外源一氧化氮介导铜胁迫下番茄幼苗中铁、 锌、 锰的累积及亚细胞分布
2014-04-08梁国鹏姜春辉崔秀敏
张 敏, 梁国鹏, 姜春辉, 崔秀敏
(土肥资源高效利用国家工程实验室, 山东农业大学资源与环境学院,山东泰安 271018)
铜(Cu)是植物正常生命活动所必需的微量营养元素,广泛参与各种生命活动。同时,铜也是一种重金属元素,过量的 Cu 会对植物产生毒害作用,如破坏生理结构、 引发代谢紊乱、 抑制植物生长发育等问题[1-4]。目前,在农业生产中,污水灌溉和含铜的肥料及农药的大量使用,导致土壤系统中 Cu 不断累积,明显高于其背景值[5]。
一氧化氮(nitric oxide,NO)是广泛分布于生物体内的小分子信号物质。研究发现 NO参与植物的生长发育和对外界胁迫反应等多种生理过程。NO 能够提高逆境胁迫下植物抗性和生物量[6-7],维持组织结构稳定[8],参与植物激素的调控,提高植物对过多铜的螯合能力[9]。铁(Fe)、 锌(Zn)、 锰(Mn),作为微量元素亦在植物生命代谢中发挥着重要作用。如 Fe 元素参与光合作用和固氮过程,并在叶绿素合成中起关键作用; Zn 是植物体 59 种复合酶的组分或活化剂,并参与生长素代谢; Mn对种子发芽、 幼苗生长、 花粉管发育和植物茎的机械强度均有重要作用[10]。Fe2+、 Zn2+、 Mn2+在植物根系吸收及转运过程中与 Cu2+存在复杂的相互作用。研究发现,铜胁迫下,外源 NO 可调控番茄组织与细胞内过多的铜趋于更合理的分布[11],势必也会影响带有相同电荷的Fe2+、 Zn2+、 Mn2+的宏观及微域分布。
本文以番茄为试验材料,采用营养液培养,研究外源 NO 调控的 Cu 胁迫下番茄幼苗中 Fe、 Zn、 Mn 的累积及亚细胞分布,探索 Cu 胁迫下外源 NO 介导的番茄的营养机理。
1 材料和方法
1.1 供试材料
供试番茄(SolanumlycopersicumL.)为“改良毛粉802F1”。Hoagland营养液组成(mmol/L): Ca(NO3)2-4H2O 1.25、 KNO30.5、 NH4NO30.73、 KH2PO40.5、 H3BO34.6×10-2、 Na2MoO42.6×10-3、 MgSO41.0、 EDTA-Fe 2×10-2、 MnCl29×10-3、 ZnSO47.6×10-4,以上试剂均为分析纯,用蒸馏水配制。NO和Cu2+的供体及适宜浓度由预备试验完成, CuCl2提供Cu2+,硝普钠[Na2Fe(CN)5、 NO、 SNP购自Sigma公司]提供NO,先用蒸馏水配成200 μmol/L的母液,4℃避光保存,用时按所需浓度稀释。牛血红蛋白(Hb,购自Sigma公司)为NO的清除剂。
1.2 试验设计

1.3 测定项目与方法
1.3.1 生物量的测定 处理后第 8 d,移出番茄幼苗,测定各处理植株的株高,并将植株分为根系、 茎、 叶柄、 叶片四部分,用去离子水冲洗干净,吸水纸吸干表面水分,称鲜重。
1.4 数据处理
采用 Fe、 Zn、 Mn 转运因子(transport factor,TF) 计算 Fe、 Zn、 Mn 从根到地上部的运输特征[14]:
TF=(Cstem+Cleaf+Cpetiole) /Croot
式中: Cstem、 Cleaf、 Cpetiole和 Croot分别表示茎、 叶片、 叶柄和根系中的 Fe、 Zn、 Mn 的累积量。
采用Microsoft Excel软件对数据进行处理及绘图,采用 DPS 软件进行数据统计分析,采用最小显著差异法(LSD)进行多重比较和差异显著性检验(α=0. 05)。
2 结果与分析
