高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响
2015-03-10庞晓攀贾婷婷李倩倩雒眀伟郭正刚
庞晓攀, 贾婷婷, 李倩倩, 雒眀伟, 肖 玉, 赵 旭, 郭正刚
兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室, 兰州 730020
高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响
庞晓攀, 贾婷婷, 李倩倩, 雒眀伟, 肖 玉, 赵 旭, 郭正刚*
兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室, 兰州 730020
高原鼠兔(Ochotonacurzoniae)有效洞穴密度变化会引发高山嵩草(Kobresiapygmaea)草甸植物群落及其种群分布格局发生变化。采用野外调查法研究了高原鼠兔有效洞穴密度对高山嵩草群落特征及其主要种群分布格局的影响。结果表明:随高原鼠兔有效洞穴密度增加,高山嵩草草甸植物群落的优势种没发生明显变化,部分伴生种发生更替;高度、盖度、多样性指数和均匀度指数呈现降低态势;地上生物量和丰富度指数变化不明显;高山嵩草和矮火绒草(Leontopodiumnanum)种群的盖度、密度以及生物量均呈现降低态势,而达乌里秦艽(Gentianadahurica)和小花草玉梅(Anemonerivularisvar.flore-minors)种群的盖度、密度和生物量呈增加趋势。高山嵩草和矮火绒草的种群分布格局从8个/625m2和14个/625m2的聚集分布分别变为34个/625m2时的均匀分布和随机分布,达乌里秦艽和小花草玉梅种群从8个/625m2和14个/625m2的随机分布变为23个/625m2和34个/625m2时的聚集分布,这说明高原鼠兔有效洞穴密度变化改变了高山嵩草群落的特征和主要植物种群的空间分布格局,而对应群落特征和种群分布格局改变的有效洞穴密度为14个/625m2和23个/625m2。
植物群落组分; 植物多样性; 生物量; 密度; 空间分布格局
高原鼠兔(Ochotonacurzoniae)是青藏高原高寒草甸生态系统的关键种[1],当其密度适宜时,能优化高寒草甸植物群落的组成,提高植物的物种多样性[2],增加可食牧草的初级生产力[3],拓展种子植物的分布范围[4],增强土壤的水源涵养能力[5]。而当其密度超过高寒草甸承载力范畴时,便会致灾,威胁高寒草甸生态系统的健康,表现为土壤N和P含量配置比例失调[6],植物群落内不可食牧草急剧增加[7],原有优势植物高山嵩草种群的生态位明显衰减[8]。这说明高原鼠兔密度变化必然引起高寒草甸植物群落通过组分变化做出适应性应答,而这种适应性应答主要通过群落内主要植物种群特征及其分布格局的变化而实现的。植物种群特征主要包括某个植物种群的盖度、高度、密度和生物量。这些可量化指标往往反映一个植物种群在群落内地位的变化过程,以及该植物种群与环境的相互适应性。随着植物种群与环境的相互反馈适应过程的进行,植物种群的分布格局也将随之调整,而植物种群分布格局变化不仅受自身生物学特性的约束(如植物的繁殖方式)[9],而且受生存环境变化的影响[10],更受植物群落内不同种群间竞争过程的限制[11]。因此剖析植物群落内主要种群特征及其分布格局的变化特征,是解析植物群落适应因高原鼠兔密度变化而引起变化后环境的主要途径之一[12]。
高原鼠兔具有选择性采食、营家族式生活和运动迁移的特征,且通过地下洞穴和地面日夜运动,直接观测其单位面积内的密度数量具有不确定性[13],因此目前常用高原鼠兔有效洞穴密度替代高原鼠兔的方法研究高原鼠兔密度变化对高寒草甸群落特征的影响[3,8]。目前已经证实高原鼠兔有效洞穴密度的变化能够影响植物群落的地下和地上生物量、植物物种多样性,以及生草层厚度等数量和表观特征[7,14],事实上高寒草甸群落对高原鼠兔有效洞穴密度的响应是通过植物群落内部主要种群的响应而实现的,但目前高寒草甸群落内主要植物种群对高原鼠兔有效洞穴密度响应过程并不十分清楚。因此研究高原鼠兔有效洞穴密度变化对高寒草甸群落内主要植物种群特征及分布格局的影响,有利于揭示高原鼠兔有效洞穴密度变化改变植物群落数量和表观特征的机制。
高山嵩草(Kobresiapygmaea)草甸是青藏高原高寒草甸的主要地带性植被之一,分布于海拔3800—4500m的地带,是高山和高寒气候综合作用的产物,也是夏秋季家畜的主要放牧地[15]。由于高山嵩草草甸的优势种高山嵩草高度较低,往往是高原鼠兔栖息的重要植被类型之一[16],也是高原鼠兔容易致灾的主要高寒草甸类型[17]。因此研究高山嵩草群落及其种群分布格局对高原鼠兔有效洞穴的响应,是确定洞穴密度引起高山嵩草植物群落变化阈值的基础。本研究通过分析高原鼠兔有效洞穴密度对高山嵩草和主要伴生种的种群特征及其分布格局的影响,以期为揭示高山嵩草植物群落应答高原鼠兔有效洞穴密度变化的机制提供科学依据。
1 研究地区概况与方法
1.1 研究地区概况
