菠萝蜜花粉萌发及胚发育的显微观察
2015-07-13李映志余庆李洪波吴钿叶春海
李映志,余庆,李洪波,吴钿,叶春海
(广东海洋大学农学院,广东 湛江 524088)
菠萝蜜(Artocarpus heterophyllus Lam.),又名木菠萝、树菠萝,属桑科桂木属常绿乔木,其果大、果肉爽脆、香气浓郁,含有较多的碳水化合物,是著名的热带水果之一[1]。菠萝蜜起源于印度,1 000多年前引种至中国,在广东、广西、海南和云南等地均有分布和栽培[2]。
菠萝蜜的花为单性花,雌雄同株,果实为雌花序发育而来的聚复果[3],花序中的小花可发育形成果苞或果腱,而果苞的数量及分布与果实的大小及果实是否畸形密切相关。尽管国内外学者在菠萝蜜的种质资源[4–6]、品质生理[7–11]、形态特征[3,12–13]以及加工利用[14–15]等方面开展了较为详细的研究,但有关菠萝蜜受精和胚发育过程的研究少见报道,而菠萝蜜受精和胚发育过程是研究菠萝蜜果实、果苞和果腱形成机理及菠萝蜜畸形果发生原因的基础。为此,笔者采用荧光染色和整体透明技术结合显微切片技术,观察菠萝蜜受精及胚发育的过程,以期为菠萝蜜的栽培及育种研究提供依据。
1 材料和方法
1.1 材料
试验材料选用位于广东省湛江市南亚热带作物研究所内的菠萝蜜种质圃(E21°08′、N110°16′,海拔45 m)的‘GHsj12S45’ 种质,收集于雷州半岛地区,8年生嫁接植株,砧木为湿苞种子实生苗。正常开花结果,常规管理。
1.2 方法
1.2.1 花粉萌发与花粉管生长过程观察
分别在人工自交授粉后1、2、4、6、12、24、48、72、96 和120 h 采集雌花序,按长、宽约2 cm切块后,用FAA 液固定。花柱的压片参照李和平[16]的方法并略作改动:雌花序块固定24 h 后剥出雌蕊,蒸馏水冲洗3次后,用1 mol/L 的NaOH 于45℃水浴下软化15min,用0.1%的苯胺蓝染液染色3 h,蒸馏水洗去浮色和杂质后压片,最后显微镜下观察拍照。
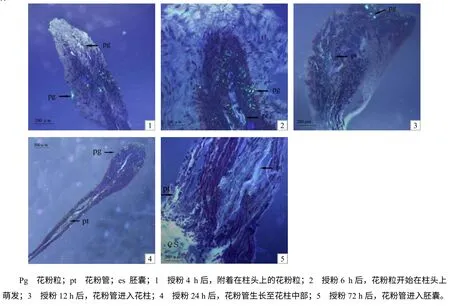
图1 菠萝蜜花粉萌发及花粉管生长进程 Fig.1 Pollengermination and pollen tubegrowth of jackfruit
1.2.2 受精及胚发育过程观察
采集自然授粉下菠萝蜜谢花后0、5、9、13、16、18、21、26 d 的雌花序,按长、宽约2 cm 切块后,用FAA 固定液固定。采用透明整染法[17]和石蜡切片法[16]观察受精及胚发育的过程。
1.2.3 显微照像
所有制片后的材料均在Olympus BX51 型显微镜下观察,DP71 冷CCD 芯片成像,DP–BSW 软件采集图像。
