乌梁素海冰封期营养盐及浮游植物的分布特征
2014-10-22李卫平徐静于玲红李兴韩佩江赵忠井璟
李卫平,徐静 ,于玲红,李兴,韩佩江赵忠,井璟
1.内蒙古科技大学能源与环境学院,内蒙古 包头 014010;2.安徽马钢工程技术集团有限公司设计研究院,安徽 马鞍山 243000;3.内蒙古师范大学内蒙古节水农业工程研究中心,内蒙古 呼和浩特 010022;4.内蒙古巴彦淖尔市河套水务集团有限公司,内蒙古 巴彦淖尔 015000;5.内蒙古巴彦淖尔市环境监测站,内蒙古 巴彦淖尔 015000
浮游植物是水生态系统的初级生产者,也是整个水生态系统中物质循环和能量流动的基础,同时,还能够迅速响应水体的营养状态变化(韩博平等,2003)。有些浮游植物可以直接用作环境监测的指示生物,而且相对于理化条件而言,其密度、生物量、种类组成和多样性能更加准确的反映出水体的营养水平(刘红等,2006)。在湖泊环境中,营养盐是影响浮游植物生长的关键因素,同时又是导致水体富营养化的主要原因(李文霞和冯海艳,2006;周敏,2012)。以往对营养物质与浮游植物群落结构特征的研究都是在常温水体条件下进行的,而在冰封条件下,水体和冰体中的营养盐与浮游植物群落特征的研究尚不多见。因此,本实验以乌梁素海为研究对象,研究湖水结冰后,营养盐与浮游植物在冰体和水体中的分布情况,以期为该方向的研究提供理论基础。
1 研究区域及方法
1.1 区域概况与采样点布设
乌梁素海地处我国内蒙古自治区境内,位于我国北方干旱半干旱地区,属温带大陆性气候,光照辐射强、降水量少、蒸发强烈,昼夜温差大,年均降水量约为285 mm,蒸发量约为2456 mm;冬季寒冷干燥,夏季炎热少雨,全年平均气温约为9.74 ℃,每年的11月至翌年3月为冰封期。湖泊面积现为273 km2,其主要补给水源是河套灌区的农田退水,湖区的水动力条件较弱,最大流速为0.28 m·s-1,最小为 0。目前,每年随农田退水进入乌梁素海的总氮和总磷分别约为3411.58和111.88 t(吕超等,2012;李卫平等,2008;徐兆安等,2012),如此大量的营养物质输入,使得整个湖区呈Ⅴ类(参考GB 3838—2002地表水环境质量标准)水质,且呈现富营养化状态。根据乌梁素海的水域污染状态和进出水位置,本研究共布设了 10个采样点,见图1,其中J11为进水口、V3为出水口、L15和S6均为明水区域、N13为芦苇区、Q10为旅游区,这些采样点对于冰封期水体的研究具有一定的代表性。湖区北部为小海子,芦苇覆盖大量明水区域且常年不受人为活动影响,故而不做研究。

1.2 样品采集与处理
本研究于2012年11月至2013年2月在乌梁素海使用冰柱采集器破冰采样,将收集到的的冰样分为3层:表层冰、中层冰、冰—水界面冰,分别置于水桶内,及时送回实验室,在室温条件下自然融化,然后测定融水中的 TP、TN、COD和 Chla浓度。现场监测的指标为水深、冰厚和流速;同时,用取水瓶(聚乙烯材料)分别取冰—水界面水、中层水、水—底泥界面水3组水样,并于12 h内于实验室分析TP、TN、COD和Chla浓度,测定方法参照《水和废水监测分析方法》(国家环境保护总局,2002)。测试所用标准曲线相关性严格达标(r>0.999)所有监测指标均做3组平行,取平均值为测定结果。试验数据用Microsoft Excel 2003进行初步分析和作图。指标测定方法及采样时间分别见表1和表2。
浮游植物定量样品用采水器在现场取 1000 mL水面下0.5 m处的水样,加入15 mL鲁哥试液固定;静置沉淀24 h,用虹吸管抽掉上清液,余下20~25 mL沉淀物转入30 mL定量瓶中,并定容至30 mL。将在室温条件下融化的冰块作为冰体中的浮游植物定量样品,取1000 mL,处理步骤同上。镜检前将定容后的水样摇匀,取 0.1 mL于 20 mm×20 mm的浮游生物计数框内,置10×40倍显微镜下进行种类鉴定和细胞计数。每个采样点细胞计数3次,取平均值,浮游植物种类鉴定参考文献(周凤霞和陈剑虹,2011;马放等,2010)进行,由《湖泊富营养化调查规范》可查得藻类计数计算方法(金相灿和屠清瑛,1990)。