研究地区位于青藏高原东部,行政隶属于甘肃省甘南自治州玛曲县,地理坐标为33°06′30″—34°30′15″ N,100°40′45″—102°29′00″ E,海拔3300—4806 m。气候为高寒湿润性气候,年均温度1.2 ℃,1月平均温度-10 ℃,7月平均温度11.7 ℃。年降水量约为564 mm,主要集中于5—9月,年蒸发量1000—1500 mm。全年日照时数约2613.9 h,年内霜期大于270 d,无绝对无霜期。土壤为亚高山草甸土,表层有5—10 cm草皮层,有机质含量可达10%—15%。植被类型主要是高寒草甸,以莎草科嵩草属的高山嵩草为优势种,伴生种有小花草玉梅(Anemonerivularisvar.flore-minors)、矮火绒草(Leontopodiumnanum)、达乌里秦艽 (Gentianadahurica)、莓叶萎陵菜(Potentillafragarioides)等。全县受鼠害危害的草地面积占总草地面积的比例30%左右,其中高原鼠兔是主要的危害鼠类,自20世纪70年代以来,高原鼠兔危害面积逐渐加大,单位面积有效洞穴数量逐渐增加,已经严重威胁高寒草甸生态系统的健康。
1.2 试验设计和样方调查
由于啮齿动物的有效洞穴密度与动物种群密度具有很好的相关性[3,7- 8,13,18],本研究亦采用有效鼠洞口数代替高原鼠兔密度的方法研究高原鼠兔密度变化对高山嵩草群落特征及其分布格局的影响。2012年5月在玛曲县阿孜站地势相对平坦的地块,随机选取了36个25 m×25 m的样地[3,6,8],用连续3d堵洞法测定各样地的有效洞口数,然后调查每个样地的裸斑面积和草丛高度,有效洞穴密度从5个到52个/625m2。为确定有效洞穴密度梯度,以有效洞穴、裸斑面积和草丛高度为变量,将其聚成4个类群。以每个类群的平均值为基准确定高原鼠兔有效洞穴梯度,其分别为10(10±3)、15(15±2)、21(21±3)、31(31±4)[6,8],每个梯度选择3个样地作为重复,共计12个样地,保证这12个样地在同一草场内,然后对这12个样地标记,并且围栏,避免放牧对高原鼠兔作用的影响。2012年11月到2013年2月份放牧,2013年3月至2013年10月继续围栏。2013年8月再次查数每个样地内的有效洞穴密度数量,虽然有效洞穴密度较2012年有稍微的变化,但其梯度效应仍然明显,分别为8(8±2)、14(14±2)、23(23±4)、34(34±5)。每个样地内采用“W”型布设5个1m×1m的样方,样方间隔约8 m,做样方调查。
统计样方内出现的所有植物种类,测定群落的总盖度和高度,其中盖度采用针刺法测定,植物高度测定采用直尺随机测定100次物种的株高,取其平均值。然后分种测定植物高度、盖度、地上生物量,分种盖度仍然采用针刺法,高度采用测定目标物种的平均值(随机测定30株,若不足20株,则全部测定)。地上生物量采用分种收获法测定,将植物其地面剪下带回实验室在(80℃)鼓风干燥箱内恒温烘干24 h,称其干重[3]。群落总生物量采用分种生物量总和计算。种群密度测度因植物不同采用不同的方法测定,双子叶植物以样方内出现的某种植物的总株数计算,该植物是否计入种群密度主要以其根茎是否在样方内为准;单子叶植物以样方内某种植物的枝条数计算,枝条数的统计主要以一级分枝为准[12]
1.3 植物群落数量特征的计算
物种重要值计算:
IV=(RHI + RCO + RBI)/3
式中,VI为重要值,RHI 为相对高度,RCO 为相对盖度,RBI 为相对生物量[3]。
物种多样性采用丰富度指数,均匀度指数和多样性指数测度,其中多样性指数采用Shannon-Wiener 指数(H′):
式中,H′为多样性指数,Pi为种的相对重要值。丰富度指数采用Patrick指数,即样方内物种总数S(每个鼠洞密度下各样方内物种的平均数)。均匀度指数采用Pielou指数:
P=H′/lnS
式中,P为均匀度指数,H′为多样性指数,S为样方内物种总数[19]。
1.4 植物种群空间分布格局的计算
种群空间分布格局的评价方法较多,但方差均值比率法和聚集强度指数法结合运用,可较高的分析处于临界状态时的植物种群空间分布格局[20],因此本研究采用这两种方法结合,测定植物群落内主要植物种群的空间分布格局。
方差均值比率法(V/m)中V和m的计算公式分别为
式中,N为样方数,Xi为每样方中的个体数。若V/m=1,则种群为均匀分布型;若V/m>1,则种群为聚集分布;如V/m<1,则种群呈均匀分布。然后采用t检验判断置信度,t值计算公式为
t=(V/m-1)/[2/(N-1)]1/2
式中,N为样方数,N-1为自由度,然后通过N-1自由度和95%的置信度t分布表,进行显著性检验。
聚集强度指数法分别采用丛生指数(I)、负二项参数(K)、聚块性指标(m*/m)和Cassie指标(Ca)测度:
I=(V/m-1)
式中,V为样本方差,m为样本均值。当I=0时,种群为随机分布;当I>0时,种群为聚集分布;当I<0时,种群为均匀分布。
K=m2/(V-m)
式中,V为样本方差,m为样本均值。当K>0时,种群呈聚集分布;当K趋于无穷大(一般为8以上),种群呈随机分布;当K<0时,种群呈均匀分布。
聚块性指标m*/m,当m*/m>1时,种群为聚集分布;当m*/m=1时,种群为随机分布;当m*/m<1时,种群为均匀分布,其中
m*=m+(V/m-1)
式中,V为样本方差,m为样本均值。
