柳杉初级种子园遗传多样性1)
2014-08-02刘子梁欧阳磊黄信金翁怀峰施季森
徐 进 刘子梁 欧阳磊 黄信金 翁怀峰 施季森
(林木遗传与生物技术省部共建教育部重点实验室(南京林业大学),南京,210037)(福建省林业科学研究院) (福建省霞浦杨梅岭林场) (林木遗传与生物技术省部共建教育部重点实验室(南京林业大学))
柳杉初级种子园遗传多样性1)
徐 进 刘子梁 欧阳磊 黄信金 翁怀峰 施季森
(林木遗传与生物技术省部共建教育部重点实验室(南京林业大学),南京,210037)(福建省林业科学研究院) (福建省霞浦杨梅岭林场) (林木遗传与生物技术省部共建教育部重点实验室(南京林业大学))
采用SSR分子标记技术,对福建霞浦杨梅岭国有林场柳杉初级种子园12个种源地,共计96份优良无性系植株进行遗传多样性的分析,结果表明:26对SSR分子标记检测出柳杉种子园处于较高的遗传多样性水平,共扩增得到103个多态位点。平均等位基因数3.961 5个,平均有效等位基因数2.728 2个;平均观察杂合度为0.496 6,Shannon信息指数平均为0.994 5,Nei多样性指数平均为0.533 6。其遗传变异主要存在于种源内, 不同种源基因分化系数为0.167 0。
柳杉;SSR;初级种子园;遗传多样性
Cryptomeriafortune; SSR; Primary seed orchard; Genetic diversity
柳杉属植物分为柳杉(Cryptomeriafortune)和日本柳杉(CryptomeriajaponicaD. Don)两个种,其中柳杉为我国特有树种,柳杉种内变异丰富,叶色、枝条类型及树姿等存在各种变异,具有选育不同类型的巨大潜力和极高的用材和观赏价值,其天然林主要分布在中国东南部的天目山、武夷山等地,江苏、安徽、湖南、湖北等地均有引种栽培[1]。遗传多样性是生物多样性的基础,种内的多样性是物种以上各水平多样性最重要的来源。遗传变异、生活史特点、种群动态及其遗传结构等决定或影响着一个种群与其它物种及其环境相互作用的方式。不同物种和不同个体间的多态性,最终都会体现在基因的多态性上,表现在DNA分子水平的多样性分析。而分子标记可以提供大量的遗传变异信息,是进行群体遗传结构和育种群体遗传关系研究的有力工具[2]。分子标记提供的遗传变异信息可以辅助制定核心群体收集的策略,以保障育种材料的高水平遗传变异[3]。
利用分子标记研究遗传多样性已在林木种质资源上广泛开展,柳杉属中的另一个种——日本柳杉在此方面已开展一些有效的研究[4-9],而我国的柳杉遗传特性也进行了一些研究和探讨[10-12],但对柳杉遗传种质资源尚未进行系统地研究,特别是利用分子技术手段。本研究正是利用SSR分子标记对来自福建霞浦杨梅岭的初级种子园进行遗传多样性的分析。以期为柳杉种质资源保护和对柳杉育种群体的长期经营具有重要指导意义。
1 材料与方法
取自福建省霞浦杨梅岭国有林场初级种子园,地处太姥山系,属浙闽东南沿海丘陵立地亚区,种源林植株收集了12个种源地,它们是浙江的石墙、天目、文成、江山、临海、福建的霞浦、莆田、仙游、福鼎、周宁、南平、三明,在每个种源中分别选取7~10个无性系单株,共有96份样本。所有样品均用用冰盒保存带回实验室后立即存入冰箱-80 ℃保存备用。
DNA提取:采用CTAB法进行柳杉DNA的提取。①液氮研磨;②CTAB缓冲液水浴30 min;③V(氯仿)∶V(异戊醇)=24∶1,抽提3~4次至无沉淀为佳;④放置冰箱-20 ℃保存后用异丙醇沉淀;⑤75%乙醇洗涤沉淀;⑥风干后溶于ddH2O中,-4 ℃度保存备用。用前将纯化后的DNA稀释至20 ng/μL。
引物筛选:从106对来自日本柳杉的SSR引物中进行筛选[4-7,9],由Introvigen公司合成。从材料中随机选出7个不同样本,利用聚丙烯酰胺凝胶电泳,经固定、银染、显影等步骤,最终筛选出具有多态性的SSR引物用于柳杉初级种子园遗传多样性分析。
多样性分析:通过人工读带的方式将结果记录下来,再利用Popgen32分析柳杉初级种子园遗传多样性,分析的参数有:等位基因数、有效等位基因数、Shannon信息指数、基因多样度指数。
2 结果与分析
