor for the fruiting ofH.scoparium.This plant's autumn fruiting strategy,together with the long flowering season,the numerous flowers produced per plant,and the biological features of the pollen itself,show that the adaptation ofH.scopariumto the actual environmental conditions occurs at several levels.Therefore,the reproductive biology of this species might be affected by future climate changes.
This research was funded by the National Natural Science Foundation of China(31400392,91025002,31370466),the Foundation for Excellent Youth Scholars of CAREERI,CAS(Y451111001),and a General Financial Grant from the China Postdoctoral Science Foundation(2014M552516).
Asikainen E,Mutikainen P,2005.Pollen and resource limitation in a gynodioecious species.American Journal of Botany,92(3):487–494.DOI:10.3732/ajb.92.3.487.
Ayre DJ,Whelan RJ,1989.Factors controlling fruit set in hermaphroditic plants:Studies with the Australian Proteaceae.Trends in Ecology &Evolution,4(9):267–272.DOI:10.1016/0169-5347(89)90197-3.
Baker AM,Thompson JD,Barrett SCH,2000.Evolution and maintenance of stigma-height dimorphism in Narcissus.II.Fitness comparisons between style morphs.Heredity,84(5):514–524.DOI:10.1046/j.1365-2540.2000.00686.x.
Brookes RH,Jesson LK,Burd M,2008.A test of simultaneous resource and pollen limitation inStylidium armeria.New Phytologist,179(2):557–565.DOI:10.1111/j.1469-8137.2008.02453.x.
Burd M,1995.Ovule packaging in stochastic pollination and fertilization environments.Evolution,49(1):100–109.
Burd M,1998."Excess" flower production and selective fruit abortion:A model of potential benefits.Ecology,79(6):2123–2132.DOI:http://dx.doi.org/10.1890/0012-9658(1998)079[2123:EFPASF]2.0.CO;2.
Campbell DR,1987.Interpopulational variation in fruit set:The role of pollination limitation in the Olympic Mountains.American Journal of Botany,74(2):269–273.DOI:10.2307/2444029.
Campbell DR,Halama KJ,1993.Resource and pollen limitations to lifetime seed production in a natural plant population.Ecology,74(4):1043–1051.DOI:http://dx.doi.org/10.2307/1940474.
Casper BB,Niesenbaum RA,1993.Pollen versus resource limitation of seed production:A reconsideration.Current Science,65(3):210–214.DOI:10.1111/j.1469-8137.2007.01997.x.
Dafni A,1996.Autumnal and winter pollination adaptations under Mediterranean conditions.Bocconea,5(1):171–181.
Dudash MR,1993.Variation in pollen limitation among individuals ofSabatia angularis(Gentianaceae).Ecology,74(3):959–962.DOI:http://dx.doi.org/10.2307/1940820.
Dudash MR,Fenster CB,1997.Multiyear study of pollen limitation and cost of reproduction in the iteroparousSilene virginica.Ecology,78(2):484–493.DOI:10.2307/2266024.
Fenster CB,1991.Effect of male pollen donor and female seed parent on allocation of resources to developing seeds and fruit inChamaecrista fasciculata(Leguminosae).American Journal of Botany,78(1):13–23.
Fenster CB,Ritland K,1994.Evidence for natural selection on mating system in Mimulus(Scrophulariaceae).International Journal of Plant Science,155(5):588–596.DOI:10.1086/297197.
Gaudeul M,Till-Bottraud I,2004.Reproductive ecology of the endangered alpine speciesEryngium alpinumL.(Apiaceae):Phenology,gene dispersal and reproductive success.Annals of Botany,93(6):711–721.DOI:10.1093/aob/mch098.
Harder LD,Aizen MA,2010.Floral adaptation and diversification under pollen limitation.Philosophical Transactions of the Royal Society of London(Series B),365(1539):529–543.DOI:10.1098/rstb.2009.0226.
Heslop-Harrison J,1987.Pollen germination and pollen tube growth.International Review of Cytology,107:1–77.
Karoly K,1992.Pollinator limitation in the facultatively autogamous annualLupinus nanus(Leguminosae).American Journal of Botany,79(1):49–56.DOI:10.2307/2445196.
Larson BMH,Barrett SCH,2000.A comparative analysis of pollen limitation in flowering plants.Biological Journal of the Linnean Society,69(4):503–520.DOI:10.1111/j.1095-8312.2000.tb01221.x.
Liu ZM,Zhao WZ,2001.Shifting-sand control in central Tibet.Ambio,30(6):376–380.DOI:10.1639/0044-7447(2001)030[0376:SSCICT]2.0.CO;2.
McIntosh ME,2002.Plant size,breeding system,and limits to reproductive success in two sister species ofFerocactus(Cactaceae).Plant Ecology,162(2):273–288.DOI:10.1023/A:1020329718917.
Niesenbaum RA,1993.Light or pollen-seasonal limitations on female reproductive success in the understory shrubLindera benzoin.Journal of Ecology,81(2):315–323.DOI:10.2307/2261501.
Pan CC,Liu LD,Hou YL,et al.,2010.Flowering characteristics and breeding system ofHedysarum scopariumin the middle reaches of Heihe River.Journal of Desert Research,30(5):1099–1103.
Pan CC,Liu LD,Zhao HL,et al.,2012.Reproduction ofHedysarum scoparium(Fabaceae)in patched habitat is pollen limited,but not just pollinator limited.Journal of Arid Land,4(1):19–28.DOI:10.3724/SP.J.1227.2012.00019.
Rotman N,Rozier F,Boavida L,et al.,2003.Female control of male gamete delivery during fertilization inArabidopsis thaliana.Current Biology,13(5):432–436.
Stephenson AG,1981.Flower and fruit abortion:Proximate causes and ultimate functions.Annual Review of Ecology and Systematics,12:253–279.DOI:10.1146/annurev.es.12.110181.001345.
Sutherland S,Delph LF,1984.On the importance of male fitness in plants:Patterns of fruit-set.Ecology,65(4):1093–1104.DOI:http://dx.doi.org/10.2307/1938317.
Turner LB,1993.The effect of water stress on floral characters,pollination and seed set in white clover(Trifolium repensL.).Journal of Experimental Botany,44(7):1155–1160.DOI:10.1093/jxb/44.7.1155.
Vaughton G,1991.Variation between years in pollen and nutrient limitation of fruit-set inBanksia spinulosa.Journal of Ecology,79(2):389–400.
Willson MF,Burley N,1983.Mate Choice in Plants.Princeton,NJ:Princeton University Press.
Young LW,Wilen RW,Bonham-Smith PC,2004.High temperature stress ofBrassica napusduring flowering reduces micro-and megagametophyte fertility,induces fruit abortion,and disrupts seed production.Journal of Experimental Botany,55(5):485–495.
Zimmerman JK,Aide TM,1989.Patterns of fruit production in a neotropical orchid:Pollinator vs.resource limitation.American Journal of Botany,76(1):67–73.DOI:10.2307/2444775.
杂志排行

Sciences in Cold and Arid Regions的其它文章
- Toward sustainable desertification reversion:A case study in Horqin Sandy Land of northern China
- Land desertification and restoration in Middle East and North Africa(MENA)region
- Effects of land-use changes on organic carbon in bulk soil and associated physical fractions in China's Horqin Sandy Grassland
- Adaptation to climate change in desertified lands of the marginal regions in Egypt through sustainable crop and livestock diversification systems
- A mechanism for the origin and development of the large-scale dunefield on the right flank of the lower reach of Laoha River,Northeast China
- Characteristics of high arsenic groundwater in Hetao Basin,Inner Mongolia,northern China