Ca=1/K,K为负二项分布的参数;当Ca=0时,种群为随机分布;当Ca>0时,种群为聚集分布;Ca<0时,种群为均匀分布。
1.5 数据分析
植物群落特征和种群主要特征分析先进行数据的方差齐次性检验,如果满足方差齐性(P>0.05),然后进行单因素方差分析,若差异显著(P<0.05),再应用Duncan法对平均值进行多重检验;如果数据不具方差齐次性,则用非参数法进行显著性分析,上述分析过程的软件采用SPSS 17.0统计软件包中的One-Way ANOVA。主要种群分布格局采用方差比率法,利用独立样本t检验进行显著性检验。
2 结果
2.1 高原鼠兔有效洞穴密度对高山嵩草群落组分和初级生产力的影响
高原鼠兔有效洞穴密度变化对高山嵩草群落特征具有明显的影响,当其从8个/625m2增至34个/625m2时,高寒草甸植物群落的优势种没有明显变化,为高山嵩草,主要伴生种发生了不同程度的变化,部分伴生种出现在4种有效鼠洞密度梯度下,如矮火绒草、小花草玉梅和达乌里秦艽,而部分伴生种却发生了一定的更替(表1),如有效洞穴密度为8个/625m2时的青藏苔草(Carexmoorcroftii)和大耳叶风毛菊(Saussureamacrota),而有效洞穴密度在34个/625m2时演变为垂穗披碱草(Elymusnutans)和鹅绒委陵菜(Potentillaanserine)。高山嵩草重要值虽然随高原鼠兔有效洞穴密度增加而逐渐降低,但仍然在各个有效洞穴密度下最大,说明高山嵩草在群落内的地位和作用虽然逐渐减弱,但仍然居于主导地位。伴生种达乌里秦艽和小花草玉梅重要值却随高原鼠兔有效洞穴密度增加而逐渐增加,说明其在群落内的地位和作用逐渐加强。
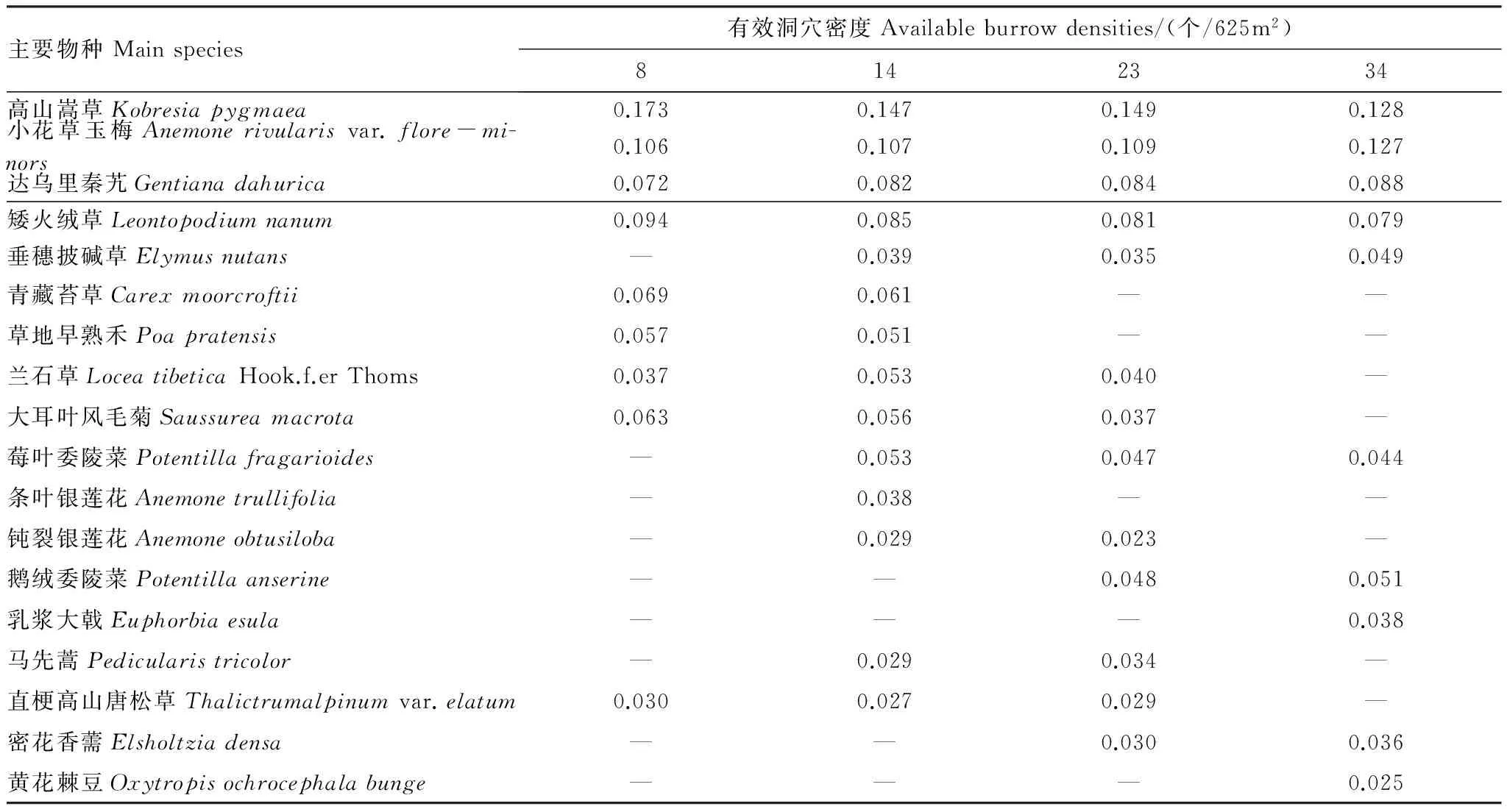
表1 不同高原鼠兔有效洞穴密度下高寒草甸植物群落的主要物种及其重要值Table 1 Species composition and their important value of alpine plant communities at different available burrow densities of plateau pika
“—”表示该植物种出现的样方数少于9个
不同高原鼠兔有效洞穴密度下植物群落高度、盖度和生物量均满足方差齐次性。单因素方差分析结果表明,高原鼠兔有效洞穴密度显著影响植物群落高度和盖度,而对生物量没有显著影响(表2)。多重比较结果表明,随着高原鼠兔有效洞穴密度增大,群落高度和盖度具有降低的态势(图1),表现为有效鼠洞密度为34个/625m2时群落高度和盖度显著低于鼠洞密度为8个/625m2时的群落高度和盖度(P<0.05)。
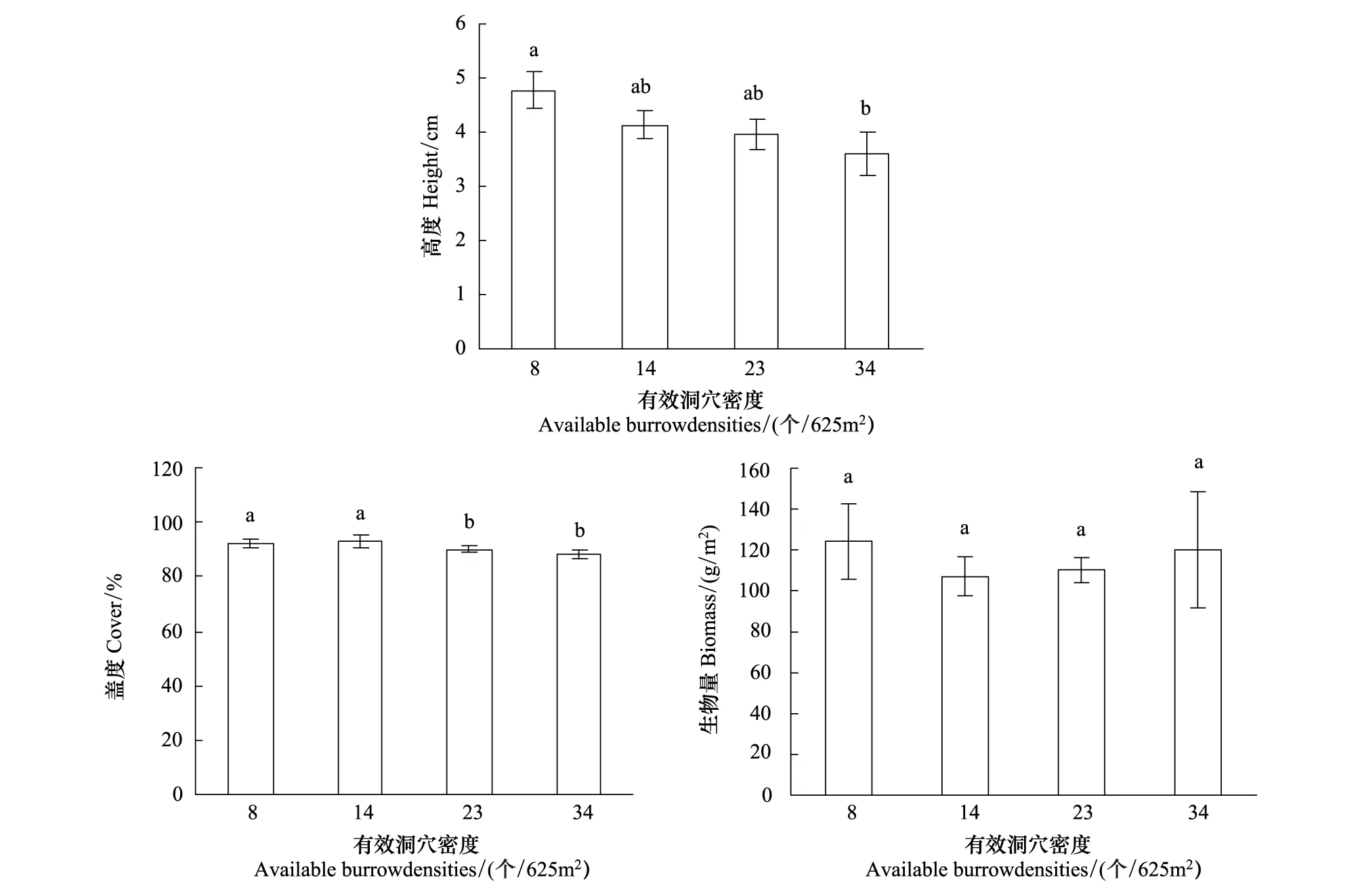
图1 高原鼠兔有效洞穴密度对高山嵩草群落高度、盖度及生物量的影响Fig.1 Effect of available burrow density of plateau pika on height, cover and biomass of K. capillifolia community不同小写字母表示差异显著(P<0.05)
表2 不同高原鼠兔有效洞穴密度下高寒草甸植物群落高度、盖度和生物量的方差分析结果
Table 2 One-way ANOVA result of height, cover and biomass of alpine plant community at different available burrow densities of plateau pika
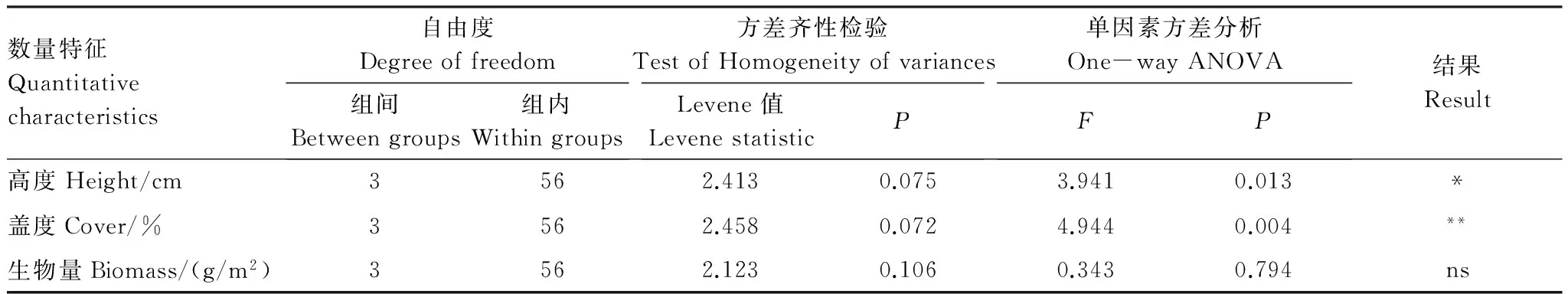
数量特征Quantitativecharacteristics自由度Degreeoffreedom组间Betweengroups组内Withingroups方差齐性检验TestofHomogeneityofvariancesLevene值LevenestatisticP单因素方差分析One-wayANOVAFP结果Result高度Height/cm3562.4130.0753.9410.013*盖度Cover/%3562.4580.0724.9440.004**生物量Biomass/(g/m2)3562.1230.1060.3430.794ns
**极显著水平; ns表示没有显著性
2.2 高原鼠兔有效洞穴密度对高山嵩草群落物种多样性的影响
不同高原鼠兔有效洞穴密度下植物丰富度指数、均匀度指数和多样性指数均满足方差齐次性。单因素方差分析结果表明,高原鼠兔有效洞穴密度显著影响均匀度和多样性指数,而对丰富度指数没有显著影响(表3)。多重比较结果表明,随着高原鼠兔有效洞穴密度增加,均匀度和多样性指数具有降低的变化态势(图2),有效洞穴密度为14个/625m2时的均匀度和多样性指数显著大于有效洞穴密度为34个/625m2时的均匀度指数和多样性指数(P<0.05)。
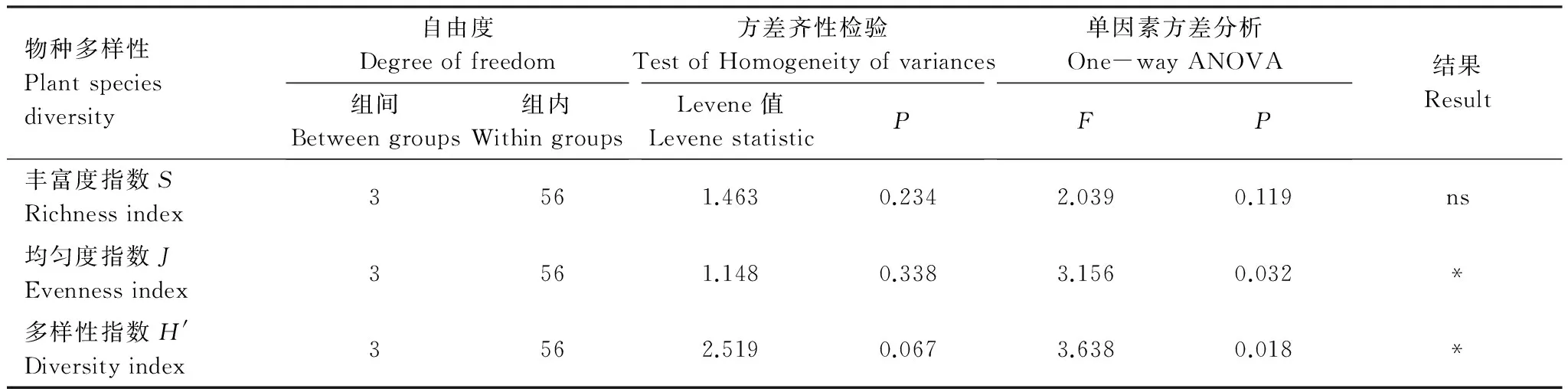
表3 不同高原鼠兔有效洞穴密度下高寒草甸植物物种多样性的方差分析结果Table 3 One-way ANOVA result of species diversity of alpine plant community at different available burrow densities of plateau pika
