杨树2n花粉形态判别方法的比较1)
2015-03-10杨倩陈欣张平冬康向阳
杨倩 陈欣 张平冬 康向阳
(北京林业大学,北京,100083) (林木育种国家工程实验室(北京林业大学))
杨树2n花粉形态判别方法的比较1)
杨倩 陈欣 张平冬 康向阳
(北京林业大学,北京,100083) (林木育种国家工程实验室(北京林业大学))
采用醋酸洋红染色法,在研究胡杨和毛白杨不同无性系间花粉粒形态差异的基础上,研究不同形态判别标准条件下2n花粉发生频率调查的准确性。研究结果表明,胡杨花粉粒的平均直径为29.7 μm,比毛白杨花粉粒的平均直径32.4 μm略小。胡杨或毛白杨无性系对花粉粒直径影响显著。通过调查四分体时期四分体、三分体以及二分体的比例,可估算2n花粉的理论频率。胡杨和毛白杨各无性系2n花粉的理论频率与以2n花粉粒直径大于花粉粒平均直径的1.28倍为标准时统计的2n花粉实际频率差异不显著,而与以2n花粉粒直径大于37.0 μm为标准时统计的2n花粉实际频率差异极显著。这说明以37.0 μm作为判别胡杨或毛白杨2n花粉形态判别标准会导致较大的误差。而以各无性系花粉粒平均直径的1.28倍作为2n花粉形态判别标准更加科学合理。
胡杨;毛白杨;2n花粉;形态判别;花粉粒直径
Based on the differences of pollen grain shape among clones inPopuluseuphraticaandPopulustomentosa, we studied the accuracy of estimate in percentage of 2npollen determined under different standards. The average diameter was 32.4 μm forP.tomentosa, which was slightly larger than that ofP.euphratica(29.7 μm). Clonal effect was significant for diameter of pollen grains. The theoretical percentage of 2npollen could be estimated based on the rates of tetrad, triad and dyad at tetrad stage. However, the actual percentages of 2npollen could be achieved according to diameter from two standards of 2npollen beyond 37.0 μm and diameter of 2npollen as 1.28 