2.1 柳杉SSR多态性引物与扩增结果
从106对日本柳杉SSR引物中,选出25对具有多态性的引物用于柳杉初级种子园遗传多样性分析,图1是引物Cjgssr31、BY900902、BY888980对部分群体进行检测的效果图。


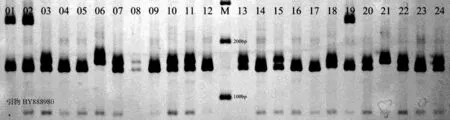
图1 引物扩增效果图
2.2 柳杉初级种子园亲本遗传多样性
2.2.1 等位基因多样性
通过Popgen32软件分析,26对SSR引物对种子园12个种源群体共扩增出103个多态位点,其等位基因相关数据见表1。由表1知等位基因数最少2个,最多8个,平均等位基因数3.961 5,平均有效等位基因数2.728 2。Shannon信息指数为0.987 1,Nei多样性指数平均为0.536 2。其中BY894091位点多样性程度最低,仅在南平种源有某一优良无性系(编号91)中表现为杂合子状态,且和约500 bp左右片段一同扩增,其余无性系均为纯合子,初步推测该无性系在该位点的突变可能与表型性状相关。通过该数据还发现,柳杉在物种水平上,Shannon信息指数的范围为0.033 2(位点BY894091)到1.830 2(位点CS1525),不同位点对Shannon信息指数贡献不同,差异较大,平均Shannon信息指数为0.994 5,遗传多样性比较丰富,也说明柳杉种子园的遗传结构优良,可为种子园建设提供丰富的原材料。
外科护士VTE预防知识子问卷得分(2.50±1.47)分,平均得分率为25.0%。各条目具体情况见表2、表3。
2.2.2 遗传杂合度
通过表2可以看出,在物种水平上,观察杂合度最低的是位点BY894091的0.010 6,最高的是Cjgssr7、Cjgssr181、CS1525的1.000 0,平均观察杂合度为0.496 6;期望杂合度由最低位点BY894091的0.010 6到最高的CS1525位点的0.821 4,平均期望杂合度为0.536 5。Nei多样性指数由位点BY894091的0.010 6到位点CS1525的0.817 1,平均Nei多样性指数为0.533 6。Shannon信息指数和Nei多样性指数基本一致。和其他裸子植物相比,柳杉在物种水平上同样表现出较高的遗传多样性。

表1 不同种源柳杉等位基因相关数据
2.3 柳杉初级种子园遗传结构
柳杉12个种源遗传多样性数据汇总见表2。根据表2可知,等位基因数最低的为江山种源1.730 8,最高的为天目种源3.692 3,平均观察等位基因数为2.769 2,平均有效等位基因数为2.216 8。
由表3可知,在群体水平上,Shannon信息指数由高到低依次为霞浦种源、天目种源、莆田种源、石墙种源、仙游种源、福鼎种源、周宁种源、南平种源、文成种源、临海种源、三明种源、江山种源,其中最低的江山Shannon信息指数为0.419 8,最高的霞浦Shannon信息指数为0.968 8,平均为0.7596。Nei多样性指数由最低的江山0.274 0到最高的霞浦0.533 3,平均为0.446 6,多样性变化较大,群体间相似性程度偏低,说明柳杉在种群水平上存在较高的变异,和Shannon信息指数基本保持一致。

表2 种源林位点杂合度指数
杂合度是用来衡量种群基因座上出现杂合状态所占整个种群的比率。从表3知柳杉在群体水平上,平均观察杂合度为0.482 4,平均期望杂合度为0.501 3。观察杂合度和期望杂合度基本保持一致,说明采样群体可以很好地反应柳杉种源遗传多样性状况。
2.4 不同种源多样性指数方差分析
2.5 种源基因分化系数和基因流
由Popgen32分析12个柳杉种源遗传分化系数和基因流得知(表5),平均遗传分化系数为0.167 0,说明16.7%的分化存在于柳杉群体间,83.3%的分化存在于群体内部,大部分遗传变异存在于群体内,说明柳杉群体内存在较强的遗传分化能力。
因为当基因流>1时,种群间存在一定的基因交流,使群体趋向于Hardy-Winberg平衡;基因流<1时,就说明种群间的基因交流程度较低,容易造成种群间的遗传分化;基因流>4时,各群体为一个随机单位[13]。由表5分析可知,12个柳杉种源得出其平均基因流基因流为1.247 5(大于1),说明不同种源柳杉群体间在分化的同时还存在一定的基因交流。这可能说明在历史上,尽管柳杉呈间断的或孤立的分布状态,但由于人类的活动引起各地相互引种栽培,使得柳杉在不同种源间得以进行基因交流。