*显著水平;**极显著水平;ns没有显著性
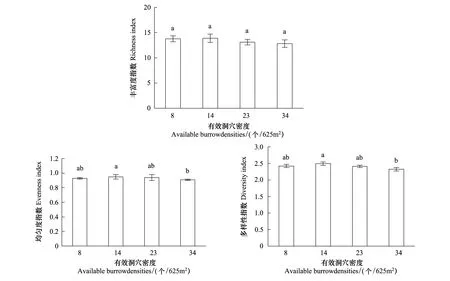
图2 高原鼠兔密度对草甸群落植物丰富度指数、均匀度指数和多样性指数的影响Fig.2 Effect of available burrow densities of plateau pika on the richness index, evenness index, and diversity index of alpine meadow community
2.3 高原鼠兔有效洞穴密度对高山嵩草群落主要种群密度和初级生产力的影响
不同高原鼠兔有效洞穴密度下高山嵩草、矮火绒草、达乌里秦艽以及小花草玉梅密度、盖度、高度以及生物量均满足方差齐次性。单因素方差分析结果表明,高原鼠兔有效洞穴密度对高山嵩草、矮火绒草、达乌里秦艽以及小花草玉梅的密度、盖度和生物量有显著影响,而对矮火绒草和小花草玉梅的高度无显著影响,但对高山嵩草和达乌里秦艽高度的影响具有显著性(表4)。多重比较结果表明,随高原鼠兔有效洞穴密度增加,高山嵩草和矮火绒草种群的盖度、密度和生物量均具有降低态势,当有效洞穴密度小于或等于14个/625 m2时,高山嵩草和矮火绒草种群的盖度、密度和生物量差异不显著(图3),而有效洞穴密度增至34个/ 625 m2时,高山嵩草和矮火绒草种群的盖度、密度、生物量均显著小于有效洞穴密度为8个/625 m2时的高度、盖度、密度和生物量 (P<0.05),但高山嵩草和矮火绒草种群的高度随着高原鼠兔有效洞穴密度增加的变化过程出现分异,表现为高原鼠兔有效洞穴密度增加具有降低高山嵩草种群高度的趋势,而对矮火绒草种群的高度没有显著影响。达乌里秦艽和小花草玉梅种群的密度、盖度和生物量随高原鼠兔有效洞穴密度增加呈增加的趋势,但两个植物种群响应有效洞穴密度变化的过程具有一定的分异性,其中达乌里秦艽和小花草玉梅种群的盖度和生物量在有效洞穴密度小于或等于23个/625 m2时差异不显著,而达乌里秦艽和小花草玉梅种群的密度分别在有效洞穴密度小于或等于23和14个/625m2时差异不显著,但达乌里秦艽和小花草玉梅的密度、盖度和生物量在34个/625 m2时显著大于8个/625m2时的密度、盖度和生物量。
表4 不同高原鼠兔有效洞穴密度下高山嵩草群落主要种群特征的方差分析结果
Table 4 One-way ANOVA result of main plant populations characteristics at different available burrow densities of plateau pika inK.capillifoliacommunity
数量特征Quantitativecharacteristics种群Plantpopulations自由度Degreeoffreedom组间Betweengroups组内Withingroups方差齐性检验TestofHomogeneityofvariancesLevene值LevenestatisticP单因素方差分析One-wayANOVAFP结果Result密度Density/(个/m2)高山嵩草Kobresiapygmaea3562.7300.05239.7880.000**矮火绒草Leontopodiumnanum3561.3920.25529.2450.000**达乌里秦艽Gentianadahurica3560.4840.6952.8850.044*小花草玉梅Anemonerivularisvar.flore-minors3560.9180.96357.8320.000**高度Height/cm高山嵩草Kobresiapygmaea3561.7450.16811.3490.015*矮火绒草Leontopodiumnanum3561.7730.1631.8690.145ns达乌里秦艽Gentianadahurica3561.2290.3085.0620.004**小花草玉梅Anemonerivularisvar.flore-minors3560.4490.7190.1300.942ns盖度Cover/%高山嵩草Kobresiapygmaea3562.2110.0973.7990.015*矮火绒草Leontopodiumnanum3562.4470.0736.2590.001**达乌里秦艽Gentianadahurica3562.0650.1158.9690.000**小花草玉梅Anemonerivularisvar.flore-minors3562.1210.1073.8320.013*生物量Biomass/(g/m2)高山嵩草Kobresiapygmaea3560.9320.4313.9950.012*矮火绒草Leontopodiumnanum3562.5210.0679.7740.000**达乌里秦艽Gentianadahurica3562.0510.1173.9590.012*小花草玉梅Anemonerivularisvar.flore-minors3562.0120.1043.6960.016*