times as that of normal pollen inP.euphraticaandP.tomentosa, respectively. Byχ2test, the estimate of the theoretical percentage of 2npollen agreed with the actual percentages of 2npollen that was estimated by the first standard. However, there was significant difference between the estimate of the theoretical percentage of 2npollen and the actual percentages of 2npollen that was estimated by the second standard. Therefore, the first standard for discrimination of 2npollen (37.0 μm) would result in great estimate error. It is more reasonable for taking the another standard in determining the percentage of 2npollen.
自1936年瑞典Nilsson-Ehle[1]发现巨型欧洲山杨被证明为天然三倍体以来,三倍体育种已成为杨树遗传改良的重要方法,日益受到树木育种学家们的关注。现已在欧洲山杨[2]、美洲山杨[3]、银白杨[4]、香脂杨[5]以及毛白杨[6]等树种中均发现了天然杨树三倍体的存在。近年研究发现,我国广泛栽培的‘I-214’、‘中林46’、‘沙兰杨’、‘廊坊3号杨’、‘武黑1号’、‘辽河杨’等著名杨树品种实际上都是来源于天然2n花粉的三倍体[7-8]。由此可见,杨树三倍体已在我国工业用材林培育、生态环境保护等方面发挥了重要作用。
利用天然2n花粉与单倍性雌配子杂交是选育杨树三倍体的一条重要途径,实现了花粉染色体加倍带来的倍性效应与杂交产生的杂种优势的结合。目前,在毛白杨(P.tomentosa)[9-10]、响叶杨(P.adenopoda)[11]、通辽杨(P.mionii×P.nigra‘Tong liao’)[12]、香脂杨(P.balsamifera)[13]以及灰杨(P.canescens)[14]等杨属树种中都发现有天然未减数2n花粉的存在。由于杨树花粉的离体萌发率低,致使花粉染色体计数困难,2n花粉的产生与否往往是通过测量花粉粒的直径来判别[9]。但由于杨属的不同派间或同一派内的不同种间花粉粒直径存在较大差异,致使天然2n花粉的发生频率的统计存在困难,往往难以准确统计。本研究针对这一难题,选用胡杨和毛白杨为研究对象,通过调查四分体时期四分体、三分体以及二分体的比例,估算天然2n花粉的理论频率,与不同形态判别标准条件下实际统计出的2n花粉频率进行卡方检验(χ2),筛选出杨树天然2n花粉的形态判别方法,对于推动杨树三倍体育种具有重要的理论与实践意义。