从固定指数上观察(表5),26个位点当中有10个位点的固定指数小于0,另外16个位点均大于0,平均为0.079 2,说明本研究中柳杉12个种源的26个位点中纯合体占优势。

表3 种子园遗传多样性分析

表4 柳杉13个种源多样性指数方差分析
注:** 表示在0.01水平上差异极显著。

表5 遗传分化系数和基因流
2.6 不同种源间遗传距离和聚类分析
通过UPGMA聚类,不同种源柳杉聚类结果见图2。由图2可将柳杉分为3类群,即临海群体为一类,文成群体和江山群体为一类,剩下的三明群体、南平群体、福鼎群体、周宁群体、仙游群体、霞浦群体、石墙群体、莆田群体、天目群体为一类,而且从聚类图来看,它们之间存在逐步分化的关系,其中天目群体和莆田群体亲缘关系最近,其次是石墙群体、霞浦群体、仙游群体、周宁群体。聚类分析结果和柳杉实际地理距离有一定的差异,反应了在历史时期中,人类的活动造成柳杉在不同地理位置的传播和繁殖可能对柳杉种群遗传结构的分布造成一定的影响。

图2 柳杉12个种源间亲缘关系UPGMA聚类图
3 结论与讨论
使用26对多态性引物对12个地理种源柳杉进行遗传多样性分析,结果共扩增出103个多态位点,平均等位基因数3.961 5个,平均有效等位基因数2.728 2,平均Shannon信息指数0.994 5,Nei多样性指数平均为0.533 6。在群体水平上,Shannon信息指数由由高到低依次为霞浦群体、天目群体、莆田群体、石墙群体、仙游群体、福鼎群体、周宁群体、南平群体、文成群体、临海群体、三明群体、江山群体,其中最高的霞浦群体0.968 8,最低的江山群体0.419 8,多态性信息丰富。
在群体水平上,Nei多样性指数最低的江山为0.274 0,最高的霞浦为0.533 3,多样性变化较大,种群间相似性程度偏低,说明柳杉种群内部存在较高的遗传变异。观察杂合度和期望杂合度基本保持一致,说明采样群体可以很好地反应柳杉种源遗传多样性状况。Nei多样性指数平均为0.533 6,和其他研究林木植物遗传多样性得出的结果相比:日本柳杉(C.japonica)Nei指数为0.666[5],马尾松(Pinusmassoniana)为0.543 8[14],北美乔松(P.strobus)为0.515[15],小干松(P.contorta)为0.758[16],柽柳(Tamarixchinensis)为0.456[17],挪威云杉(Piceaabies)为0.789[18],栓皮栎(Quercusvariabilis)为0.804[19]。从这些数据中可以看出,柳杉的遗传多样度水平处于林木树种中的中等水平。
12个种源的柳杉基因分化系数为0.167 0,即16.7%的遗传分化存在与种群间,83.3%的遗传分化存在于种群内,说明柳杉不同种源间存在较强的分化能力。和其他一些林木树种基因分化系数相比:日本柳杉(C.japonica)天然群体为0.028[9],栓皮栎(Q.variabilis)为0.045 519,海岸松(P.pinaster)为0.111 0[20],脂松(P.Resinosa)为0.280[21],资源冷杉(Abiesziyuanensis)为0.250[22],云杉(Piceaasperata)为0.362[23],密叶红豆杉(Taxusfuana)为0.584 2[24]。柳杉的遗传分化系数也处于中等偏低水平。日本柳杉天然群体间的遗传分化系数如此低的原因可能是柳杉属植物属于风媒植物,异花授粉,且世代周期长,造成遗传分化不明显[9]。
在基因流方面,柳杉不同种源基因流为1.247 5,大于1,说明柳杉不同种源间存在一定的基因交流。在其他一些树种方面,基因流有不同的表现,例如云杉(P.asperata)为0.55[23],栓皮栎(Q.variabilis)为5.23[19],马尾松(P.massoniana)为1.03[14],密叶红豆杉(T.fuana)为0.355 8[24]。不同的树种基因流差别较大,这可能与树种的生理特性、地理分布和人类活动有关。一般认为,植物的基因流动主要靠花粉的扩散和种子的传播实现的,这也是植物基因流的主要形式[18]。借助风力可以将柳杉花粉传播很远的距离,加上各地相互引种栽培,即使柳杉种群呈间断的或孤立的分布状态,也可以实现一定的基因交流。