2.1 外源 NO 对 Cu 胁迫下番茄幼苗生长的影响
从图 1 可以看出,不同处理下番茄幼苗的生物量和株高均有显著差异。与对照(CK)相比,Cu 胁迫处理(Cu)下番茄幼苗生物量和株高显著下降了33.7% 和 23.1%。添加外源 SNP (Cu+SNP)显著缓解了 Cu 胁迫引起的生长抑制,使番茄幼苗的生物量几乎恢复到对照水平,株高虽然没有恢复到对照水平,但仍显著高于Cu 胁迫处理。添加血红蛋白后(Cu+SNP+Hb),显著消除了 NO 的缓解效果,说明外源 NO 可以促进 Cu 胁迫下番茄幼苗的生长发育。

图1 外源一氧化氮对铜胁迫下番茄幼苗生物量和株高的影响Fig.1 Effects of exogenous NO on the biomass and height of tomato seedlings under Cu stress
2.2 外源 NO 对 Cu 胁迫下番茄幼苗 Fe 含量和累积量的影响
由表 1 可以看出,番茄幼苗各部位的 Fe 含量和累积量变化幅度较大,根系和叶柄的 Fe 含量和累积量最高。不同处理间各部位 Fe 含量和累积量差异显著。与对照(CK)相比,Cu 胁迫处理显著降低番茄幼苗根系、 茎、 叶柄和叶片的 Fe 含量,分别减少了 43.6%、 21.5%、 5.5% 和 13.7%。添加外源SNP(Cu+SNP)可显著提高Cu胁迫下番茄幼苗的Fe含量,根系、 茎、 叶柄、 叶片中Fe含量分别比Cu胁迫处理增加了 262.2%、 35.3%、 29.3%和 16.3%,添加血红蛋白后(Cu+SNP+Hb),番茄幼苗茎和叶柄中的Fe含量比Cu+SNP处理降低了15.6%和7.1%,根系和叶片中Fe含量增加了13.3%和 45.5%,均达到显著差异水平。
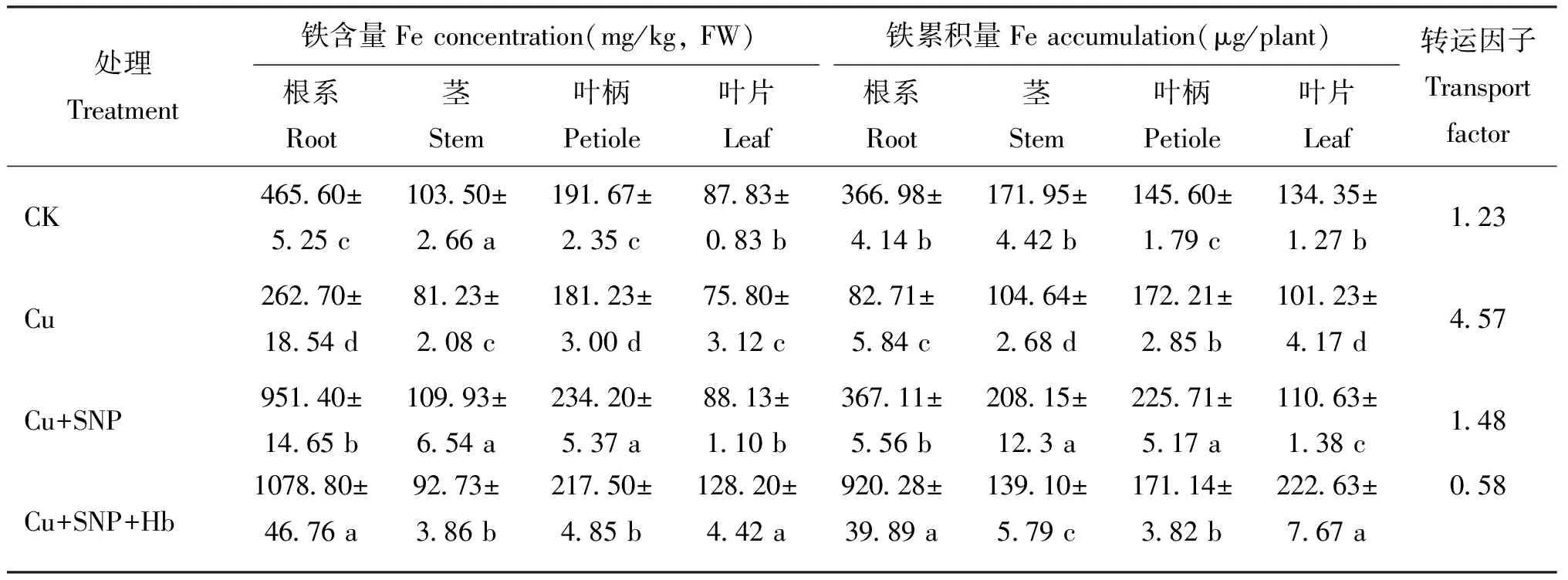
表1 番茄幼苗不同部位的铁含量和累积量
与 CK 相比,Cu 胁迫处理下,番茄幼苗各部位 Fe 累积量均显著降低。与 Cu 胁迫处理相比,Cu+SNP 处理显著提高番茄各部位 Fe 累积量。Cu+SNP+Hb 处理的番茄幼苗茎和叶柄中 Fe 含量显著降低,根系和叶片中 Fe 含量显著增加,但仍显著高于 CK 和 Cu 胁迫处理。
CK 处理的 Fe 转运因子 (TF) 为 1.23,Cu 胁迫处理 TF 为 4.57,添加外源 SNP 使 TF 降低为 1.48,表明外源 SNP 能显著降低根系中的 Fe 向地上部转运。
2.3 外源 NO 对 Cu 胁迫下番茄幼苗 Fe 的亚细胞分布的影响
表 2 显示,Cu 胁迫处理与对照(CK)相比,番茄幼苗根系细胞中细胞壁的 Fe 含量显著提高。 添加外源 SNP(Cu+SNP)可显著提高细胞器和液泡的 Fe 含量,分别比 Cu 胁迫处理提高了 54.5% 和 133.2%,而细胞壁的 Fe 含量显著降低 27.3%,差异均达显著水平。添加血红蛋白后(Cu+SNP+Hb),显著消除了 NO 的效果,与 Cu+SNP 处理相比,细胞壁和液泡 Fe 含量显著降低 35.1% 和 39.4%,而细胞器的 Fe 含量显著上升 21. 4%。表明番茄幼苗根系受到 Cu 胁迫后,外源 NO 可以提高细胞器中的活性铁,有利于细胞进行正常代谢。
与 CK 相比,Cu 胁迫处理提高了番茄幼苗茎细胞中细胞器和液泡的 Fe 含量。Cu 胁迫处理下,茎细胞中细胞器和液泡的 Fe 含量分别比CK 提高 49.2% 和 16.6%,而细胞壁的 Fe 含量显著降低 52.9%。添加外源 SNP(Cu+SNP) 后,细胞壁 Fe 含量占茎细胞 Fe 总含量的比例与 Cu 胁迫处理相比升高 10.6%,细胞器 Fe 含量所占比例升高 9.1%, 液泡 Fe 含量所占比例降低 19.7%。添加血红蛋白后(Cu+SNP+Hb),不同亚细胞的 Fe 含量均显著降低。