1 材料与方法
胡杨雄花枝采自内蒙古自治区额济纳旗东河乌兰图格地区的胡杨天然林;毛白杨雄花枝采自山东国有冠县苗圃的毛白杨基因库,塑料布包裹后运回北京林业大学。
将胡杨、毛白杨雄花枝移入北京林业大学温室进行水培,温室温度控制10~25 ℃。每隔3~4 h采集雄花芽,去除鳞片后用卡诺固定液进行固定,利用醋酸洋红压片法,实时观察小孢子母细胞减数分裂进程。待观察到花粉母细胞发育至四分体时期,分别无性系固定4~8个雄花芽。胡杨和毛白杨花药在冰箱中-4 ℃固定24 h后,转移至70%的酒精中保存待用。
四分体时期四分体、三分体以及二分体比例的调查采用醋酸洋红染色压片法,制片参考康向阳[10]的方法:取在卡诺固定液(V(乙醇)∶V(乙酸)=3∶1)中固定的花芽中部小花,置于洁净载玻片上,除去花盘,滴加1~2滴醋酸洋红,用镊子夹碎花药制作涂片,盖上盖玻片,经大拇指挤压后进行镜检、照相。分别不同无性系观察550个以上的花粉母细胞,分别计算出四分体、三分体以及二分体所占的比例。依据公式E=((2×二分体数目+三分体数目)/(2×二分体数目+3×三分体数目+4×四分体数目))×100%便可以求得天然2n花粉的理论频率。
当雄花序发育成熟,花药开裂后,收集胡杨和毛白杨花粉,经醋酸洋红染色后,在Olympus BX51光学显微镜下观察,采用Olympus DP70照相系统进行数码照相以及花粉粒直径测量。每个无性系测量500个以上的花粉粒,试验重复3次。天然2n花粉的频率分别按以下方法统计:①花粉粒直径大于37 μm的花粉视为天然2n花粉;②假设减数分裂过程中,花粉母细胞的体积不变,则四分体可以形成4个花粉粒,而二分体可以形成2个天然2n花粉粒,则可以计算出天然2n花粉粒的直径约为正常花粉粒直径的1.28倍,大于该无性系花粉粒平均直径1.28倍的花粉粒可视为天然2n花粉。天然2n花粉频率为2n花粉占观察花粉粒总数的百分比。
花粉粒直径数据采用SPASS 18.0软件包进行方差分析和多重比较。变异系数=(标准差/平均值)×100%。
2 结果与分析
2.1 胡杨花粉形态特征与天然2n花粉频率