聚类分析发现,12个种源柳杉可分为3类,即临海群体为一类,文成群体和江山群体为一类,剩下的三明群体、南平群体、福鼎群体、周宁群体、仙游群体、霞浦群体、石墙群体、莆田群体、天目群体为一类。
[1] 中国科学院中国植物志编辑委员会.中国植物志:41卷[M].北京:科学出版社,2005.
[2] Smith J S C, Smith O S. Fingerprinting crop varieties[J]. Advances in Agronomy,1992,47: 85-129.
[3] Brown A H D. Core collections:a practical approach to genetic resourses management[J]. Genome,1989,31:818-824.
[4] Tani N, Takahashi Tiwata H, Mukai Y, et al. A consensus linkage map for sugi (Cryptomeriajaponica) from two pedigrees, based on microsatellites and expressed sequence tags[J]. Genetics,2003,165(3):1551-1568.
[5] Moriguchi Y, Iwata H, Ujino I T. Development and characterization of microsatellite markers forCryptomeriajaponicaD. Don[J]. Theoretical and Applied Genetics,2003,106(4):751-758.
[6] Tani N, Takahashi T, Ujino I T, et al. Development and characteristics of microsatellite markers for sugi (CryptomeriajaponicaD. Don) derived from microsatellite-enriched libraries[J]. Annals of Forest Science,2004,61(5):569-575.
[7] Roberts S, Romano C, Gerlach G. Characterization of EST derived SSRs from the bay scallop, argopecten irradians[J]. Molecular Ecology Notes,2005,5(3):567-568.
[8] Tomoyuki Kado, Hiroshi Yoshimaru, Yoshihiko Tsumura. DNA Variation in a Conifer, Cryptomeria japonica (Cupressaceaesensulato)[J]. Genetics,2003,165(3):1547-1559.
[9] Tomokazu Takahashi, Naoki Tani, Hideaki Taira, et al. Microsatellite markers reveal high allelic variation in natural populations ofCryptomeriajaponicanear refugial areas of the last glacial period[J]. Journal of Plant Research,2005,118(2):83-90.
[10] 黄信金.柳杉种源变异与联合选择[J].浙江林学院学报,2010,27(6):884-889.
[11] 欧阳磊.柳杉种子园亲本遗传变异规律与选择[J].中南林业科技大学学报, 2010,30(3):90-94.