Cu 胁迫处理与 CK 相比,显著提高番茄幼苗叶柄细胞中细胞壁和液泡的 Fe 含量,分别比CK 提高 13.1% 和 15.3%, 而细胞器的 Fe 含量显著降低了 19.2%。与 Cu 胁迫处理相比,添加外源 SNP(Cu+SNP)后显著增加了番茄幼苗根细胞中不同亚细胞的 Fe 含量,细胞壁、 细胞器和液泡的 Fe 含量分别提高了 30.2%、 43.5% 和 18.8%。 添加血红蛋白后(Cu+SNP+Hb),细胞壁、 细胞器和液泡中的 Fe 含量比 Cu+SNP 处理分别降低了 11.3%、 14.9% 和 19.4%。
Cu 胁迫处理下,番茄幼苗叶片细胞中液泡的 Fe 含量比 CK 提高 8.9%,而细胞壁和细胞器则比 CK 分别降低了 11.6%、 20.5%。与 Cu 胁迫处理相比,Cu+SNP 处理的细胞器和液泡的 Fe 含量分别提高 14.9% 和 15.0% 细胞壁 Fe 含量降低了 24.0%。Cu+SNP+Hb 处理的细胞壁和细胞器的 Fe 含量占叶片细胞 Fe 总含量的比例,与 Cu 胁迫处理相比分别提高了 5.0% 和 4.3%,液泡的 Fe 含量所占比例下降了 9.3%。

表2 番茄幼苗中铁的亚细胞分布
2.4 外源 NO 对 Cu 胁迫下番茄幼苗 Zn 含量和累积量的影响
由表 3 可以看出,番茄幼苗各部位的 Zn 含量和累积量变化幅度较大,根系和叶片的Zn 含量和累积量最高。不同处理间番茄幼苗各部位的 Zn 含量和累积量差异显著。Cu 胁迫处理下,根系、 茎、 叶柄的 Zn 含量分别比 CK 减少了 85.1%、 19.8%、 15.0%,而叶片中 Zn 含量增加了 28.4%。Cu+SNP 处理可显著提高 Cu 胁迫下番茄幼苗根系、 叶柄和叶片的 Zn 含量,分别比 Cu 胁迫处理下提高了 8.3%、 27.9% 和 17.2%,而茎中 Zn 含量降低了 17.1%。Cu+SNP+Hb 处理的番茄幼苗茎、 叶柄和叶片中的 Zn 含量与 Cu+SNP 处理相比降低 18.0%、 7.4%和 21.3%,而根中的 Zn 含量为 Cu+SNP 处理的 3.83 倍,差异显著。
Cu 胁迫处理下,番茄幼苗根系和茎的 Zn 累积量比 CK 显著降低,叶柄和叶片中的 Zn 含量则显著增加。与 Cu 胁迫处理相比,添加外源 SNP 显著提高了番茄各部位 Zn 的累积量。添加血红蛋白后,番茄幼苗根系和叶片的 Zn 累积量显著提高,而茎和叶柄的 Zn 累积量显著降低。
CK 处理的 Zn 转运因子(TF)为 0.85,Cu胁迫处理后 TF 为 13.48,添加外源 SNP 使 TF 降低为 11.84,表明外源 SNP 能显著降低根系中的 Zn 向地上部转运。
2.5 外源 NO 对 Cu 胁迫下番茄幼苗 Zn 的亚细胞分布的影响
与 CK 相比,Cu 胁迫显著降低番茄幼苗根系不同亚细胞结构中的 Zn 含量,细胞壁、 细胞器和液泡的 Zn 含量分别降低 23.9%、 75.7%和35.9% (表4)。添加外源 SNP(Cu+SNP)可显著提高液泡的 Zn 含量,比 Cu 胁迫处理提高了 91.1%,细胞器的 Zn 含量仅提高了 0.7%,而细胞壁的 Zn 含量降低了 31.9%。添加血红蛋白后(Cu+SNP+Hb),显著消除了 NO 的效果,与 Cu+SNP 处理相比,细胞壁 Zn 含量显著上升了 95.7%,液泡的 Zn 含量降低 13.4%.而细胞器的 Zn 含量无明显变化。