表1 实验监测项目及方法Table 1 The monitoring project and methods of test

表2 采样时间Table 2 Sampling times

式中:N为藻类密度,104cell·mL-1;A为浮游生物计数框面积,mm2;Ac为计数面积,即视野面积×视野数,mm2;Vw为l L水样经沉淀浓缩后的样品体积,mL;V为计数框体积,mL;n为计数所得藻类的个数。
运用浮游植物优势度Y来分析乌梁素海冰封期浮游植物群落结构特征,将优势度Y>0.02的藻类定为优势种。各参数计算公式如下:

式中:ni为第i种浮游植物的的数量;N为浮游植物总数量;fi为第i种浮游植物在所采样品中出现的频率。
2 结果与分析
2.1 营养盐的分布特征
2.1.1 冰体分析
现场监测结果显示,湖水自2012年11月初开始结冰,11月至12月为湖冰的快速生长期,湖冰厚度在翌年1月份达到最大,厚度在0.22~0.74 m之间,之后开始缓慢消融。其中,湖中心的冰体最厚,为0.74 m,湖区的南部和北部较薄。这是因为乌梁素海自北向南有总排干、通济渠等灌渠和排水沟与湖体相连,使得西部湖区水体的流动性较大,冰体厚度相对于东部湖区较小。
从时间分布上看:冰层中的营养盐含量整体呈下降的趋势。这是由于在结冰过程中,冰体会自动排除杂质,以保持其纯度(姜慧琴,2011;高红杰等,2008),被排除的营养盐由冰体逐渐迁移进入水体(图2)。
冰体中营养盐的垂向分布规律为:在冰体形成期,表层冰>冰水界面冰≥中层冰。这是由乌梁素海特殊的气候条件所致,昼夜温差大使得湖水在结冰初期处于反复的冻融过程,白天温度较高,冰体融化时会释放原本被冻结在体内的营养盐,由于湖区水动力条件较弱,被释放的营养盐大多停留在冰水界面层;夜晚温度降低时,营养盐被再次冻结,如此反复使得该时期的冰体表层营养盐含量较高(张岩,2012;傅鑫廷,2009)。冰层厚度稳定后,冰体中的营养盐自上而下逐渐迁移进入水体,导致中、下层冰体中的营养盐含量略大于表层(图3)。
2.1.2 水体分析


乌梁素海湖泊冰封期各采样点的水深均在0.28~2.45 m之间,其平均值为1.20 m,其中,湖泊的南部和北部水深较深,而其冰厚度值较小;湖泊的中部则与之相反,这表明水深和冰厚存在较好的负相关性。这可以从热力学角度和湖冰的形成机理上加以解释:在受冷面积一定的条件下,单位时间内作用于一定受冷面积上的冷能是相同的,而等量的冷能要通过冰体的冷传导作用到冰下水体中,从而增加冰层厚度。而较浅的水体在获得相同冷能时温度的降低更为明显,从而冰厚的增加较深水体要快。但是,在上层获取冷能的同时,下层水体也在获得大地所传导的等量热能,同理,较浅的水体在获得相同热能时温度也会明显升高,但在冰体的生长过程中,冷能的传导起主要作用,最终导致水深和冰厚存在较好的负相关性(姜慧琴,2011)。
从图4中不难发现,在湖水的结冰过程中,被排斥而无法进入冰体的营养盐大多活跃在冰水界面水中;水底泥界面水受到水与底泥之间的营养盐交换作用影响(Cho等,2002),故而水体中的营养盐含量垂向分布规律为:冰水界面水>水底泥界面水≥中层水。从时间分布上看:冰水界面水的营养盐含量无明显变化,中层和水底泥界面水营养盐含量呈上升趋势(图5)。
2.2 浮游植物结构及其分布特征