胡杨成熟花粉粒经醋酸洋红染色后,在光学显微镜下测量花粉粒直径,计算出各无性系花粉粒的形态特征值(见表1)。从表1可以看出,胡杨花粉粒的平均直径为29.7 μm。不同胡杨无性系间花粉粒直径存在一定的差异,其中以无性系HY14的花粉粒最大,其平均直径为33.0 μm,花粉粒直径的变化范围介于23.2~44.5 μm,变异系数为10.6;无性系HY28的平均花粉粒最小,平均直径仅26.5 μm,花粉粒直径的变化范围介于22.5~47.7 μm,变异系数为10.9%。不同无性系间花粉粒直径的方差分析结果表明,无性系对胡杨花粉粒大小的影响达到了显著水平。无性系HY14、HY20、HY34、HY23、HY33以及HY8的花粉粒直径显著大于无性系HY28、HY24以及HY25的花粉粒直径。
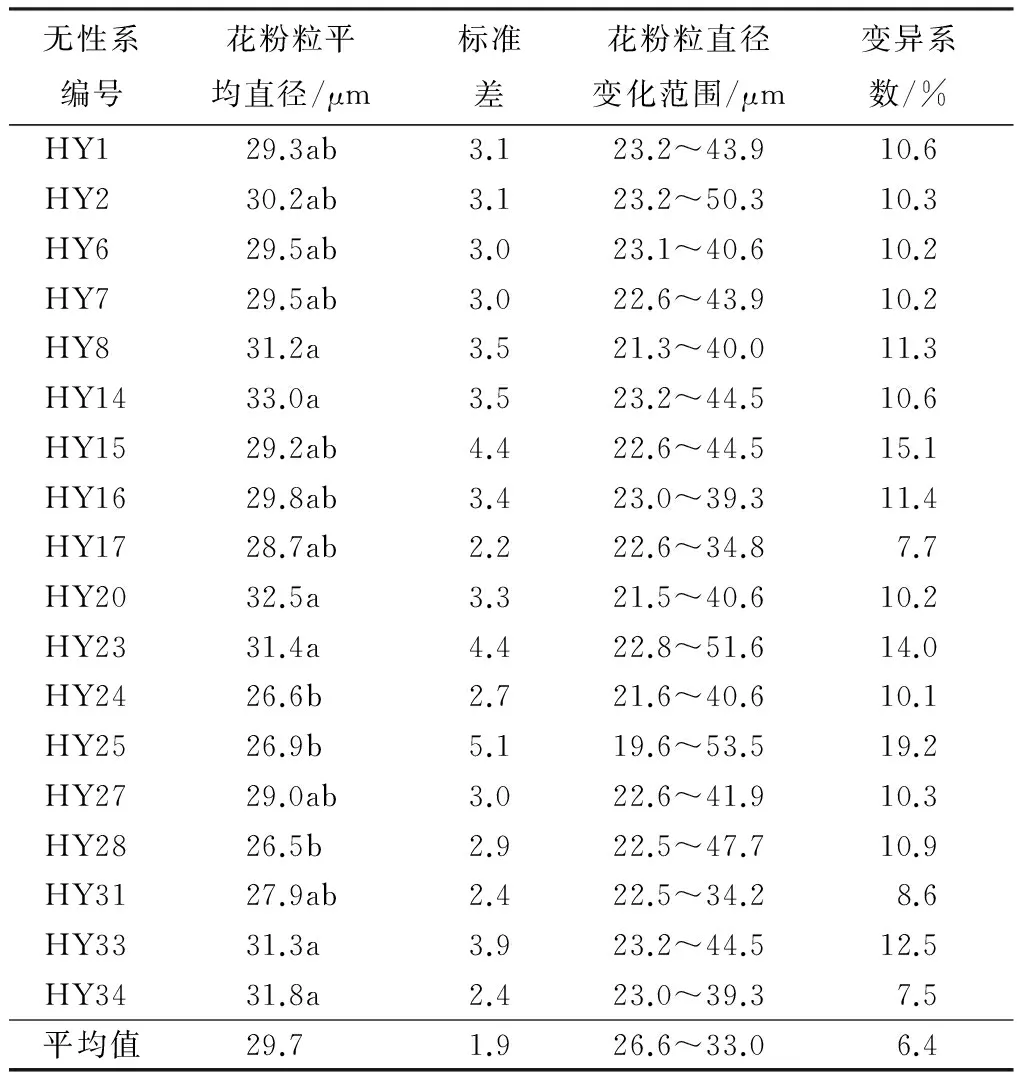
表1 不同胡杨无性系的花粉形态特征值
注:同列不同小写字母表示在0.05水平差异显著。
一般来说,花粉母细胞进入四分体时期后,四分体可以形成四个单倍性花粉粒;三分体形成一个2n花粉粒和两个单倍性花粉粒;二分体形成两个2n花粉粒。因此,根据胡杨减数分裂四分体时期四分体、三分体以及二分体的比率,可以计算出胡杨产生2n花粉的理论频率(见表2)。不同胡杨无性系2n花粉的理论频率在0.19%~3.85%之间变化。另外,分别根据花粉粒直径大于37.0 μm与大于平均花粉粒直径的1.28倍两种标准统计出胡杨各无性系花粉中天然2n花粉的实际频率(见表2)。从表2可以看出,当采用2n花粉粒直径大于37.0 μm为标准时,胡杨各无性系的2n花粉发生的实际频率在0~10.2%变动;当采用2n花粉粒直径大于花粉粒平均直径的1.28倍为标准时,无性系的2n花粉发生的实际频率稍小,介于0~4.37%之间。卡方(χ2)检验结果表明,胡杨各无性系2n花粉的理论频率与以2n花粉粒直径大于花粉粒平均直径的1.28倍为标准时统计的2n花粉实际频率差异不显著,而与以2n花粉粒直径须大于37.0 μm为标准时统计的2n花粉实际频率差异极显著。
表2 胡杨减数分裂四分体时期的小孢子群与天然2n花粉频率
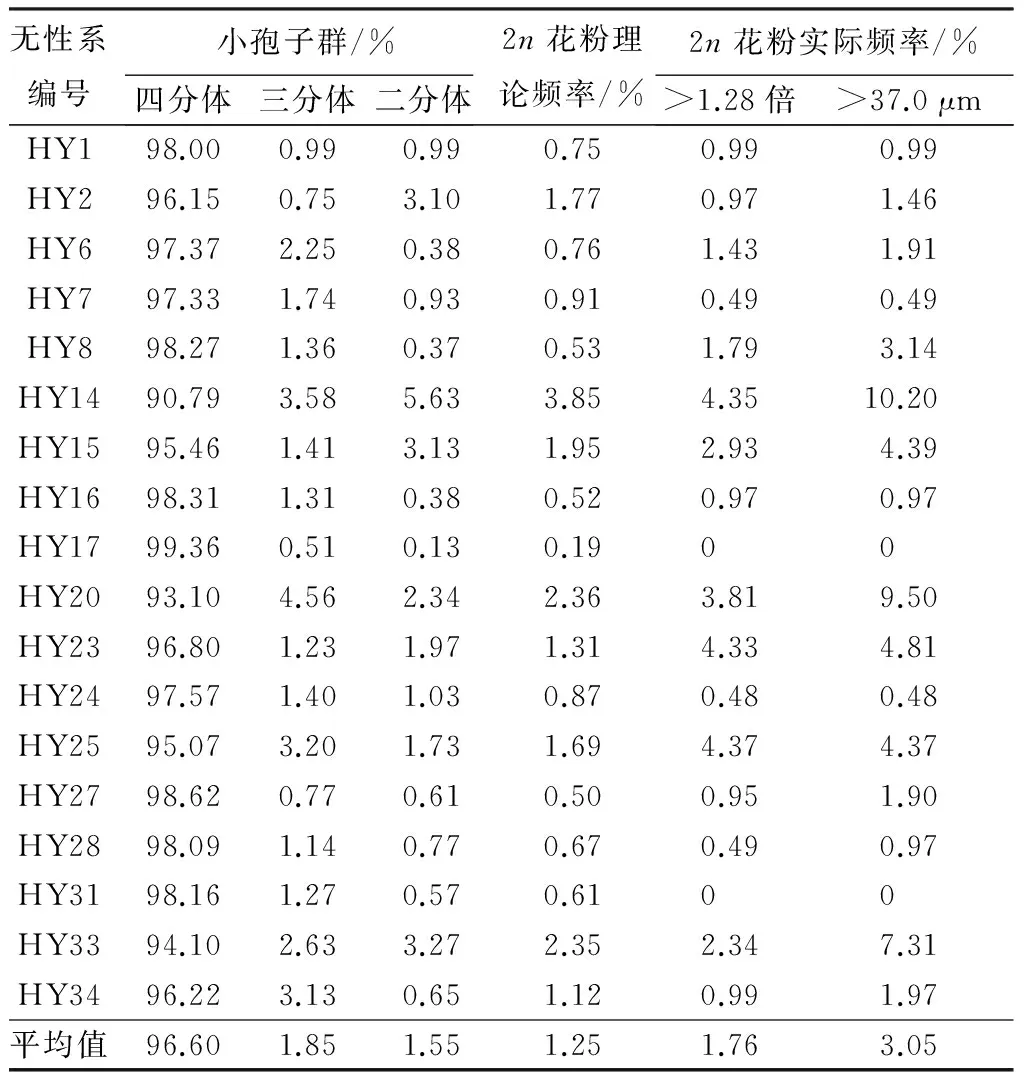
无性系编号小孢子群/%四分体三分体二分体2n花粉理论频率/%2n花粉实际频率/%>1.28倍>37.0μmHY198.000.990.990.750.990.99HY296.150.753.101.770.971.46HY697.372.250.380.761.431.91HY797.331.740.930.910.490.49HY898.271.360.370.531.793.14HY1490.793.585.633.854.3510.20HY1595.461.413.131.952.934.39HY1698.311.310.380.520.970.97HY1799.360.510.130.1900HY2093.104.562.342.363.819.50HY2396.801.231.971.314.334.81HY2497.571.401.030.870