*表示显著水平;**表示极显著水平;ns表示没有显著性
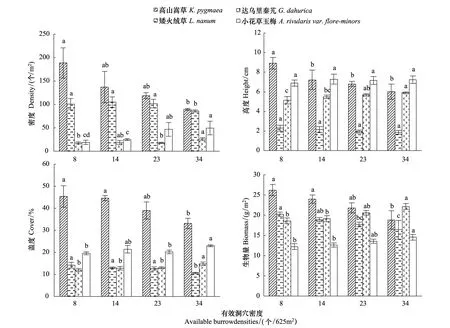
图3 高原鼠兔有效洞穴密度对高山嵩草群落主要种群密度、高度、盖度及生物量的影响Fig.3 Effect of available burrow densities of plateau pika on density, height, cover and biomass of main plant populations in K. capillifolia community
2.4 高原鼠兔有效洞穴密度对高山嵩草群落主要种群分布格局的影响
方差均值比率法和聚集指数法分析结果表明,高原鼠兔有效洞穴密度对高山嵩草群落主要种群的分布格局具有明显的影响。随着高原鼠兔有效洞穴密度的增加,高山嵩草种群的分布格局在8和14个/625m2时为聚集分布,到23个/625m2时变为随机分布,至34个/625m2时则为均匀分布(表5,表6)。矮火绒草种群的分布格局在有效洞穴密度小于或等于23个/625m2时为聚集分布,而有效洞穴密度为34个/625m2时则转变为随机分布。达乌里秦艽和小花草玉梅种群的分布格局在有效洞穴密度小于或等于14个/625m2时均为随机分布,而当有效洞穴密度高于或等于23个/625m2时,则变为聚集分布。
表5 不同有效洞穴密度下高山嵩草群落主要种群的方差均值比率法分布格局的分析
Table 5 Spatial distribution patterns of main plant populations inK.capillifoliacommunity assessed by Variance/Mean value at different available burrow densities of plateau pika

种群Plantpopulations有效洞穴密度ABD/(个/625m2)mVV/mt结果Result高山嵩草8188.607449.8039.5054.45**聚集分布Kobresiapygmaea14137.005617.0041.0056.57**聚集分布23118.60128.201.080.11*随机分布3489.4020.370.23-1.09*均匀分布矮火绒草8100.602555.3025.4034.51**聚集分布Leontopodiumnanum14105.004007.5038.1752.56**聚集分布23101.603199.5631.4943.12**聚集分布3486.0089.201.040.05*随机分布达乌里秦艽87.607.701.010.02*随机分布Gentianadahurica148.208.701.060.09*随机分布237.8023.202.972.79*聚集分布348.8074.708.4910.59**聚集分布小花草玉梅819.0020.501.0790.11*随机分布Anemonerivularisvar.1424.8025.701.040.05*随机分布flore-minors2346.781308.6927.9838.15**聚集分布3449.501774.9435.8649.30**聚集分布
ABD: available burrow densities;m: 样本均值sample mean;V: 样本方差sample variance;V/m: 样本方差比样本均值;*: 显著水平;**: 极显著水平
表6 不同有效洞穴密度下高山嵩草群落主要种群聚集指数分布格局的分析
Table 6 Spatial distribution patterns of main plant populations inK.capillifoliacommunity assessed by aggregation index under different available burrow densities of plateau pika
种群Plantpopulations有效洞穴密度ABD/(个/625m2)IKm*/mCa结果Result高山嵩草838.504.901.200.20聚集分布Kobresiapygmaea1440.003.431.290.29聚集分布230.081465.971.000.00随机分布34-0.77-115.780.99-0.01均匀分布矮火绒824.404.121.240.24聚集分布Leontopodiumnanum1437.172.831.350.35聚集分布2330.493.331.300.30聚集分布340.042311.251.000.00随机分布达乌里秦艽80.01577.601.000.00随机分布Gentianadahurica140.06134.481.000.01随机分布231.973.951.250.25聚集分布347.491.181.850.85聚集分布小花草玉梅80.08240.671.000.00随机分布Anemonerivularisvar.140.04683.381.000.00随机分布flore-minors2326.981.731.580.58聚集分布3434.861.421.700.70聚集分布
