酱油二次沉淀蛋白质的分离、鉴定及氨基酸分析
2014-05-10孙鹏飞1高献礼1闫爽1陆健1
孙鹏飞1,2,3,4,高献礼1,2,3,闫爽1,2,3,陆健1,2,3,4*
(1.江南大学粮食发酵工艺与技术国家工程实验室,江苏 无锡 214122;2.江南大学工业生物技术教育部重点实验室,江苏 无锡 2141222;3.江南大学生物工程学院,江苏 无锡 214122 4. 宿迁市江南大学产业技术研究院,江苏 宿迁 223800)
酱油起源于我国,是我国的传统调味品,已有2500多年的历史。近年来,我国酱油行业呈现产销两旺的景象,酱油年销售量已经突破600万吨[1]。在此期间,我国酱油的质量也得到了不断的提高,但酱油二次沉淀问题仍然未得到较好的解决,是困扰我国酱油行业发展的重要质量问题[2]。酱油在巴氏杀菌后析出的沉淀称作一次沉淀,其可通过过滤消除,对终产品质量影响不大。酱油在储存和销售过程中瓶底出现的沉淀为二次沉淀[3],其严重影响我国酱油的外观质量,特别是与澄清的日本酱油对比时,这种外观问题严重影响消费者的购买倾向。目前,日本酱油的价格是我国酱油的8~10倍,但仍垄断了欧美市场并对国内的高端市场造成了较大冲击,其中,我国酱油的二次沉淀问题是导致两者价格差异较大和我国酱油在高端市场缺乏竞争力的重要原因之一[4]。
目前,国内学者和本实验的前期工作主要研究了酱油二次沉淀的主要成分、影响其形成的因素及我国酱油二次沉淀含量的分布[2-6]。日本学者对引起酱油二次沉淀的促进因子和引起酱油热凝集作用的酶分别进行了分离和鉴定[7-9]。由以上研究可知,目前国内外对二次沉淀的研究尚停留在常规指标分析和影响其形成因素的分析上,而对组成酱油二次沉淀蛋白质的质量分布、蛋白质来源及蛋白质氨基酸组成特征等信息的研究尚未见报道。
蛋白质被认为是引起酱油二次沉淀形成的关键因素,而深入了解酱油二次沉淀蛋白质的来源和氨基酸组成特征对解决酱油二次沉淀问题尤为重要[3-6]。因此本实验通过N-三(羟甲基)甘氨酸-十二烷基磺酸钠-聚丙烯酰胺凝胶电泳(Tricine-SDS-PAGE)和高效液相色谱(HPLC)技术对酱油二次沉淀中的主要成分蛋白质进行分析,并通过基质辅助激光解析电离飞行时间串联质谱(MALDI-TOF/TOF MS)技术追溯蛋白质的来源,以期对其中蛋白质有更深入的认识,为酱油二次沉淀问题的解决提供新的依据。
1 材料与方法
1.1 材料与仪器
酱油 购于江苏无锡华润万家超市,主要酿造原料为脱脂大豆、小麦、食盐,高盐稀态酿造,氨基酸态氮≥0.4g/100mL,食品添加剂有焦糖色、谷氨酸钠、苯甲酸钠、对羟基苯甲酸乙酯、黄原胶,产地北京,生产日期为 2012.5.31;Tricine和蛋白质标准品(分子量 4.1 ku-45.0ku) 上海生工生物工程公司;NH4HCO3、二硫苏糖醇(DTT)、碘乙酰胺(IAA)、三氟乙酸(TFA) 美国Sigma公司;其他试剂 均为分析纯。
垂直电泳仪 美国Bio-Rad公司;Ultraflextreme飞行时间串联质谱仪 德国Bruker公司;Agilent 1100型氨基酸专用高效液相色谱仪 美国安捷伦公司;H1850R型台式高速冷冻离心机 长沙湘仪离心机仪器有限公司;QT漩涡混合器 上海琪特分析仪器有限公司;JD-801凝胶成像仪 江苏省捷达科技发展有限公司。