2 结果与分析
2.1 花粉萌发与花粉管生长
菠萝蜜的花粉及花粉管经0.1%水溶性苯胺蓝染色后,在荧光显微镜下发出黄绿色荧光。授粉1 h后,可见有菠萝蜜花粉粒黏附在柱头表面,随着时间的延长,柱头上黏附的花粉粒增加1~5 倍,但至4 h 时仍未见有花粉粒萌发(图1–1);授粉6 h 后,花粉粒开始萌发,花粉管伸长(图1–2);授粉12 h 后,柱头上可见视野内90%以上花粉粒萌发形成花粉管(图1–3);授粉后24~48 h,在所观察的柱头中,有86.3%的柱头可见有花粉管生长至花柱中部(图1–4)。授粉72 h 后,花粉管进入胚囊(图1–5)。
2.2 菠萝蜜受精及胚发育过程观察结果
2.2.1 菠萝蜜成熟胚囊的结构
菠萝蜜的胚珠为典型的倒生胚珠。谢花当天,可见有发育完全的胚囊。胚囊腔外层为珠心,合点端的珠心细胞多且厚,染色较深,近珠孔端的珠心细胞少,染色较浅。珠心外层细胞与珠被绒毡层排列紧密,仅珠孔端分开(图2–1)。在胚囊中,靠近合点端的3个反足细胞呈品字形排列(图2–2);近珠孔端为卵器,2个助细胞与1个较大的卵细胞紧靠在一起,呈三角形排列(图2–3);胚囊中央是2 核的中央细胞(图2–4)。菠萝蜜大多数胚囊的2个极核在受精前融合成一个二倍体次生核(图2–5)。
2.2.2 菠萝蜜胚及胚乳的发育过程
谢花后第5天,受精极核变大,细胞质变浓(图2–6),此时初生胚乳细胞和合子细胞均未分裂,反足细胞完全退化。谢花后第9天,受精极核开始分裂,并随即产生细胞壁,形成胚乳细胞(图2–7);此时合子也开始分裂(图2–8),横向分裂成远珠孔端的顶细胞和近珠孔端的基细胞。
谢花后第13天,胚囊体积增大,胚乳细胞继续分裂,呈长条形,占据大部分胚囊(图2–9)。谢花后第16天,顶细胞经过多次分裂,细胞排列紧密,胚发育到达球形胚时期,与周围的胚乳细胞间存在间隙,同时,胚乳细胞也不断分裂、增多,分布在胚囊壁的内侧周围(图2–10)。
谢花后第18天,胚细胞继续分裂,细胞排列紧密,球形胚顶部两侧的细胞分裂较中间快,形成2个突起,进入早期心形胚阶段(图2–11)。
谢花后第21天,大部分胚发育至子叶胚阶段,两片子叶大小不等,子叶间形成胚芽,胚乳仍充满整个胚囊(图2–12)。
2.2.3 菠萝蜜胚形成中的败育现象
本研究中共观察到以下5 种菠萝蜜胚形成中的败育现象。
谢花后当天,在雌花序中,除正常胚囊外,可见有部分胚珠未形成胚囊(图3–1),另外一些胚珠虽能形成胚囊,但胚囊腔内无任何细胞结构(图3–2)。