表3 番茄幼苗不同部位的锌含量和累积量
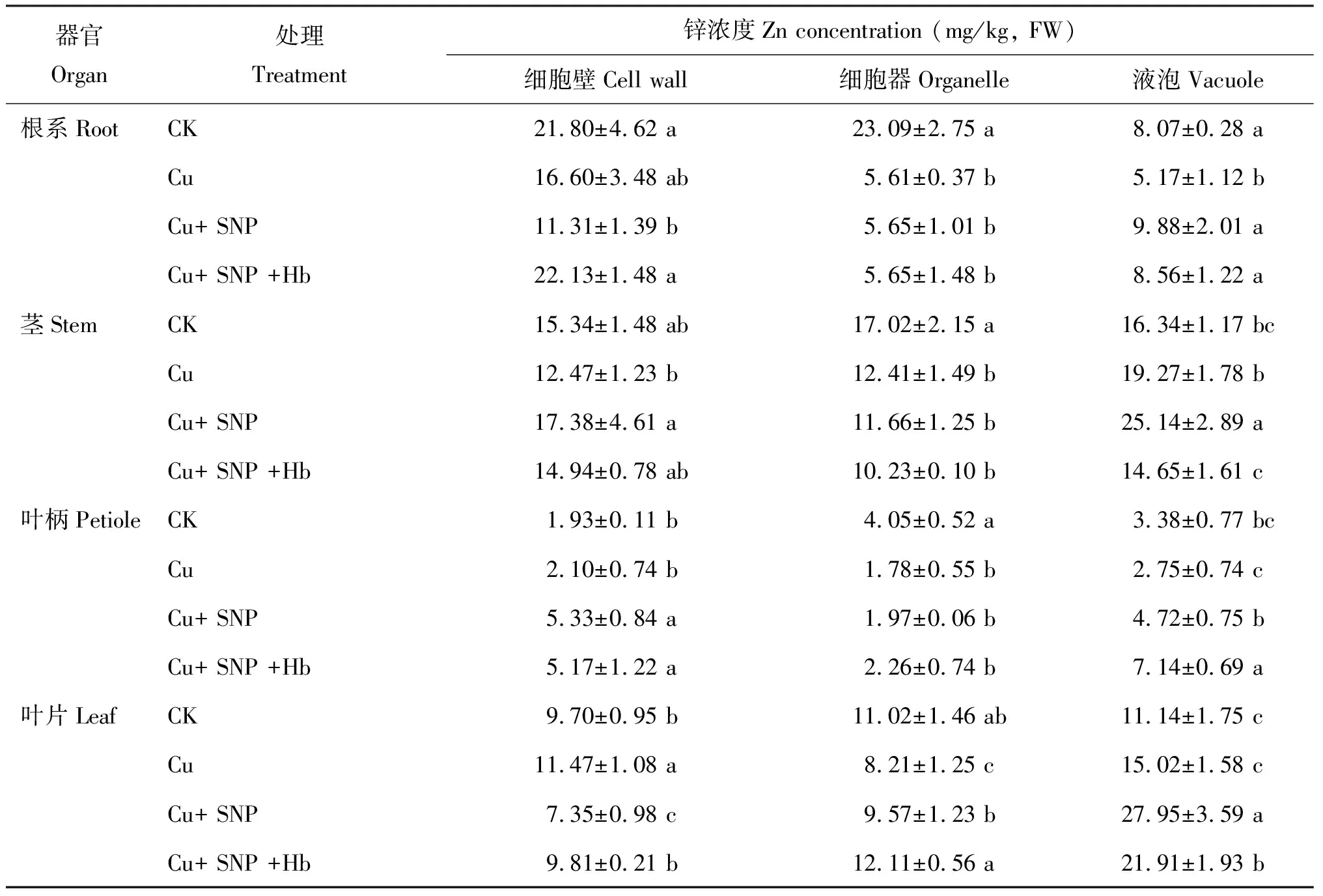
表4 番茄幼苗中锌的亚细胞分布
Cu 胁迫处理比 CK 番茄幼苗茎细胞中液泡的 Zn 含量有所提高。 Cu 胁迫处理下,番茄幼苗茎细胞中液泡的 Zn 含量比 CK 提高 17.9%,而细胞壁和细胞器 Zn 含量则比 CK 分别降低了 18.7% 和 27.1%。添加外源 SNP 后(Cu+SNP),细胞壁和液泡 Zn 含量占茎细胞 Zn 总含量的比例与 Cu 胁迫处理相比分别升高 3.8% 和 2.8%,细胞器 Zn 含量所占比例降低了 6.6%。添加血红蛋白后(Cu+SNP+Hb),不同亚细胞的 Zn 含量均显著降低。
Cu 胁迫处理显著提高了番茄幼苗叶柄细胞中细胞壁的 Zn 含量,比 CK 提高 8.8%,而细胞器和液泡的 Zn 含量分别降低 56.0%、 18. 6%。与 Cu 胁迫处理相比,添加外源 SNP (Cu+SNP)后显著增加番茄幼苗根细胞中不同亚细胞结构中的 Zn 含量,细胞壁、 细胞器和液泡的 Zn 含量分别提高了153.8%、 10.7%和71.6%。添加血红蛋白后,即 Cu+SNP+Hb 处理细胞壁中 Zn 含量比 Cu+SNP 处理降低 3.0%,但差异不显著,细胞器和液泡 Zn 含量分别提高了14.7% 和 51.3%。
Cu 胁迫处理下,番茄幼苗叶片细胞中细胞壁和液泡的 Zn 含量比 CK 分别提高了18.2% 和 34.8%,而细胞器中 Zn 含量降低 25.5%。添加外源 SNP 后,细胞器和液泡中 Zn 含量比 Cu 胁迫处理分别提高 16.6% 和 86.1%,而细胞壁中 Zn 含量下降 35.9%。添加血红蛋白后,细胞壁和细胞器的 Zn 含量占叶片细胞 Zn 总含量的比例与 Cu+SNP 处理相比分别提高了 6.0% 和 6.3%,液泡的 Zn 含量所占比例显著下降,降低幅度为 12.3%。
2.6 外源 NO 对 Cu 胁迫下番茄幼苗 Mn含量及累积量的影响
由表 5 可以看出,番茄幼苗各部位的 Mn 含量和累积量变化幅度较大,番茄幼苗各部位的 Mn 含量和累积量在不同处理间差异显著。与对照(CK)相比,Cu胁迫处理下,番茄幼苗根系、 叶柄和叶片 Mn 含量分别比对照增加了 15.2%、 20.7%、 29.2%,茎中 Mn 含量减少 39.3%。添加外源 SNP(Cu+SNP)可显著提高 Cu 胁迫下番茄幼苗根系和茎的 Mn 含量,分别比 Cu 胁迫处理下提高 14.6% 和 40.2%; 但叶柄和叶片 Mn 含量显著降低了 34.8% 和 30.2%。添加血红蛋白后(Cu+SNP+Hb),番茄幼苗根系和茎中的 Mn 含量降低了 5.1% 和 18.8%,而叶柄和叶片中的 Mn 含量显著增加了 72.7% 和27.3%。

表5 番茄幼苗不同部位的锰含量和累积量
与对照相比,Cu 胁迫处理下,番茄幼苗叶柄和叶片的 Mn 累积量显著增加,根系和茎中的 Mn 累积量显著降低。与 Cu 胁迫处理相比,添加外源 SNP 显著提高了番茄根系和茎中 Mn 的累积量,显著降低叶柄和叶片中 Mn 的累积量。添加血红蛋白后(Cu+SNP+Hb),番茄幼苗茎中 Mn 累积量显著降低,根系、 叶柄和叶片中 Mn 累积量显著增加。
CK 处理的 Mn 转运因子(TF)为 4.16,Cu 胁迫处理后转运因子为 6.70,添加外源 SNP 使 TF 降低为 6.01,表明外源 SNP 能显著降低根系中的 Mn 向地上部转运。