ABD: available burrow densities;I: 丛生指标Clumping index;K: 负二项参数Negative binomial parameter;m*/m: 聚块性指标Patchiness index;Ca:Cassie指标Cassie index
3 讨论
高原鼠兔有效洞穴密度增加不仅意味着单位面积高山嵩草草甸内高原鼠兔选择性采食的行为有所加强,而且意味着高山嵩草草甸上裸斑面积增加,这种变化一方面增加了高山嵩草群落内高原鼠兔喜食植物生长受抑制的程度[21],释放了高原鼠兔不喜食植物生长的潜势[22],一方面大幅改变了高山嵩草草甸原有吸收和反射太阳辐射的过程,导致土壤水分含量变化速率加快,改变了原有嵩草草甸内植物组分的生境[17,23],迫使高山嵩草草甸植物群落组分发生应答。本研究结果表明,高原鼠兔有效洞穴密度从8个/625m2增加至34个/625m2的过程中,湿中生的青藏苔草和大耳叶风毛菊[24- 25]退出群落,而旱中生的垂穗披碱草和鹅绒委陵菜则演变为主要伴生种[26- 27],说明高山嵩草群落生境旱生化后已经不适宜湿生性植物的茁壮生长,而有利于中旱生植物被抑制生长潜势的释放。高寒草甸群落伴生种的更替势必影响其高度和盖度,当高原鼠兔有效洞穴密度等于或者大于23个/625m2,高寒草甸群落盖度显著降低,主要是因掘土活动而产生的裸斑数量逐渐增多[13],但给中旱生型达乌里秦艽和莓叶萎陵菜等植物的生长创造了良好的条件[8],然而这些旱生物种高度较低,导致群落高度有所降低。然而地上生物量对高原鼠兔有效洞穴密度响应不明显,这与高原鼠兔对青海果洛地区高寒草甸群落生物量的影响相一致[3],主要原因是高原鼠兔有效洞穴密度较低的时候,以无性繁殖为主的中生植物高山嵩草和湿中生的矮火绒草占据主导地位,随高原鼠兔有效洞穴密度增加,群落内湿中生和以无性繁殖为主的植物比例有所降低,但达乌里秦艽和小花草玉梅等中旱生植物比例明显增加,且达乌里秦艽和小花草玉梅等植物叶形较宽,从而弥补了湿生植物比例降低所减少的生物量。因此,高山嵩草群落随着高原鼠兔有效洞穴密度增加时虽然高度和盖度有所降低,但在物种更替过程中,株型较高高山嵩草和盖度的矮火绒草比例下降,而达乌里秦艽和小花草玉梅等宽叶形植物比例有所增加,使群落地上生物量维持在一个相对稳定的水平。
高山嵩草群落内伴生种随高原鼠兔有效洞穴密度增加而发生一定程度更替,不仅影响群落盖度和高度,而且影响植物多样性的变化[28]。虽然高原鼠兔有效洞穴密度对高山嵩草群落的物种丰富度没有明显的影响,但高原鼠兔有效洞穴密度为14个/625m2时均匀度指数和多样性指数最大,说明当高原鼠兔有效洞穴密度从8个/625m2增加至34个/625m2的过程中,虽然样方内出现的物种有所变化,但通过湿中生物种的退出和中旱生物种的入侵,其物种总数变化不是非常明显,然而群落内不同植物种群的分布均匀性则发生了明显的改变,这主要是不同植物种群适宜生境存在差异所致[2,8]。高原鼠兔有效洞穴密度小于或等于23个/625m2时,裸斑数量相对较小,为湿中生和中旱生植物提供了共存的生境,物种多样性变化不明显,但当高原鼠兔有效洞穴密度超过23个/625m2时,一些湿生或阴生的植物退出群落,而中旱生植物生长能力逐渐加强,植物群落内种群分布的均匀性有所降低,从而一定程度上降低了物种多样性。
高原鼠兔有效洞穴密度增加引起高山嵩草群落组分和特征的变化,其主要通过高山嵩草群落内不同植物种群对微环境变化的适应程度而实现[22,29]。当高原鼠兔有效洞穴密度小于或等于23 个/625m2时,高山嵩草和矮火绒草种群的数量指标值变化不明显,但高原鼠兔有效洞穴密度超过23个/625m2时,其种群数量指标值明显减少,这是因为高原鼠兔有效洞穴密度增加与土壤地表蒸发增加和植物根系层土壤水分含量减少具有一致性[29],当高原鼠兔有效洞穴密度小于23个/625m2时,生境内土壤水分等资源供给能力虽然逐渐减弱,但还没有成为高山嵩草和矮火绒草的限制因子,而当高原鼠兔有效洞穴密度超过23个/625m2时,生境内的土壤水分等资源的供给能力可能已经演变为高山嵩草和矮火绒草的限制因子。中旱生的达乌里秦艽和小花草玉梅种群的数量指标值明显变化发生的高原鼠兔有效洞穴密度介于14个/625m2到23个/625m2,而小于14个/625m2或大于23个/625m2时则无明显变化,说明随着生境旱化,达乌里秦艽和小花草玉梅的适宜性增强。因此不同植物种群数量特征的变化对应最敏感的高原鼠兔有效洞穴密度是14个/625m2至23个/625m2。
当高原鼠兔有效洞穴密度小于14个/625m2时,土壤水分相对充沛,生境湿润,对喜生于湿润环境的高山嵩草和矮火绒草而言[30- 31],表现为资源充沛且呈斑块状镶嵌分布,其具有较强的资源竞争和利用能力[8],种群空间分布格局为聚集分布,而对于旱、中生宽叶达乌里秦艽和小花草玉梅而言[32],此时他们的资源供给相对不足,其空间分布格局则呈为随机分布。随着高原鼠兔有效洞穴密度的增加,土壤环境逐渐旱化,湿中生的高山嵩草和矮火绒草在群落内的竞争能力逐渐下降,竞争性利用资源能力降低,资源供给逐渐不利于他们的生长,高山嵩草和矮火绒草种群的空间分布格局从聚集分布转变为随机分布,其中高山嵩草在34个/625m2时则变为均匀分布,其生长受环境限制趋强,而对达乌里秦艽和小花草玉梅而言,旱生化的环境却具有资源充沛,且斑块分布的特征,此时裸斑面积增加,给以种子繁殖为主的达乌里秦艽和小花草玉梅提供了很多的定居空间和栖息地,而高原鼠兔的啃食行为可能促进了达乌里秦艽和小花草玉梅种子的萌发,因此其空间分布格局演变聚集分布。