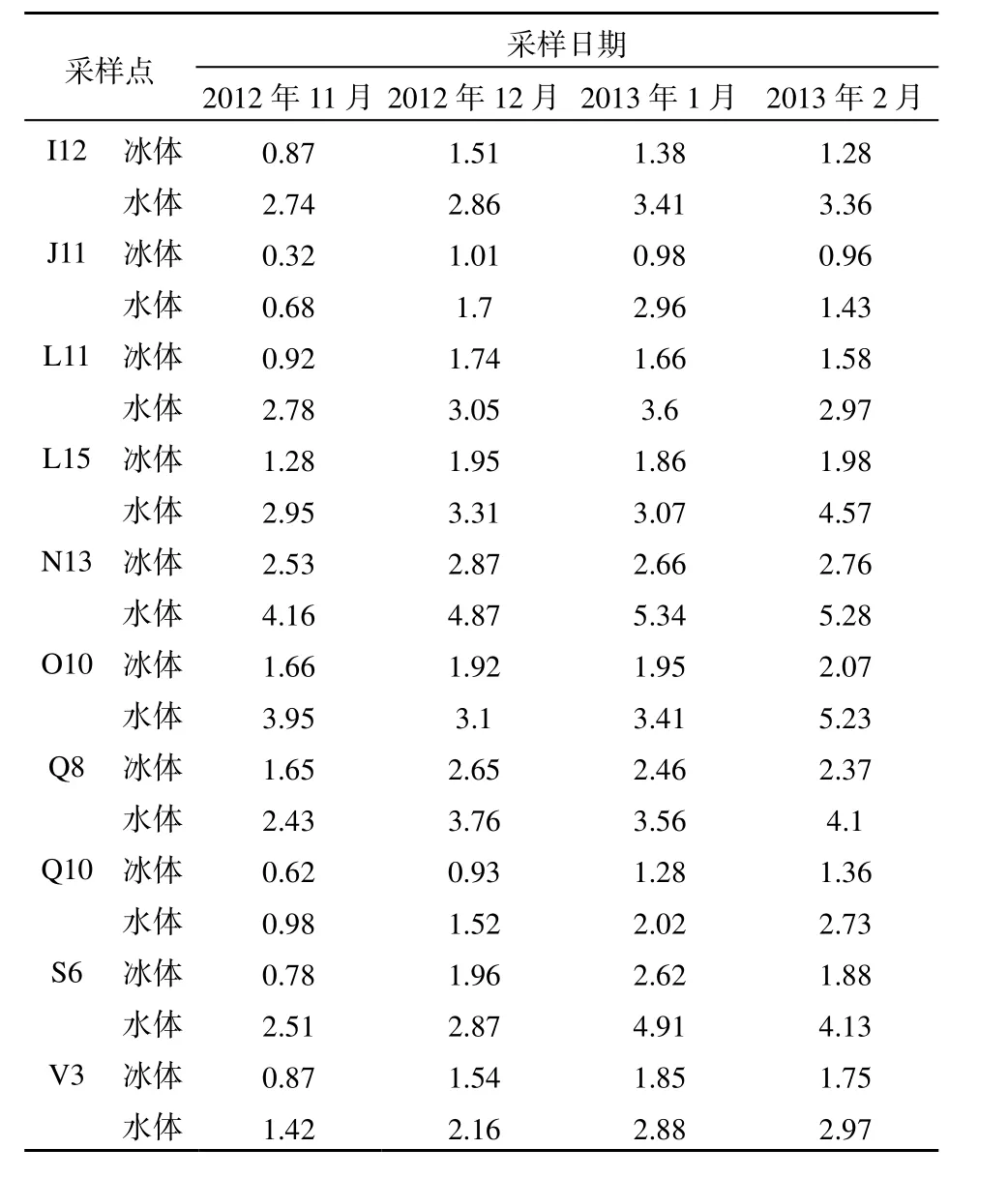
表3 乌梁素海冰体和水体中的藻类密度Table 3 The number of algae in Lake Wuliangsuhai water and ice 104个·mL-1
表3记录了乌梁素海冰封期10个采样点的藻类密度。在冰体的形成和生长时期,由于大气和水之间的热量传递作用,一部分藻类以漂浮的状态悬浮在水体表面,之后随着水体的结冰过程,漂浮在水体表面的藻类被冻结在冰体中,从而使得冰体中的藻类密度逐渐上升(Matsuo和Sasyo,1981;龚乃虎,1999;Knight,1979;徐立冲等,1997);之后随着冰层厚度的增加,开始形成较大体积而藻类密度较少的冰晶体,使得冰体中的藻类密度趋于稳定。水体中的藻类密度整体呈现缓慢上升趋势,一方面是由于冰封期水体的营养物质充足,藻类开始大量繁殖生长;另一方面又由于冬季水温较低,藻类的生长和繁殖受到了一定的限制,这也是冬季湖泊不宜暴发水华的原因之一。
此次调查共检出浮游植物7门67属105种(含变种),隶属于绿藻门、硅藻门、蓝藻门、裸藻门、金藻门、隐藻门以及甲藻门,未检测出黄藻门,这是因为黄藻门主要是淡水产浮游藻类,尤其喜欢在较清洁的水体中生长,而乌梁素海的现有水质达不到其生长繁殖要求。