1.2 实验方法
1.2.1 二次沉淀收集 酱油二次沉淀制备方法采用静置和冷冻干燥法[2-3],将酱油(500mL,玻璃瓶装)静置10d,虹吸法抽去上清,留下最后约20mL酱油和沉淀,摇匀后在4500r/min的条件下离心30min,获得酱油上清S1以及二次沉淀P1,P1经过冷冻干燥3d后便获得粉状沉淀P2(收集量约为0.75g/ L)。
1.2.2 酱油上清蛋白质收集 将收集过沉淀的酱油上清液S1与20%的三氯乙酸(TCA)溶液1:1混合,在冰浴条件下沉降1h,混合物在10000r/min的条件下离心10min,弃上清,获得上清蛋白质S2。
1.2.3 凝胶电泳分析 将二次沉淀P2和上清蛋白质S2复溶于溶剂中(含8mol/L尿素、1%SDS、1%β-巯基乙醇、0.05mmol/L Tris),在漩涡振荡器上混合均匀后,10000r/min离心5min,取上清用于电泳。
Tricine-SDS-PAGE分析 由于成品酱油中多肽多在10ku以下[10],本研究采用了可以分析小于10ku多肽的Tricine-SDS-PAGE凝胶系统,参照曹佐武[11]改良的实验方法,浓缩胶浓度5%,分离胶浓度16%,阳极缓冲液Tris 12.11g定容至500mL,6mol/L HCl调pH至8.9,阴极缓冲液Tris 6.06g、Tricine 8.96g、SDS 0.5g定容至500mL,样品上样量均为15μL,30V电泳半小时,100V电泳至结束。凝胶在含有50%乙醇和10%冰醋酸的固定液中固定半小时后,在考马斯亮蓝R-250染色液中染色3h,然后用含10%甲醇和10%冰醋酸的脱色液脱色至背景清晰,电泳图谱用JD-801凝胶成像仪记录,通过捷达801图像分析软件3.3分析。
1.2.4 MALDI-TOF/TOF MS鉴定 将凝胶上蛋白质条带使用自制切点器切下,装于1.5mL进口EP管中,经脱色、脱水后用胰蛋白酶进行酶解,酶解液经抽提浓缩后,在Anchorchip靶板上进行点靶,待样品完全干燥后,将靶板放入仪器 Ultraflextreme MALDI-TOF MS/MS(Bruker)进行质谱数据的采集,使用肽段标准品(peptidecalibstandard II)进行校正,数据结果使用Bio Tools软件整合。搜索参数为:物种属为全物种;数据库NCBI;切割酶为胰酶;允许最大漏切位点为1,Mascot分值大于55即为鉴定成功。
1.2.5 氨基酸分析
1.2.5.1 游离氨基酸测定
酱油上清S1以及二次沉淀P2用TCA(浓度5%)静置沉淀1 h后用双层定性滤纸过滤,滤液经10000r/min离心10min,取上清液,利用高效液相色谱法测定游离氨基酸的含量。采用HYPERSIL OSD色谱柱(250mm×4.6mm,5μm),柱温40℃,紫外检测器波长338nm,洗脱流速1.0mL/min。
1.2.5.2 总氨基酸测定
酱油上清S1以及二次沉淀P2经过110℃水解22h,水解液经过过滤蒸酸等步骤后用高效液相色谱法测定氨基酸组成。高效液相色谱测定条件同游离氨基酸测定条件。
1.2.5.3 氨基酸疏水性分析
根据Lozano[12]的经验公式考察氨基酸的疏水性:
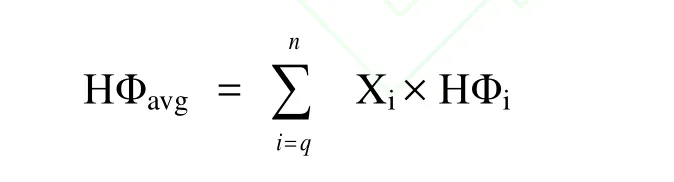
Xi表示各氨基酸摩尔百分含量,HΦi表示各氨基酸的疏水值。
1.2.6 数据分析 所有实验数据均测定3次,取其平均值。运用SPSS 11.5 软件进行方差分析。
2 结果与分析
2.1 电泳分析
图1为样品的Tricine-SDS-PAGE图谱,酱油上清S2主要有两条条带c和d,条带c经软件计算分子量为22.9ku,经积分光密度计算所占百分比为1.65%,d处条带连成一片,分子量约从2.0ku至14.0ku,可能是大豆蛋白质在酱油酿造过程中被蛋白酶切割成了分子量连续的多肽,所以电泳条带出现了连片的情况。条带d经积分光密度计算,占比例为98.35%,是S2中蛋白质的最重要组成部分。酱油沉淀P2主要有两条条带a和b,经软件计算分子量分别约为34.1ku和22.7ku,经积分光密度计算,两者分别占P2蛋白质的19.80%和80.20%。由以上分析可知,S2中主要的蛋白质是分子量从2.0ku到14.0ku的多肽;而P2中主要的蛋白质是一种分子量约为22.7ku的蛋白质,在S2中同样发现了一种分子量约为22.9ku的蛋白质,两者是否为同一种蛋白质需进行MALDI-TOF/TOF MS鉴定 。
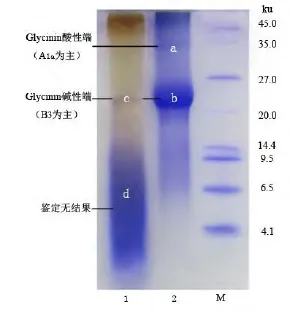
图1 Tricien-SDS-PAGE图谱Fig. 1 Tricine-SDS-PAGE electrophoresis of the samples
2.2 MALDI-TOF/TOF MS鉴定
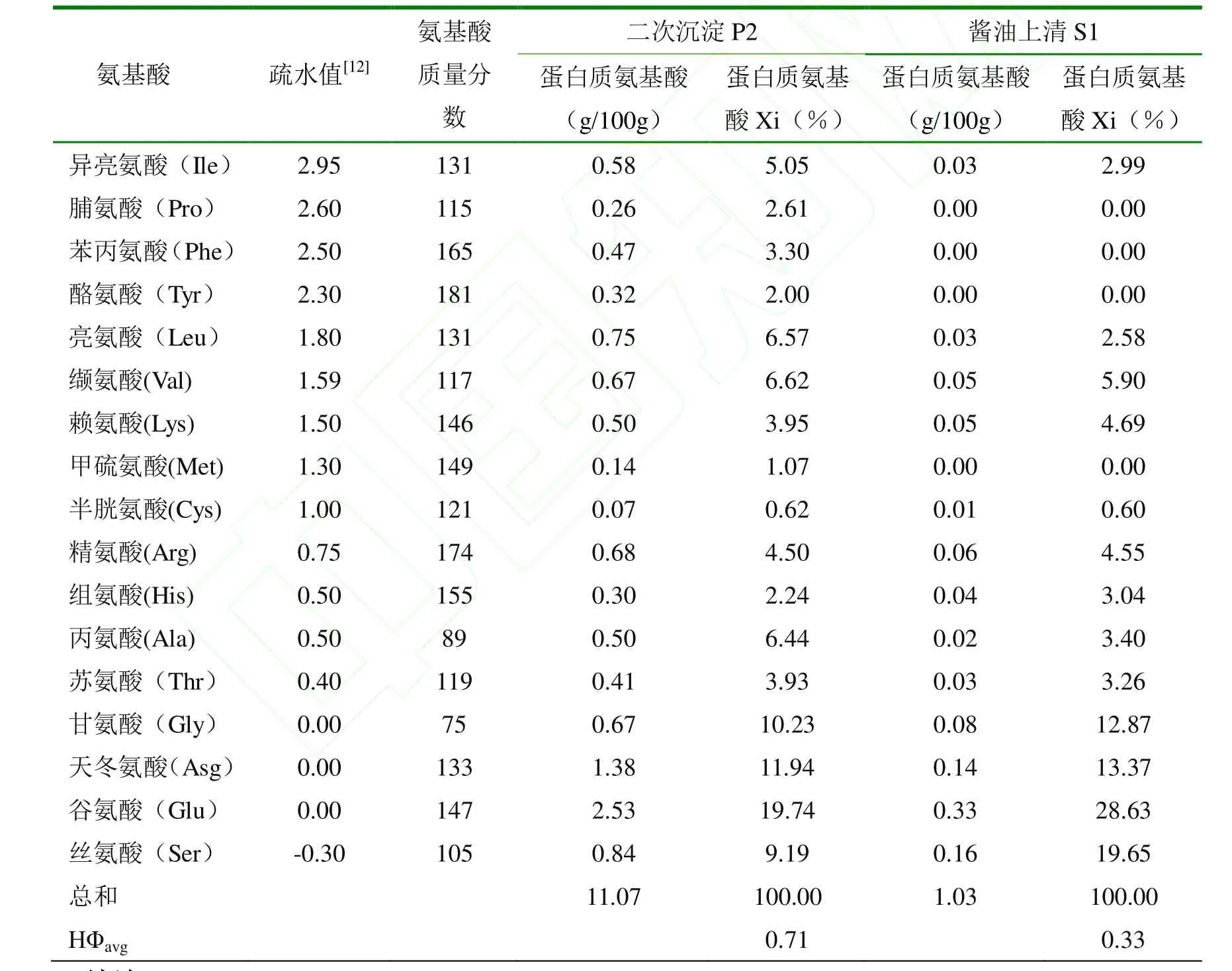
为了鉴定沉淀中蛋白质的来源及上清蛋白质是否与酱油二次沉淀蛋白质同源(条带 b和条带c),在图1中取点a、b、c、d进行 MALDI-TOF/TOF MS鉴定。条带a可信度最高的鉴定结果为大豆球蛋白(Glycinin) G1亚基,Mascot分值155,条带b和c可信度最高鉴定的结果为大豆球蛋白(Glycinin) G4亚基,Mascot分值分别为254和76,条带d未鉴定成功。大豆蛋白质主要含有7S和11S两个亚基,Glycinin是指11S 球蛋白(Globulin),Glycinin 中目前主要鉴定出的亚基有 G1(A1aB1b)、G2(A2B1a)、G3(A1bB2)、G4(A5A4B3)和G5(A3B4),每个亚基含有一个分子量大约在32ku左右的酸性端以及一个分子量在20ku左右的碱性端[13]。条带 a的分子量与 Glycinin亚基酸性端相吻合,条带 b和 c分子量与Glycinin亚基碱性端分子量相吻合。袁德保[14]曾测定了Glycinin各酸性端与碱性端的疏水性除了在其等电点下,大小依次为A4 酱油在经过巴氏消毒过滤等步骤出厂,澄清的酱油在货架上随时间逐步产生沉淀,沉淀并非一步形成的。由图1以及MALDI-TOF/TOF MS鉴定结果分析可得,P2中的蛋白质b是上清中的蛋白质c逐步沉降而来,即上清中的蛋白质c是沉淀蛋白质的主要来源。P2中蛋白质a应该也可以在S2中找到,可能因为其在S2中丰度太低而没有跑出可见条带。