高山嵩草群落特征和主要种群分布格局对高原鼠兔有效洞穴密度的响应过程表明,当高原鼠兔有效洞穴密度大于23个/625m2时,高山嵩草草甸群落内原有优势植物种群的功能和地位具有弱化趋势,而阔叶型杂草种群的功能和地位具有强化态势,因此,将高原鼠兔有效洞穴密度维持在其合理的范围内,有利于高原鼠兔鼠害防控中趋利避害。
[1] Smith A T, Foggin J M. The plateau pika (Ochotonacurzoniae) is a keystone species for biodiversity on the Tibetan plateau. Animal Conservation, 1999, 2(4): 235- 240.
[2] Bagchi S, Namgail T, Ritchie M E. Small mammalian herbivores as mediators of plant community dynamics in the high-altitude arid rangelands of Trans-Himalaya. Biological Conservation, 2006, 127(4): 438- 442.
[3] Guo Z G, Li X F, Liu X Y, Zhou X R. Response of alpine meadow communities to burrow density changes of plateau pika (Ochotonacurzoniae) in the Qinghai-Tibet Plateau. Acta Ecologica Sinica, 2012, 32(1): 44- 49.
[4] Alcántara J M, Rey P J, Sánchez-Lafuente A M, Valera F. Early effects of rodent post-dispersal seed predation on the outcome of the plant-seed disperser interaction. Oikos, 2000, 88(2): 362- 370.
[5] 孙飞达, 郭正刚, 尚占环, 龙瑞军. 高原鼠兔洞穴密度对高寒草甸土壤理化性质的影响. 土壤学报, 2010, 47(2): 378- 383.
[6] 李倩倩, 赵旭, 郭正刚. 高原鼠兔有效洞穴密度对高寒草甸优势植物叶片和土壤氮磷化学计量特征的影响. 生态学报, 2014, 34(5): 1212- 1223.
[7] 孙飞达, 龙瑞军, 路承香. 高原鼠兔不同洞穴密度对高寒草地植物群落组成及多样性的影响. 干旱区资源与环境, 2010, 24(7): 181- 186.
[8] 贾婷婷, 毛亮, 郭正刚. 高原鼠兔有效洞穴密度对青藏高原高寒草甸群落植物生态位的影响. 生态学报, 2014, 34(4): 869- 877.
[9] Pottier J, Marrs R H, Bédécarrats A. Integrating ecological features of species in spatial pattern analysis of a plant community. Journal of Vegetation Science, 2007, 18(2): 223- 230.
[10] Deléglise C, Loucougaray G, Alard D. Spatial patterns of species and plant traits in response to 20 years of grazing exclusion in subalpine grassland communities. Journal of Vegetation Science, 2011, 22(3): 402- 413.
[11] 高福元, 赵成章, 卓马兰草. 高寒退化草地不同海拔梯度狼毒种群分布格局及空间关联性. 生态学报, 2014, 34(3): 605- 612.
[12] 晁增国, 汪诗平, 徐广平, 胡宜刚, 张振华. 围封对退化矮嵩草草甸群落结构和主要种群空间分布格局的影响. 西北植物学报, 2008, 28(11): 2320- 2326.
[13] 韩天虎, 花立民, 许国成. 高原鼠兔危害级别划分. 草业学报, 2008, 17(5): 130- 137.
[14] 石红霄, 于健龙. 高原鼠兔洞口密度对高寒嵩草草甸植被及土壤水分的影响. 中国草地学报, 2010, 32(4), 109- 112, 116- 116.
[15] 王启基, 李世雄, 王文颖, 景增春. 江河源区高山嵩草(Kobresiapygmaea)草甸植物和土壤碳、氮储量对覆被变化的响应. 生态学报, 2008, 28(3): 885- 894.