[12] 黄信金.柳杉半同胞子代遗传变异与优良遗传型选择[J].中南林业科技大学学报,2010,30(7):50-54.
[13] Wright S. Evolution and the genetics of populations[M]. Chicago: University of Chicago Press,1984.
[14] 艾畅,徐立安,赖焕林,等.马尾松种子园的遗传多样性与父本分析[J].林业科学,2006,42(11):146-150.
[15] Echt C S, May Marquardt P M, Zahorchak R, et al. Characterization of microsatellite markers in eastern white pine[J]. Genome,1996,39(6):1102-1108.
[16] Thomas B R, Macdonald S E, Hicks M, et al. Efects of reforestation methods on genetic diversity of lodgepole pine: all assessment using microsatelite and randomly amplified polymophic DNA markers[J]. Theor Appl Genet,1999,99(3/4):793-801.
[17] 李锐.柽柳SSR标记开发及群体遗传结构分析[D].南京:南京林业大学,2007.
[18] Pfeiffer A, Olivieri A M, Morgante M. Identification and characterization of microsatellites in Norway spruce (PiceaabiesK.)[J]. Gemome,1997,40(4):411-419.
[19] 徐小林, 徐立安, 黄敏仁, 等.栓皮栎天然群体SSR遗传多样性研究[J].遗传,2004,26(5):683-688.
[20] Mariette S, Chagne D, Lezier C, et al. Genetic diversity within and among Pinus pinaster populations: comparison between AFLP and microsatellite markers[J]. Heredity,2001,86(4):469-479.
[21] Jacquelyn Boys, Marilyn Cherry, Selvadurai Dayanandan. Microsatellite analysis reveals genetically distinct populations of red pine (Pinusresinosa, Pinaceae)[J]. American Journal of Botany,2005,92(5):833-841.
[22] Tang Shaoqing, Dai Wenjuan, Li Mingshun, et al. Genetic diversity of relictual and endangered plant Abies ziyuanensis (Pinaceae) revealed by AFLP and SSR markers[J]. Genetica,2008,133(1):21-30.
[23] 罗建勋,杨龙华,辜云杰,等.云杉天然群体遗传多样性的综合评价[J].西北农林科技大学学报:自然科学版,2011,39(4):69-76.
[24] Amin Shah, Li Dezhu, Gao Lianming, et al. Genetic diversity within and among populations of the endangered species Taxus fuana (Taxaceae) from Pakistan and implications for its conservation[J]. Biochemical Systematics and Ecology,2008,36(3):183-193.
1) 江苏省科技支撑计划(BE2010312);江苏省高校自然科学研究重大合同(11KJA220003);国家林业公益性行业科研专项(201304104)资助。
徐进,女,1965年6月生,林木遗传与生物技术省部共建教育部重点实验室(南京林业大学),教授。E-mail:xjinhsh@njfu.com.cn。
2013年7月20日。
S722
Genetic Diversity ofCryptomeriafortuneifrom Primary Seed Orchard/Xu Jin, Liu Ziliang(Key Laboratory of Forest Genetics & Biotechnology, Nanjing Forestry University, Nanjing 210037, P. R. China); Ouyang Lei(Fujian Academy of Forestry); Huang Xinjin, Weng Huaifeng(State-owned Forest Farm of Fujian Yangmeiling); Shi Jisen(Key Laboratory of Forest Genetics & Biotechnology, Nanjing Forestry University)//Journal of Northeast Forestry University.-2014,42(4).-1~5
责任编辑:潘 华。
We used SSR molecular maker to detect genetic diversity ofCryptomeriafortuneifrom Fujian Xiapu Yangmeilin National Seed Orchard. 96 are excellent clones collected from 12 different provenances. A relatively high level of genetic diversity is detected inC.fortunei’sspecies with 26 polymorphic microsatellite loci. 103 loci are polymorphic markers. Average number of alleles and effective number of alleles are 3.961 5 and 2.728 2, respectively. The mean expected heterozygosity is 0.496 6. Shannon information index is 0.987 1, Nei diversity index is 0.536 2. The majority of genetic variation is within provenance (0.167 0).