2.7 外源 NO 对 Cu 胁迫下番茄幼苗 Mn 的亚细胞分布的影响
由表 6 可知,与对照(CK)相比,Cu 胁迫显著降低番茄幼苗根系细胞中不同亚细胞的 Mn 含量,细胞壁、 细胞器和液泡的 Mn 含量分别降低 65.2%、 47.1%和82.8%。添加外源 SNP(Cu+SNP)可提高细胞器的 Mn 含量,比 Cu 胁迫处理提高 12.7%,而细胞壁和液泡的 Mn 含量分别降低 12.2% 和 6.0%。与 Cu+SNP 处理相比,添加血红蛋白后(Cu+SNP+Hb),番茄幼苗根系不同亚细胞结构的 Mn 含量显著提高,细胞壁、 细胞器和液泡的 Mn 含量分别提高了 252.7%、 101.1%和43.6%。

表6 番茄幼苗中锰的亚细胞分布
Cu 胁迫处理下,番茄幼苗茎细胞中细胞壁和细胞器的 Mn 含量比对照分别提高了 6.4% 和 156.0%,而液泡 Mn 含量降低 2.5%。添加外源 SNP 后,细胞壁和液泡 Mn 含量与 Cu 胁迫处理相比差异不显著,而细胞器 Mn 含量显著降低 46.2%。添加血红蛋白后,细胞壁和液泡 Mn 含量比 Cu+SNP 处理分别降低了 15.7% 和 20.5%,细胞器 Mn 含量提高 9.4%。
Cu 胁迫处理显著提高了番茄幼苗叶柄细胞中细胞壁和液泡的 Mn 含量,分别比对照提高 27.2% 和 66.8%,而细胞器的 Mn 含量显著降低 22.2%。与 Cu 胁迫处理相比,添加外源 SNP 后显著增加番茄幼苗根细胞中不同亚细胞结构的 Mn 含量,细胞壁、 细胞器和液泡 Mn 含量分别提高 18.0%、 2.7%和27.3%。添加血红蛋白后,细胞壁和液泡 Mn 含量比 Cu+SNP 处理分别降低了 41.3% 和 15.8%,细胞器 Mn 含量升高 12.3%。
Cu 胁迫下,番茄幼苗叶片细胞壁、 细胞器和液泡的 Mn 含量分别比对照提高 50.8%、 69.8%和2.8%。添加外源 SNP 后,液泡中 Mn 含量比 Cu 胁迫处理提高 11.4%,而细胞壁和细胞器 Mn 含量分别下降 43.2% 和 35.1%。添加血红蛋白后,细胞器的 Mn 含量占叶片细胞 Mn 总含量的比例与 Cu+SNP 处理相比提高了 6.3%,细胞壁和液泡的 Mn 含量所占比例分别下降了 6.0% 和 0.3%。
3 讨论与结论
铁(Fe)是生物体生命活动中必需的微量元素之一,其参与生物体内呼吸作用、 光合作用、 DNA 合成、 氮素同化和固定、 激素合成、 活性氧的形成与消除等重要的生理代谢过程[15]。植物正常生长发育不仅需要从环境中吸收充足的铁,更需要其在植物体内高效运转和在器官中合理分布以保证完成生命周期[16]。本试验中,铜(Cu) 胁迫使番茄幼苗各部位的 Fe 含量和累积量显著降低。这可能是由于过量 Cu2+存在竞争性抑制根系对 Fe2+的吸收,或者根系积累的大量 Cu2+限制了 Fe2+向地上部运输。添加外源 SNP 可显著增加番茄各部位的 Fe 含量,其中根系和叶柄中 Fe 含量和累积量最高。而添加血红蛋白可使 NO 的缓解效果消除。这可能是因为 SNP 释放出 NO,调控质膜上 ATPase 等功能蛋白活性,提高了 Cu 胁迫下番茄根系对 K、 Ca、 Fe、 Zn 等离子的吸收,使其在器官间的分配更加趋于合理,以维持胞质的离子稳态,从而缓解过多铜的毒害[17]。