谢花后第5天,雌花序中仍存在3 种不正常胚囊。第1 种为空腔胚囊,胚囊腔内无任何细胞结构;第2 种为胚囊退化、败育,即胚囊腔内留下“线状”痕迹,或者胚囊腔内被珠心细胞“充满”(图3–3);第3 种为胚珠败育,珠柄端胚珠组织皱缩程度较高,整个胚珠呈黄褐色,苏木精染液染不上色(图3–4)。
谢花后第9天至第13天,雌花序中除上述3种不正常胚囊外,还有大量未受精胚囊,中央细胞清晰可见(图3–5)。
谢花后第16天,肉眼下正常胚囊体积明显大于败育胚囊,此时,雌花序中仍存在未受精胚囊,其中央细胞仍可见。
谢花后第18天,未受精胚囊的腔内细胞解体,形成中空胚囊(图3–6)。
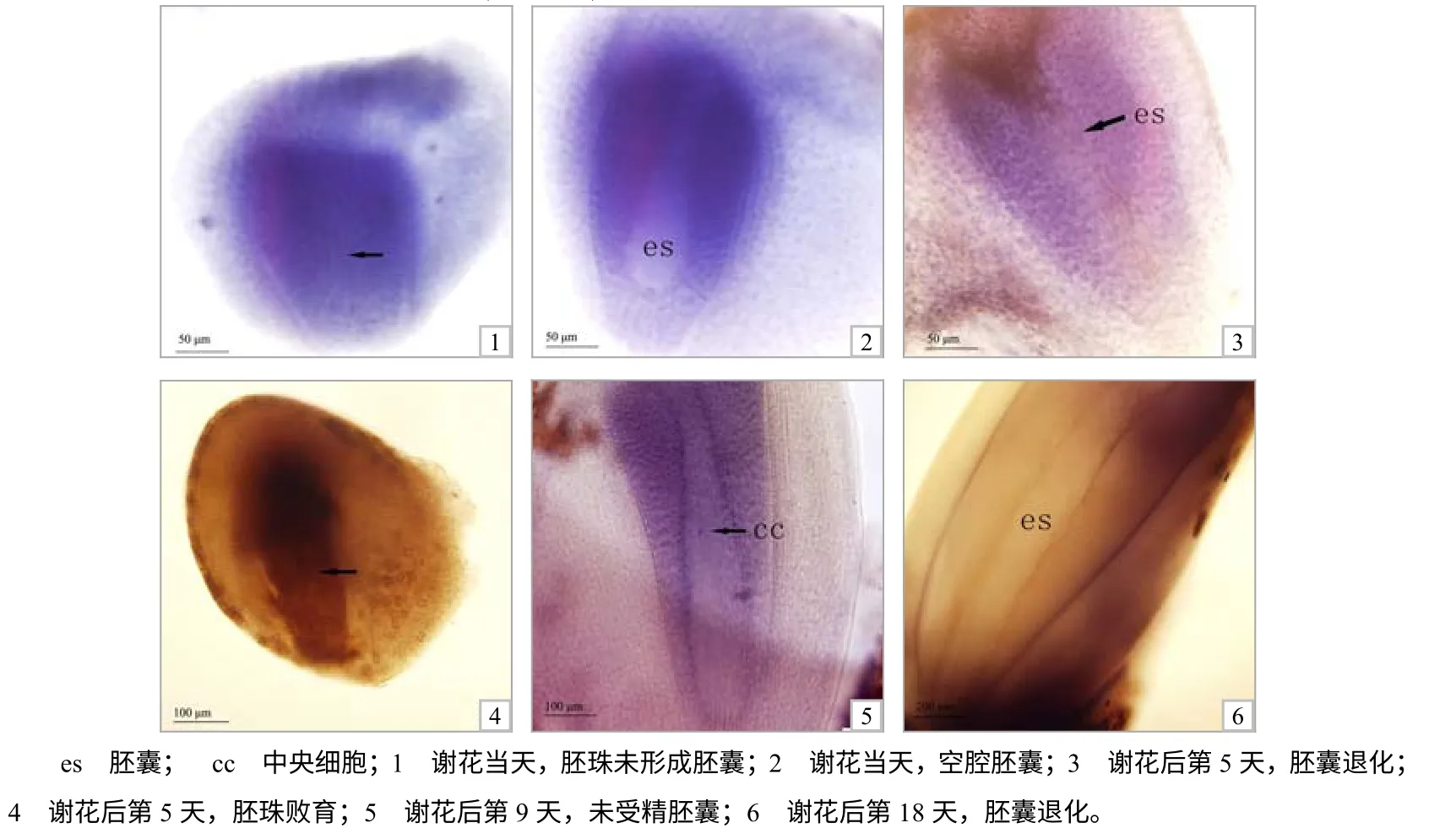
图3 菠萝蜜胚体的败育现象 Fig.3 Abortion in embryo fomation of jackfruit
3 结论与讨论
菠萝蜜花粉粒黏附在柱头上6 h 后才开始萌发,授粉72 h 后,花粉管伸入子房。然而许多植物如猕猴桃[18]、腊梅[19]、桔梗[20]等,授粉2 h 后花粉粒就已开始萌发,授粉后24~36 h 内,花粉管已伸入子房内。菠萝蜜与其他植物相比,其花粉萌发及花粉管伸长较为缓慢,这可能是由于物种间差异或其他原因造成的,其内在机理有待进一步研究。
由于菠萝蜜是聚复果,1个雌花序由多个雌花构成,每朵雌花的子房很小,因此,常规的石蜡切片法很难观察菠萝蜜胚囊的发育以及受精后胚的早期发育过程。透明整染法是观察胚囊及胚发育的重要手段[17]。本研究的观察结果表明,菠萝蜜的成熟胚囊是由8 核7 细胞构成,胚囊在受精前中央细胞融合成1个二倍体次生核,这与番木瓜[21]、荔枝[22]等的胚囊发育类型一致。同一菠萝蜜雌花序中,不同小花的胚囊发育程度不同。谢花当天的雌花序中,部分胚珠已形成成熟胚囊,而部分发育较迟的胚珠还未形成成熟胚囊,直到谢花后第5天,仍有未成熟胚囊存在。
菠萝蜜的胚囊双受精后,经过短暂的休眠,至谢花后第9天,初生胚乳核先于合子分裂,然后合子才首次分裂成顶细胞和基细胞。菠萝蜜的胚乳发育类型为细胞型胚乳。谢花后第16天,胚发育到达球形胚阶段,胚乳细胞继续分裂,且液泡化。谢花后第18天,胚发育至心形胚阶段。谢花后第21天,胚发育至子叶胚阶段。同一雌花序中,不同小花的胚发育过程不完全一致。
本试验观察到了4 种导致胚败育的现象:空腔胚囊、退化胚囊、胚珠败育以及胚囊未受精。它们与菠萝蜜果苞及果腱发育的关系密切,但其产生的原因还有待进一步研究。Farid 等[23]的研究表明,喷硼可以减少菠萝蜜畸形果的发生率,硼可能影响了菠萝蜜的花粉萌发力或胚败育率。
[1] 谭乐和,王令霞,朱红英.菠萝蜜的营养物质成分与利用价值[J].广西热作科技,1999(2):19–20.