[16] 李雄, 胡向阳, 杨永平. 青藏高原重要牧草高山嵩草的研究现状. 草业与畜牧, 2013, (1): 30- 39.
[17] 周雪荣, 郭正刚, 郭兴华. 高原鼠兔和高原鼢鼠在高寒草甸中的作用. 草业科学, 2010, 27(5): 38- 44.
[18] Johnson W C, Collinge S K. Landscape effects on black-tailed prairie dog colonies. Biological Conservation, 2004, 115(3): 487- 497.
[19] 杜虎, 彭晚霞, 宋同清, 王克林, 曾馥平, 鹿士杨, 时伟伟, 唐成, 谭秋锦. 桂北喀斯特峰丛洼地植物群落特征及其与土壤的耦合关系. 植物生态学报, 2013, 37(3): 197- 208.
[20] 吴继林. 不同方法在珍稀植物长苞铁杉种群分布格局分析中的适用性研究. 江西农业大学学报, 2001, 23(3): 345- 349.
[21] Pech R P, Arthur A D, Zhang Y M, Lin H. Population dynamics and responses to management of plateau pikasOchotonacurzoniae. Journal of Applied Ecology, 2007, 44(3): 615- 624.
[22] Wesche K, Nadrowski K, Retzer V. Habitat engineering under dry conditions: the impact of pikas (Ochotonapallasi) on vegetation and site conditions in southern Mongolian steppes. Journal of Vegetation Science, 2007, 18(5): 665- 674.
[23] Davidson A D, Lightfoot D C. Burrowing rodents increase landscape heterogeneity in a desert grassland. Journal of Arid Environments, 2008, 72(7): 1133- 1145.
[24] 武高林, 陈敏, 杜国祯. 营养和光照对不同生态幅风毛菊属植物幼苗形态可塑性的影响. 应用生态学报, 2008, 19(8): 1708- 1713.
[25] 李中强, 张萌, 徐军. 青藏高原纳木错水生植物多样性及群落生态学研究. 水生生物学报, 2011, 35(1): 1- 13.
[26] 王桔红, 崔现亮, 陈学林, 杜国祯. 中、旱生植物萌发特性及其与种子大小关系的比较. 植物生态学报, 2007, 31(6): 1037- 1045.
[27] 张大勇, 王刚, 杜国祯. 甘南山地草原人工草场的演替. 植物生态学与地植物学学报, 1990, 14(2): 103- 109.
[28] 汪殿蓓, 暨淑仪, 陈飞鹏. 植物群落物种多样性研究综述. 生态学杂志, 2001, 20(4): 55- 60.
[29] Guo Z G, Zhou X R, Hou Y. Effect of available burrow densities of plateau pika (Ochotonacurzoniae) on soil physicochemical property of the bare land and vegetation land in the Qinghai-Tibetan Plateau. Acta Ecologica Sinica, 2012, 32(2): 104- 110.
[30] 黄瑞灵, 周华坤, 刘泽华, 徐维新, 魁武. 长江源区高山嵩草物候与生物量动态及其对气候变化的响应. 西北植物学报, 2012, 32(5): 1021- 1026.
[31] 王俊峰, 王根绪, 王一博, 李元寿. 青藏高原沼泽与高寒草甸草地退化对生长期CO2排放的影响. 科学通报, 2007, 52(13): 1554- 1560.
[32] 杨文利, 曹瑞, 马虹, 李莉. 小花草玉梅(Anemonerivularisvar.flore-minore)花粉形态的变异及其进化. 内蒙古大学学报: 自然科学版, 2002, 33(2): 191- 193.
Effect of available burrow densities of plateau pika (Ochotonacurzoniae) on characteristics and distribution pattern ofKobresiapygmaeacommunity
PANG Xiaopan, JIA Tingting, LI Qianqian, LUO Mingwei, XIAO Yu, ZHAO Xu, GUO Zhenggang*
StateKeyLaboratoryofGrasslandAgro-ecosystems,CollegeofPastoralAgricultureScienceandTechnology,LanzhouUniversity,Lanzhou730020,China
The increase in available burrow density of plateau pika (Ochotonacurzoniae) changes the plant community characteristics and distribution pattern of main plant population ofKobresiapygmaeameadow in the Qinghai-Tibet Plateau. A field survey was carried out to determine the effect of available burrow density of plateau pika on plant community characteristics and distribution pattern of main plant population ofK.pygmaeameadow. This study showed that the dominant plant ofK.pygmaeacommunity were similar, while some associate plants ofK.pygmaeacommunity were replaced. The plant height, plant cover, the evenness index and diversity index ofK.pygmaeacommunity decreased, while the richness index and aboveground biomass ofK.pygmaeacommunity kept stable as the available burrow density of plateau pika increased. The plant cover, density and aboveground biomass ofK.pygmaeaandLeontopodiumnanumpopulation decreased, while the plant cover, density and biomass ofGentianadahuricaandAnemonerivularisvar.flore-minorspopulation increased with the increase of available burrow density. The spatial distribution patterns ofK.pygmaeaandL.nanumchanged from aggregation distribution at 8 and 14 available burrows per 625m2to even distribution and random distribution at 34 available burrows per 625m2, respectively. The random distribution patterns ofG.dahuricaandA.rivularisvar.flore-minorspopulation at 8 and 14 available burrows per 625m2were replaced by aggregation distribution patterns at 23 and 34 available burrows per 625m2. These results suggest that the available burrow density changes the plant community characteristics and distribution pattern of main plant population ofKobresiapygmaeameadow at 14 and 23 available burrows per 625m2.
plant community composition; plant diversity; biomass; density; spatial distribution pattern
国家自然科学基金(31172258); 国家行业(农业)公益项目(201203041); 教育部“长江学者和创新团队发展计划资助”项目(IRT13019)
2014- 03- 09;
日期:2014- 09- 28
10.5846/stxb201403090400
*通讯作者Corresponding author.E-mail: guozhg@lzu.edu.cn
庞晓攀, 贾婷婷, 李倩倩, 雒眀伟, 肖玉, 赵旭, 郭正刚.高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响.生态学报,2015,35(3):873- 884.
Pang X P, Jia T T, Li Q Q, Luo M W, Xiao Y, Zhao X, Guo Z G.Effect of available burrow densities of plateau pika (Ochotonacurzoniae) on characteristics and distribution pattern ofKobresiapygmaeacommunity.Acta Ecologica Sinica,2015,35(3):873- 884.