.480.48HY2595.073.201.731.694.374.37HY2798.620.770.610.500.951.90HY2898.091.140.770.670.490.97HY3198.161.270.570.6100HY3394.102.633.272.352.347.31HY3496.223.130.651.120.991.97平均值96.601.851.551.251.763.05
注:花粉粒平均直径大于1.28倍时χ2=18.71;花粉粒直径大于37.0 μm时χ2=80.18。
2.2 毛白杨花粉形态特征与天然2n花粉频率
毛白杨成熟花粉粒经醋酸洋红染色后,在光学显微镜下测量花粉粒直径,计算出各无性系花粉粒的形态特征值(见表3)。从表3可以看出,毛白杨花粉粒的平均直径为32.4 μm。不同毛白杨无性系间花粉粒直径存在较大的差异,其中以无性系‘抱头白’的花粉粒最大,其平均直径为37.5 μm,花粉粒直径的变化范围介于21.6~51.6 μm,变异系数为13.6%;无性系3105的平均花粉粒最小,平均直径仅29.4 μm,花粉粒直径的变化范围介于22.6~42.6 μm,变异系数为13.3%。不同无性系间花粉粒直径的方差分析结果表明,无性系对毛白杨花粉粒大小的影响达到了显著水平。‘抱头白’以及无性系5088的花粉粒直径显著大于参试无性系的花粉粒直径。
根据毛白杨减数分裂四分体时期四分体、三分体以及二分体的比率,可以计算出毛白杨产生2n花粉的理论频率(见表4)。不同毛白杨无性系天然2n花粉的理论频率在0~7.05%变化。与此同时,分别根据花粉粒直径大于37.0 μm与大于平均花粉粒直径的1.28倍两种标准统计出毛白杨各无性系花粉中天然2n花粉的实际频率(见表4)。从表4可以,当采用2n花粉粒直径大于37.0 μm为标准时,胡杨各无性系的2n花粉发生的实际频率在1.75%~56.46%之间变动;当采用2n花粉粒直径大于花粉粒平均直径的1.28倍为标准时,无性系的2n花粉发生的实际频率介于0~4.48%。卡方(χ2)检验结果表明,毛白杨各无性系2n花粉的理论频率与以2n花粉粒直径须大于花粉粒平均直径的1.28倍为标准时统计的2n花粉实际频率差异不显著,而与以2n花粉粒直径须大于37.0 μm为标准时统计的2n花粉实际频率差异极显著。

表3 不同毛白杨无性系花粉形态特征值
注:同列不同小写字母表示在0.05水平差异显著。
表4 毛白杨减数分裂四分体时期的小孢子群与天然2n花粉频率
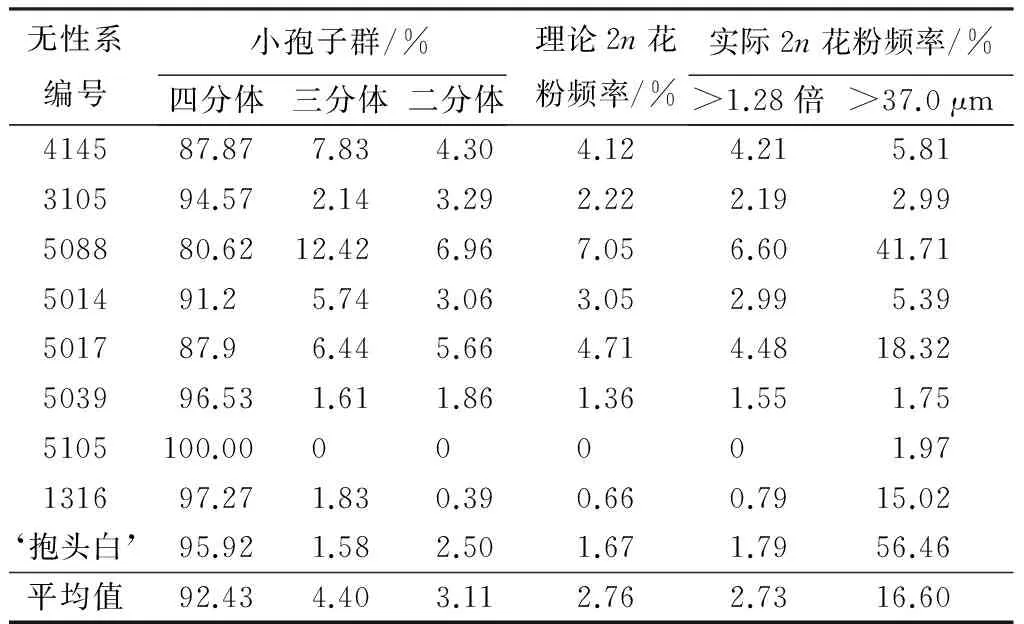