镜检结果显示,冰封期乌梁素海水体和冰体中的藻类均以绿藻门和硅藻门为主,其中绿藻门有23属35种,占总种数的33.3%;硅藻门 16属 28种,占总种数的 26.7%;蓝藻门 13属19种,占总种数的18.1%;裸藻门5属10种,占总种数的9.5%;甲藻门5属8种,占总种数的7.6%;金藻门3属3种,占总种数的2.9%;隐藻门2属2种,占总种数的1.9%。绿藻门和硅藻门在浮游植物群落结构组成中处于优势地位,表明冰封期乌梁素海浮游植物种类组成呈绿藻—硅藻型,各采样点浮游植物种类组成存在差异,但均以绿藻和硅藻种数最多,蓝藻次之,这是因为蓝藻喜高温,通常在 20 ℃以上的水体中大量生长繁殖,而冰封期水体温度在0.5~6 ℃之间波动,该温度不适宜蓝藻生长。乌梁素海冰封期优势藻类种类组成见表4。
2.3 冰体和水体的对比分析
图 6为乌梁素海冰封期营养盐和藻类密度的时间变化曲线。从时间分布上看,冰封期水体中的TN、TP、COD含量和藻类密度均呈上升趋势,且明显高于冰体中的含量,其中,水体和冰体中的TN质量浓度均值分别为2.076和1.255 mg·L-1,即水体中的TN是冰体中的1.65倍;TP质量浓度均值分别为0.875和0.361 mg·L-1,水体是冰体中的2.42倍;COD含量均值分别为 88.56和 28.83 mg·L-1,水体是冰体中的 3.07倍;藻类密度均值分别为3.145和1.64×106cell·L-1,水体是冰体中的1.92倍;该现象可以说明,在冰层的形成和生长过程中,冰体对TN、TP、COD和浮游植物均有不同程度的排斥作用,其中以 COD为最强;而其造成的直接结果就是TN、TP、COD和浮游植物由冰体逐步迁移至水体。