在图1中,虽然S2中的主要蛋白质都在条带d中,占了98.35%,但在沉淀P2中几乎找不到明显的对应条带,这说明被蛋白酶切割得到的多肽溶解性较好,不易沉淀。 表1是酱油上清S1和二次沉淀P2的氨基酸组成测定结果,蛋白质氨基酸定义为总氨基酸减去游离氨基酸[15](总氨基酸与游离氨基酸未在表中列出)。疏水值一栏数值越高疏水性越大。 在酱油上清S1和二次沉淀P2中,氨基酸摩尔百分比最高的均为谷氨酸,其次是丝氨酸、天冬氨酸和甘氨酸,这与袁德宝[14]测定的大豆蛋白质中各氨基酸含量的趋势基本相符。但这几种氨基酸在S1中所占比例分别比在P2中高45.03%、113.82%、11.98%和25.81%,它们的疏水值均等于小于0.00,是4种亲水性氨基酸。而疏水值大于等于1.00的氨基酸在P2中的比例则普遍高于S1,说明二次沉淀蛋白质中疏水性氨基酸比例高。值得注意的是,疏水值较高的脯氨酸、苯丙氨酸、酪氨酸和甲硫氨酸在S1中几乎检测不到,而在P2中却占有一定的比重。根据Lozano[12]经验公式计算的P2的平均疏水值为0.71,而S2的平均疏水值仅0.33,前者比后者高115.15%,方差分析显示两类蛋白质的平均疏水值存在显著性差异(p < 0.05)。上述分析结果与P2中的蛋白质是来源于酱油二次沉淀(不溶性蛋白质),S1中的蛋白质是来源于酱油上清液(水溶性蛋白质)的现象相吻合。同时也说明大豆蛋白质(或多肽)中部分蛋白质(或多肽)的高疏水性是导致其形成酱油二次沉淀的重要原因。 表1 氨基酸组成测定结果Table 1 Amino acid compostion analysis of sample 通过以上分析,主要得出以下结论: 3.1 酱油二次沉淀中的蛋白质分别是来源于大豆球蛋白质G4亚基碱性端B3和G1亚基酸性端A1a,两者分别占酱油二次沉淀蛋白质的80.20%和19.80%。碱性端B3和酸性段A1a是11S大豆球蛋白的重要组成部分,而11S大豆球蛋白是大豆中最重要的蛋白质来源。 3.2 酱油二次沉淀蛋白质氨基酸组成与酱油上清蛋白质相比存在明显差异,其蛋白质平均疏水值显著高于后者,这是酱油二次沉淀蛋白质氨基酸组成的主要特征,也是酱油二次沉淀蛋白质形成的重要原因。 因此,如果通过某种方式去除或者降解酱油中Glycinin G4亚基碱性端B3和G1亚基酸性端A1a,可有效解决酱油二次沉淀问题,这需要进一步的研究来证实。 [1]Gao XL, Cui C, Ren JY, et al. Changes in the chemical composition of traditional Chinese-type soy sauce at different stages of manufacture and its relation to taste[J]. International Journal of Food Science & Technology, 2011, 46(2):243-249. [2]高献礼,闫爽,陈燕斌,等. 基于统计学方法的酱油二次沉淀形成的初步研究[J]. 食品工业科技,2013, 34(03):188-191. [3]高献礼,孙鹏飞,闫爽,等. 酱油二次沉淀制备方法的对比[J]. 