无性系编号小孢子群/%四分体三分体二分体理论2n花粉频率/%实际2n花粉频率/%>1.28倍>37.0μm414587.877.834.304.124.215.81310594.572.143.292.222.192.99508880.6212.426.967.056.6041.71501491.25.743.063.052.995.39501787.96.445.664.714.4818.32503996.531.611.861.361.551.755105100.0000001.97131697.271.830.390.660.7915.02‘抱头白’95.921.582.501.671.7956.46平均值92.434.403.112.762.7316.60
注:花粉粒平均直径大于1.28倍时χ2=0.10;花粉粒直径大于37.0 μm时χ2=2 322.60。
3 讨论与结论
天然2n花粉在植物界中普遍存在。由于2n花粉比正常单倍性花粉多一套染色体,因此判别2n花粉最为直接的方法就是进行花粉染色体计数。但因杨树花粉离体萌发率低的问题至今没有解决[15],致使通过花粉染色体计数来判别的2n花粉的发生难以实现。李凤兰等[16]借助荧光染料,对花粉细胞内的DNA进行染色,然后通过检测细胞内荧光的相对强弱来判别毛白杨2n花粉。证实了毛白杨大花粉的DNA含量约为单倍性花粉的2倍。
一般来说,随着花粉染色体数目的成倍增加,花粉粒直径也会随之增大,花粉粒大小与其倍性水平往往呈正相关[17]。因此,花粉粒的大小可作为判别植物多倍性花粉的重要形态学标志[18]。植物育种学家利用这一标志,已发现马铃薯[19]、秋海棠[20]、玫瑰[21]、毛白杨[9-10]、响叶杨[11]、通辽杨[12]以及香脂杨[13]等物种中广泛存在天然2n花粉。Orjeda等[19]研究发现,马铃薯2n花粉直径大约是单倍性花粉直径的1.30倍。朱之悌等[6]曾利用37 μm作为判别毛白杨2n花粉的阈值,花粉粒直径大于37 μm的花粉粒可视为2n花粉。
在本研究中,作者以胡杨和毛白杨不同无性系为材料,对不同无性系间花粉粒大小以及不同判别标准条件下的2n花粉发生频率进行了比较,发现胡杨花粉粒的平均直径为29.7 μm,比毛白杨花粉粒的平均直径32.4 μm略小,无性系对花粉粒直径影响显著。胡杨和毛白杨各无性系2n花粉的理论频率与以2n花粉粒直径须大于花粉粒平均直径的1.28倍为标准时统计的2n花粉实际频率差异不显著,而与以2n花粉粒直径大于37.0 μm为标准时统计的2n花粉实际频率差异极显著。这说明以37.0 μm作为判别胡杨或毛白杨2n花粉形态判别标准会导致较大的误差。而以各无性系花粉粒平均直径的1.28倍作为2n花粉形态判别标准更加科学合理。
[1] Nilsson Ehle H. Note regarding the gigas form of Populus tremula found in nature[J]. Hereditas,1936,21:372-382.