研究表明,锌(Zn)在玉米细胞内的分布特征与其吸收和富集重金属能力密切相关[19]。细胞壁固持和液泡区隔化在植物对重金属的解毒、 耐性和超富集方面起着重要作用[20]。本试验中,Cu 胁迫使番茄幼苗根系 Zn 含量和累积量显著降低,叶片 Zn 含量和累积量显著提高。添加外源 SNP 可显著促进根系 Zn 的吸收和累积,提高叶片 Zn 含量和累积量。而添加血红蛋白可使 NO 的这种作用被削弱。说明正是由于 SNP 释放的 NO 作用于根系细胞质膜的磷脂双分子层,维持质膜正常的离子选择性,促进根系对 Zn 的吸收,缓解质膜受过多 Cu 的毒害[21-22]。另外,NO 作为信号分子参与抗逆信号转导,保护植物细胞免受逆境胁迫的损伤,提高根系和叶片细胞对 Zn 的吸收和富集[23]。
锌(Zn)在植物细胞的不同部位和细胞器中含量分布不一,胞内区室化分隔有利于保证缺锌和高锌胁迫下植物维持正常的生理代谢[24]。Ma等[25]研究显示,在天蓝遏蓝菜(Thlaspicaerulescens)的叶片中,77% 的锌主要储存在原生质体,其中 91% 的锌出现在液泡中。本试验中,锌主要集中在根系和茎的亚细胞结构中,少量锌存在于叶片和叶柄中。Cu 胁迫处理后,Zn 的亚细胞分布受到 Cu 胁迫处理的影响,根系各亚细胞结构和各部位细胞器中 Zn 含量显著降低,而叶片和叶柄细胞壁中 Zn 含量小幅提高,另有部分 Zn 含于茎和叶片的液泡中。添加 SNP 可显著增加各部位液泡、 细胞器的 Zn 含量,降低根系和叶片细胞壁 Zn 含量。表明外源 NO 能促进 Zn 在番茄体内的吸收和分布,并且影响 Zn 在细胞器和液泡内的积累,提高各部位细胞器对 Zn 的需求以及液泡中有机酸、 氨基酸和花青素等对过量 Zn 的储存[26]。
作为植物生命活动所必需的微量元素之一,锰(Mn)过量或缺乏,导致叶绿素合成减少、 光合作用受阻、 活性氧累积和体内激素失衡,导致生长受阻,尤其是叶部和根尖分生区的生长受到明显的抑制[27]。Xu等[28]利用同步辐射X-射线荧光光谱(synchrotron radiation X-ray fluorescence,SRXRF)微探针对商陆(PhytolaccaacinosaRoxb)根系、 茎和叶片中锰分布的分析结果表明,叶片是锰累积的主要器官,而根系、 茎和叶片的维管组织中存在高浓度的锰,有利于其长距离运输和向叶片不同组织中分配。根系主要通过有机酸的螯合作用促进植物对 Mn2+的转运解毒,同时能够将过量的 Mn2+分隔在细胞壁中; 叶片可通过酚类物质或有机酸螯合 Mn2+,并将其分隔在叶片表皮细胞和叶肉细胞的液泡中[29]。本试验中,Cu 胁迫使番茄幼苗叶柄和叶片的 Mn 含量和累积量显著提高,茎中 Mn 含量和累积量显著降低,根系 Mn 含量显著升高。添加 SNP 可显著增加根系和茎中 Mn 含量和累积量,降低叶柄和叶片中 Mn 含量和累积量。添加血红蛋白后 NO 的这种作用被减弱。表明铜胁迫下,外源 NO 可能介导了抗氧化酶保护系统,清除根尖细胞活性氧(ROS)引起的氧化损伤,保护细胞膜免受重金属胁迫的伤害,增强根系中有机酸(尤其是草酸、 柠檬酸)对过量 Mn2+的螯合,同时,促进叶柄和叶片中 Mn 向茎中转运,提高茎中 Mn 的补给,缓解 Cu 毒害[30-31]。
从番茄幼苗各部位及亚细胞中Fe、 Zn、 Mn含量和累积量的分布来看,同正常营养液处理相比,外源NO在Cu胁迫逆境下可以显著提高Fe、 Zn、 Mn的选择性吸收,调控番茄幼苗各部位及亚细胞中Fe、 Zn、 Mn的合理分布,维持胞质离子稳态和矿质营养元素平衡,缓解铜胁迫,保证番茄幼苗正常的生理代谢。
参考文献:
[1] 王晓兰, 周守标, 杨集辉, 等. 铜胁迫对弯囊苔草(Carexdispalata)生长和生理特性的影响[J]. 农业环境科学学报, 2010, 29(2): 264-269.