食品与发酵工业, 2013(1):46-50. [4]曾新安,李国基,于淑娟. 酱油沉淀成分研究[J]. 食品科学, 2002, 23(11):32-34. [5]陈有容,刘庆玮. 酱油中沉淀及混浊物产生的原因初探[J]. 天津微生物, 1989, 2:32-37. [6]张志航,李国基,于淑娟. 酱油沉淀物的溶解性能[J]. 华南理工大学学报, 2000, 28(11):59-63. [7]Tomita M, Motomura Y, Kitahara H. Purification and identification of the promoter of sediment formation from raw soy sauce by heating[J]. Journal of Fermentation and Bioengineering, 1998,86(4):373-378. [8]Tomita M, Motomura Y, Kitahar H a. Isomers and Homologues of L-Glutamic Acid 5-n-Butyl Ester as promoters of sediment formation in raw soy sauce [J]. Journal of Fermentation and bioengineering, 1999, 87(3):357-360. [9]Nakadai T, Nasuno S, Iguchi N. The action of peptidases from Aspergillus oryzae in digestion of soybean proteins[J]. Agricultural and Biological Chemistry, 1972, 36(2):261-268. [10]欧阳珊,冯云子,徐欢欢,等. 酱油制曲过程中蛋白质的降解规律研究[J]. 食品工业科技, 2012,33(7):195-197. [11]曹佐武. 有效分离 1kDa小肽的 Tricine-SDS-PAGE方法[J]. 中国生物工程杂志, 2004,24(1):74-76. [12]Lozano P, Combes D, Iborra JL. Food protein nutrient improvement by protease at reduced water activity [J]. Jounal of Food Science, 1994, 59(4):876-880. [13]Wang CM, Wu XL, Jia FX. Genetic variations of glycinin subunit genes among cultivated and wild type soybean species[J]. Progress in Natural Science, 2008, 18(1):33-41. [14]袁德保. 大豆蛋白热聚集行为及其机理研究[D]. 广州: 华南理工大学, 2010. [15]张志航,李国基,于淑娟. 酱油二次沉淀物氨基酸组分分析[J]. 齐齐哈尔大学学报, 2000,16(3):4-7.2.3 氨基酸分析结果
3 结论