[2] Tamm Yu A, Yarvekyulg L. Results of searches for triploid aspen in Estonia[J]. Lesovedenid,1975,6:19-26.
[3] Every A D, Wiens D. Triploidy in Utah aspen[J]. Madrono,1971,21(3):138-147.
[4] Dillewijn C Van. Cytology and breeding ofPopulus[J]. Ned Boschb Tijdshr,1939,12:470-481.
[5] Gurreiro M G. The silvicultural improvement ofPopulus[M]. Publ Serv Flor Aquic Portugal,1944,11(1/2):53-117.
[6] 朱之悌,康向阳,张志毅.毛白杨天然三倍体选育研究[J].林业科学,1998,34(4):22-31.
[7] 张守攻,陈成彬,韩素英.中国部分杨属植物的染色体数目[J].植物分类学报,2005,43(6):539-544.
[8] Zhang S G, Qi L W, Han S Y. A report of triploid Populus of the section Aigeiros[J]. Silvae Genetica,2004,53(2):69-75.
[9] 朱之悌,林惠斌,康向阳.毛白杨异源三倍体B301等无性系选育的研究[J].林业科学,1995,31(6):499- 505.
[10] 康向阳.毛白杨2n花粉的发生机制[J].北京林业大学学报,2002,24(5):67-70.
[11] 鲁敏.响叶杨三倍体和四倍体诱导技术研究[D].北京:北京林业大学,2013.
[12] 王君,康向阳,李代丽,等.通辽杨花粉母细胞减数分裂及其染色体行为研究[J].西北植物学报,2006,26(11):2231-2238.
[13] Manzos A M. Fastgrowing form of Populus balsamifera obtaied by polliating female flowers with fractionated pollen of the same species[J]. Dokl Akad Nauk S SSR,1960,130(2):433-435.
[14] Seitz F W. The occurrence of triploids after self-pollination of anomalous androgynous flowers of a gray poplar[J]. Z Forstgenet,1954,3(1):1-6.
[15] 康向阳.毛白杨细胞遗传与三倍体选育[M].北京:中国环境科学出版社,2002:92-97.
[16] 李凤兰,张志毅.白杨染色体加倍技术研究及三倍体育种(Ⅱ):未减数花粉DNA的细胞荧光测定[J].北京林业大学学报,1992,14(增3):59-64.
[17] Jacob Y, Pierret V. Pollen size and ploidy level in the genus rosa[J]. Acta Hortic,2000,508:289-292.
[18] Veilleux R, Mchale N, Lauer F. 2n gametes in diploid Solanum: frequency and type of spindle abnormalities[J]. Can J Genet Cytol,1982,24:301-314.
[19] Orjeda G, Freyre R, Iwanaga M. Production of 2n pollen in diploid Ipomoea trifida, a putative wild ancestor of sweet potato[J]. J Hered,1990,81:462-467.
[20] Dewitte A, Eeckhaut T, Van Huylenbroeck J, et al. Meiotic aberrations during 2n pollen formation in Begonia[J]. Heredity,2010,104:215-223.
[21] Crespel L, Ricci S C, Gudin S. The production of 2n pollen in rose[J]. Euphytica,2006,151:155-164.
Discrimination Methods in 2nPollen inPopulus
Yang Qian, Chen Xin(Beijing Forestry University, Beijing 100083, P. R. China); Zhang Pingdong, Kang Xiangyang(National Engineering Laboratory for Tree Breeding, Beijing Forestry University)/Journal of Northeast Forestry University,2015,43(2):33-35,46.
Populuseuphratica;Populustomentosa; 2npollen; Discrimination in shape; Diameter of pollen grain
1) 国家自然科学基金项目(31370659)。
杨倩,女,1992年8月生,北京林业大学林学院,在读本科生。 E-mail:caoye11yq@163.com。
张平冬,林木育种国家工程实验室(北京林业大学),副教授。E-mail:zhangpd@bjfu.edu.cn。
2014年5月15日。
S791.223
责任编辑:潘 华。