Wang X L, Zhou S B, Yang J Hetal. Copper stress on the growth and some physiological parameters ofCarexdispalata[J]. Journal of Agro-Environment Science, 2010, 29(2): 264-269.
[2] 谢建春, 赵娟, 杨世勇. 铜尾矿对油菜生长和生理功能的影响[J]. 生态与农村环境学报, 2009, 25(2): 74-79.
Xie J C, Zhao J, Yang S Y. Effects of copper mine tailings on growth and physiological functions ofBrassicacampestris[J]. Journal of Ecology and Rural Environment, 2009, 25(2): 74-79.
[3] Sharma S S, Dietz K J. The relationship between metal toxicity and cellular redox imbalance[J]. Trends in Plant Science, 2009, 14(1): 43-50.
[4] Wojcik M, Tukiendorf A. Response of wild type ofArabidopsisthalianato copper stress[J]. Biologia Plantarum, 2003, 46(1): 79-84.
[5] 高晓宁. 土壤重金属污染现状及修复技术研究进展[J]. 现代农业科技, 2013, (9): 229-231.
Gao X N. Research advances on soil heavy metal pollution and remediation technology[J]. Modern Agricultural Science and Technology, 2013, (9): 229-231.
[6] Beligni M V, Lamattina L. Nitric oxide: a non-traditional regulator of plant growth[J]. Trends in Plant Science, 2001, 6(11): 508-509.
[7] Tan Y W, Xu Y F, Zhou H. Effects of nitric oxide on seed germination and seedling growth of tall fescue under salt stress[J]. Acta Agrestia Sinica, 2010, 18(3): 394-398.
[8] 张敏, 姜春辉, 李晓云, 崔秀敏. 外源 NO 对铜胁迫下番茄幼苗根系构型及其超微结构的影响[J]. 植物生理学报, 2012, 48(11): 1105-1112.
Zhang M, Jiang C H, Li X Y, Cui X M. Effects of exogenous nitric oxide on tomato seedlings root architecture and ultrastructure under copper stress[J]. Plant Physiology Journal, 2012, 48(11): 1105-1112.
[9] 张敏, 姜春辉, 崔秀敏. 外源 NO 供体硝普钠(SNP)对铜胁迫下番茄幼苗生理生化指标的影响[J]. 植物生理学报, 2013, 49(2): 144-152.
Zhang M, Jiang C H, Cui X M. Effects of exogenous nitric oxide donor SNP on physiological and biochemical indexes in tomato seedlings under copper stress[J]. Plant Physiology Journal, 2013, 49(2): 144-152.
[10] 李兴华, 方子森, 牛俊义. 大量及微量元素对胡麻幼苗生长发育的影响[J]. 甘肃农业大学学报, 2013, 48(1): 42-48.
Li X H, Fang Z S, Niu J Y. Effects of major and trace elements on growth and development of flax seedlings[J]. Journal of Gansu Agricultural University, 2013, 48(1): 42-48.
[11] 姜春辉, 王秀峰, 尹博, 等. 外源 NO 对 Cu 胁迫下番茄 Cu 的亚细胞分布和化学形态的影响[J]. 应用生态学报, 2012, 23(11): 3033-3039.
Jang C H, Wang X F, Yin Betal. Effects of exogenous nitric oxide on the subcellular distribution and chemical forms of copper in tomato seedlings under copper stress[J]. Chinese Journal of Applied Ecology, 2012, 23(11): 3033-3039.
[12] Weigel H J, Jäger H J. Subcellular distribution and chemical form of cadmium in bean plant[J]. Plant Physiology, 1980, 65: 480-482.
[13] Rathore V S, Baiai Y P, Wittwer S H. Subcellular localization of zinc and calcium in bean (PhaseolusvulgarisL.) tissues[J]. Plant Physiology, 1972, 49: 207-211.
[14] 段曼莉, 付冬冬, 王松山, 等. 亚硒酸盐对四种蔬菜生长、 吸收及转运硒的影响[J]. 环境科学学报, 2011, 31(3): 658-665.
Duan M L, Fu D D, Wang S Setal. Effects of different selenite concentrations on plant growth, absorption and transportation of selenium in four different vegetables[J]. Acta Scientiae Circumstantiae, 2011, 31(3): 658-665.
[15] 申红芸, 熊宏春, 郭笑彤, 左元梅. 植物吸收和转运铁的分子生理机制研究进展[J]. 植物营养与肥料学报, 2011, 17(6): 1522-1530.
Sheng H Y, Xiong H C, Guo X T, Zuo Y M. Progress of molecular and physiological mechanism of iron uptake and translocation in plants[J]. Plant Nutrition and Fertilizer Science, 2011, 17(6): 1522-1530.
[16] Morrissey J, Guerinot M L. Iron uptake and transport in plants: the good, the bad, and the ionome[J]. Chemical Reviews, 2009, 109(10): 4553-4567.
[17] Zhang Y K, Han X J, Chen X Letal.Exogenous nitric oxide on antioxidative system and ATPase activities from tomato seedlings under copper[J]. Scientia Horticulturae, 2009, 123(2): 217-223.