[2] 叶春海,王耀辉,李映志,等.菠萝蜜遗传多样性的ISSR 分析[J].果树学报,2009,26(5):659–665.
[3] 吴钿,丰锋,叶春海.菠萝蜜叶和花的解剖结构研究[J].广西植物,2008,28(6):746–749.
[4] 叶春海,李映志,丰锋.雷州半岛菠萝蜜种质遗传多样性RAPD 分析[J].园艺学报,2005,32(6):1088–1091.
[5] Schnell R J,Olano C T,Campbell R J S,et al.AFLP analysis ofgenetic diversity within a jackfruitgerm plasm collection [J].Sci Horti,2001,91(3/4):261–274.
[6] 王艳红,李映志,丰锋,等.磁珠富集法分离菠萝蜜(Artocarpus heterophyllus)微卫星标记及系列分析[J].果树学报,2010,27(6):1046–1051.
[7] 纳智.菠萝蜜中香气成分分析[J].热带亚热带植物学报,2004,12(6):538–540.
[8] Ong B T,Nazimah S A H,Tan C P,et al.Analysis of volatile compounds in five jackfruit (Artocarpus heterophyllus L.) cultivars using solid-phase microextrac- tion (SPME) andgas chromatography-time-of-flight mass spectrometry(GC–TOFMS)[J].J Food Compos Anal,2008,21:416–422.
[9] 李洪波,刘胜辉,李映志,等.顶空固相微萃取和气相色谱–质谱法测定菠萝蜜果肉中的香气成分[J].热带作物学报,2013,34(4):755–763.
[10] Rahman A K M M,Huq E,Mian A J,et al.Microscopic and chemical changes occurring during the ripening of two forms of jackfrtui (Artocarpus heterophyllus Lam.) [J].Food Chem,1995,52:405–410.
[11] 李映志,董黎梨,汪永保,等.干、湿苞木菠萝果实成熟过程中糖代谢特性[J].果树学报,2014,31(1):85–90.
[12] 吴钿,叶春海,丰锋,等.菠萝蜜花药发育及花粉萌发研究[J].植物研究,2011,31(2):169–174.
[13] 唐为萍,陈树思.菠萝蜜次生木质部导管分子观察[J].果树学报,2005,22(6) :725– 727.
[14] 谭乐和,郑维全.菠萝蜜种子淀粉提取及其理化性质测定[J].海南大学学报:自然科学版,2000,18(4):388–390.
[15] Jagtap U B,Waghmare S R,Lokhande V H,et al. Preparation and evaluation of antioxidant capacity of jackfruit (Artocarpus heterophyllus Lam.) wine and its protective role against radiation induced DNA damage [J].Ind Crop Produc,2011,34:1595–1601.
[16] 李和平.植物显微技术[M].北京:科学出版社,2009:216–225.
[17] 杨弘远.用整体染色与透明技术观察胚囊、胚、胚乳和胚状体[J].植物学报,1986(6):575–578.
[18] 梁铁兵,母锡金.美味猕猴桃和软枣猕猴桃种间杂交花粉管行为和早期胚胎发生的观察[J].植物学报,1995,37 (8):607–612.
[19] 王文鹏,周莉花,刘华红,等.夏蜡梅与美国蜡梅属间杂交障碍的组织学机理[J].园艺学报,2013,40(10):1943–1950.
[20] 魏建和,马凤杰,黄璐琦,等.桔梗花粉管萌发的荧光显微观察[J].中国中药杂志,2007,32(16):1625–1627.
[21] 邱德勃,陆锋,谢石文.用酶解压片法分离巴西橡胶树和番木瓜的胚囊[J].植物学报,1991,35(5):350–355.
[22] 叶秀粦,王伏雄,钱南芬.荔枝的胚胎学研究[J].云南植物研究,1992,l4(1):59–65.
[23] Farid A T M,Halder N K,Shahjahan M.Effect of boron for correcting the deformed shape and size in jackfruit [J].Ind J Horti,2007,64:144–149.