表4 乌梁素海冰封期优势藻类种类组成Table 4 The species composition of dominant algea during icebound season in Wuliangsuhai Lake

3 结论及建议
3.1 结论
通过对冰封期乌梁素海的冰体和水体进行研究,得出的结论如下:
1)冰封期乌梁素海的冰厚介于0.22~0.74 m之间,水深在0.28~2.45 m之间,且水深和冰厚存在较好的负相关性。
2)在冰体的形成和生长过程中,营养盐和浮游植物逐渐从冰层迁移至水体中,使得冰封期水体中的营养盐和浮游植物的含量均大于冰体中的,结冰过程使得湖泊水体的富营养化状态恶化。
3)水体中 TN、TP、COD的浓度和浮游植物密度分别是冰体中的 1.85、2.42、3.07、1.29倍,这说明冰体对各污染物均有不同程度的排斥效应,其中以 COD最强;同时,也说明湖冰对有机物的排斥效应要高于其对无机物的。
4)乌梁素海冰封期浮游植物结构包括绿藻、硅藻等7门67属105种(含变种),其中以绿藻、硅藻种类数量居多,分别占总种数的 33.3%和26.7%,这表明冰封期乌梁素海浮游植物种类组成呈绿藻—硅藻型。
3.2 建议
1)由于湖水在结冰过程中,营养盐和浮游植物逐渐从冰层迁移至水体中,从而加剧了湖泊水体的富营养化状态,因此,冰封期湖泊的治理更加刻不容缓。
2)由于湖冰对污染物有不同程度的排斥效应,针对这一特点,可将其利用于降低有机物含量的相关研究中。例如,郝利娜和张维佳(2007)通过室内冷冻实验,证明了冷冻净化法能去除生活污水中80%的COD和90%的NH3-N。
CHO H, SHEPSON P B, BARRIE L A, et al.2002.NMR investigation of the quasi-brinelayer in ice/brine mixtures[J].J Phys Chem B, 106:11226-11232.
KNIGHT C A.1979.Observations of the morphology of melting snow[J].J Atmos Sci, 36: 1123-1130.
MATSUO T, SASYO Y.1981.Empirical formula for the melting rate of snowflakes[J].J Meteor Soc Japan, 59:1-8.
傅鑫廷.2009.低温及冰封条件下富营养化水体藻类分布规律研究[D].长春: 吉林大学, 35-36.
高红杰, 康春莉, 张歌珊, 等.2008.间甲酚在冰相中分布和释放规律的室内模拟研究[J].科学技术与工程, 8(10): 2731-2735.
龚乃虎.1999.关于冰晶雪花融化问题的研究——进展与展望[J].高原气象, 18(3):368-376.
国家环境保护总局.2002.水和废水监测分析方法第四版[M].北京:中国环境科学出版社: 109-125.
韩博平,林旭钿,李铁.2003.广东省大中型水库富营养化现状与防治对策研究[M].北京: 科学出版社, 7-8.
郝利娜,张维佳.2007.自然冷冻法处理生活污水的研究初探[J].中国科技信息, (23):18-19.
姜慧琴.2011.乌梁素海营养盐在冰体中的空间分布及其在冻融过程中释放规律的试验研究[D].呼和浩特: 内蒙古农业大学, 33-34.
金相灿,屠清瑛.1990.湖泊富营养化调查规范[M].2版.北京: 中国环境科学出版社, 18-215.
李卫平,李畅游,史小红,等.2008.内蒙古乌梁素海氮、磷营养元素分布特征及地球化学环境分析[J].资源调查与环境, 2(29):131-132.
李文霞,冯海艳.2006.水体富营养化与水体沉积物释放营养盐[J].地质通报, 25(5):602-608.
刘红,马徐发,熊邦喜.2006.武汉南湖的浮游植物[J].淡水渔业, 36(1):32-35.
吕超,李卫平,于玲红.2012.乌梁素海冰封期水体富营养化特征分析[J].安徽农业科学, 40(29):56-57.
马放, 杨基先, 魏利, 等.2010.环境微生物图谱[M].北京: 中国环境科学出版社, 2-190.
徐立冲,陆柱,蔡玲玲.1997.真空冷冻海水中冰晶生长速度的研究[J].水处理技术, 23(5): 280-284.
徐兆安,马桂芬,吴浩东,等.2012.内蒙古典型湖泊夏季浮游植物群落特征及与环境变量的关系[J].水生态学杂志, 4(33):58-59.
张岩.2012.乌梁素海结冰过程中污染物迁移机理及其应用研究[D].呼和浩特: 内蒙古农业大学,: 80-82.
周凤霞, 陈剑虹.2011.淡水微型生物与底栖动物图谱[M].2版.北京:化学工业出版社, 8-58.
周敏.2012.惠州西湖浮游植物群落特征及其与氮、磷营养盐关系的研究[D].广州: 暨南大学, 7-8.