[18] Graziano M, Beligni M V, Lamattina L. Nitric oxide improves internal iron availability in plants[J]. Plant Physiology, 2002, 130: 1852-1859.
[19] 周建民, 党志, 陶雪琴,周永章. NTA 对玉米体内 Cu、 Zn 的积累及亚细胞分布的影响[J]. 环境科学, 2005, 26(6): 126-130.
Zhou J M, Dang Z, Tao X Q, Zhou Y Z. Influence of NTA on accumulation and subcellular distribution of copper and zinc in corn (Zeamays)[J]. Environmental Science, 2005, 26(6): 126-130.
[20] Küpper H, Lombi E, Zhao F Jetal. Cellular compartmentation of cadmium and zinc in relation to other elements in the hyperaccumulatorArabidopsishalleri[J]. Planta, 2000, 212(1): 75-84.
[21] 张远兵, 刘爱荣, 方蓉. 外源一氧化氮对镉胁迫下黑麦草生长和抗氧化酶活性的影响[J]. 草业学报, 2008, 17(4): 57-64.
Zhang Y B, Liu A R, Fang R. Effect of exogenous nitric oxide on the growth and antioxidant enzyme activities of Lolium perenne under Cd stress[J]. Acta Prataculturae Sinica, 2008, 17(4): 57-64.
[22] 张义凯, 韩小娇, 金洪, 等. 外源 NO 对铜胁迫下番茄光合、 生物发光特性及矿质元素吸收的影响[J]. 植物营养与肥料学报, 2010, 16(1): 172-178.
Zhang Y K, Han X J, Jin Hetal. Effects of exogenous nitric oxide on photosynthetic, bioluminescent characteristics and mineral element contents in tomato under copper stress[J]. Plant Nutrition and Fertilizer Science, 2010, 16(1): 172-178.
[23] 彭金英, 黄勇平. 植物防御反应的两种信号转导途径及其相互作用[J]. 植物生理与分子生物学报, 2005, 31(4): 347-353.
Peng J Y, Huang Y P. The signaling pathways of plant defense response and their interaction[J]. Journal of Plant Physiology and Molecular Biology, 2005, 31(4): 347-353.
[24] Broadley R, White P J, Hammond J Petal. Zinc in plants[J]. New Phytologist, 2007, 173(4): 677-702.
[25] Ma J F, Ueno D, Zhao F Jetal. Subcellular localization of Cd and Zn in the leaves of a Cd-hyperaccumulating ecotype ofThlaspicaerulescens[J]. Planta, 2005, 220(5): 731-736.
[26] Cosio C, Martinoia E, Keller C. Hyperaccumulation of cadmium and zinc inThlaspicaerulescensandArabidopsishalleriat the leaf cellular level[J]. Plant Physiology, 2004, 134(2): 716-725.
[27] Santandrea G, Schiff S, Bennici A. Effects of manganese on Nicotiana species cultivated in vitro and characterization of regenerated Mn-tolerant tobacco plants[J]. Plant Science, 1998, 132(1): 71-82.
[28] Xu X H, Shi J Y, Chen Y Xetal. Distribution and mobility of manganese in the hyperaccumulator plantPhytolaccaacinosaRoxb. (Phytolaccaceae)[J]. Plant and Soil, 2006, 285(1-2): 323-331.
[29] 张玉秀, 李林峰, 柴团耀, 等. 锰对植物毒害及植物耐锰机理研究进展[J]. 植物学通报, 2010, 45(4): 506-520.
Zhang Y X, Li L F, Chai T Yetal. Mechanisms of manganese toxicity and manganese tolerance in plants[J]. Chinese Bulletin of Botany, 2010, 45(4): 506-520.
[30] Mora M L, Rosas A, Ribera Aetal. Differential tolerance to Mn toxicity in perennial ryegrass genotypes: involvement of antioxidative enzymes and root exudation of carboxylates[J]. Plant and Soil, 2009, 32(1-2): 79-89.
[31] 赵宝泉, 万宇, 杨世湖, 等. 外源 NO 供体硝普钠(SNP)对重金属 Cd 胁迫下水稻幼苗膜脂过氧化及抗氧化酶的影响[J]. 江苏农业学报, 2010, 26(3): 468-475.
Zhao B Q, Wan Y, Yang S Hetal. Effects of exogenous nitric oxide donor SNP on membrane lipid peroxidation and activity of antioxidative enzyme in rice seedlings under Cd stress[J]. Jiangsu Journal of Agricultural Sciences, 2010, 26(3): 468-475.
[32] 徐向华, 施积炎, 陈新才, 等. 锰在商陆叶片的细胞分布及化学形态分析[J]. 农业环境科学学报, 2008, 27(2): 515-520.
Xu X H, Shi J Y, Chen X Cetal. Subcellular distribution and chemical fractions of manganese in leaves of hyper accumulator phytolacca acinosa roxb[J]. Journal of Agro-Environment Science, 2008, 27(2): 515-520.
[33] 倪才英, 刘永厚, 赵振纪. 赣东北土壤中铜的毒害作用研究[J]. 江西师范大学学报(自然科学版), 1999, 23(2): 170-173.
Ni C Y, Liu Y H, Zhao Z J. A study on toxicity of copper in soil of northeast Jiangxi province[J]. Journal of Jiangxi Normal University(Natural Sciences Edition), 1999, 23(2): 170-173.
