紫扇贝、海湾扇贝及其正反杂交子代群体遗传结构的AFLP分析
2013-10-13黄晓婷胡丽萍王春德包振民
孙 妍, 黄晓婷, 胡丽萍, 王 师, 王春德, 包振民
(1.海洋生物遗传学与育种教育部重点实验室, 中国海洋大学 海洋生命学院, 山东 青岛 266003; 2.青岛农业大学 海洋科学与工程学院, 山东 青岛266109)
海湾扇贝(Argopecten irradians, Lamarck, 1819)原产于北美洲大西洋沿岸, 于 1982年引入中国[1],由于其生长迅速而受到养殖户欢迎, 目前已成为中国重要的海水养殖贝类之一[2-3]。但是, 由于引种个体数目较少, 随着养殖规模的不断扩大, 遗传多样性降低, 近交衰退现象显著, 种质资源的退化需要引种复壮和遗传育种工作的不断进行, 对产业的可持续发展造成了严重威胁[4-5]。与海湾扇贝同属的紫扇贝(A. purpuratus,Lamarck, 1819), 是原产于南太平洋的速生型中型扇贝, 主要在智力北部和秘鲁沿岸进行规模化人工养殖[6], 具有体型大、生长速度快、寿命长等优点。
杂交在改善海洋贝类种质资源中有许多成功的应用[3,7], 通过杂交育种的方式, 有望丰富现有扇贝种质资源并产生优于亲本生产性状的杂种优势。青岛农业大学王春德教授于2008年将紫扇贝引入中国,并开展了紫扇贝与海湾扇贝的杂交育种工作, 获得了杂交子一代成体[8], 在生长率、成活率上优于亲本,与海湾扇贝相比, 杂交子代成贝总质量增加125.9%~138.9%, 肉柱重增加 145.4%~156.2%, 具有明显的杂种优势[9]。在紫扇贝和海湾扇贝的杂交中,亲本双方在杂交育种中的贡献率大小、种间杂交子代与种内交配子代遗传上的差异、以及杂种优势的遗传基础等问题都需要我们对杂种及其亲本的遗传结构进行分析。
本研究应用AFLP分子标记技术, 对紫扇贝、海湾扇贝、正交子一代(紫扇贝♀×海湾扇贝♂)及反交子一代(海湾扇贝♀×紫扇贝♂)群体进行了遗传结构分析, 从分子水平探讨杂交子代的遗传组成, 同时为杂种优势机理的阐述提供理论依据。
1 材料和方法
1.1 材料
本实验所用紫扇贝(PP)、海湾扇贝(II)及其正反杂交后代(紫扇贝♀×海湾扇贝♂, PI; 海湾扇贝♀×紫扇贝♂, IP)均由青岛农业大学王春德教授提供,其中紫扇贝为秘鲁引进种, 海湾扇贝取自青岛胶南市, 杂交实验及杂种培育参见Wang等[9]的方法。
1.2 方法
基因组 DNA的提取采用常规酚-氯仿抽提法提取基因组DNA[10]。AFLP分析参照Vos等[11]的方法,引物及接头序列参照潘洁等[12], 由上海生物工程合成, 序列见表1。选择性扩增用4种EcoR I引物和3种Mse I引物, 共5对组合引物组合: E-AAC/M- CAG、E-AAC/M-CCA、E-AAG/M-CCA、E-ACA/M- CTG、E-ATC/M-CTG。扩增产物经4.5%聚丙烯酰胺凝胶电泳后, 进行银染显带。

表1 AFLP分析中所用的接头及引物序列Tab. 1 The adapter and primer sequence used in AFLP analysis
数据统计与分析银染之后得到的带谱应用Cross checker 2.8软件进行统计分析, 有带记为1, 无带记为 0, 获得 1、0矩阵。统计各群体扩增位点数和多态位点数。用Pop Gene32软件计算群体内的多态位点比率、Nei’ s基因多样性、香农氏多样性指数和群体间遗传相似度及遗传距离。为评估遗传变异来源于群体内还是群体间, 用Arlequin3.11进行分子方差分析(Analysis of Molecularof Variance, AMOVA), 计算群体间变异来源和遗传分化指数。
2 实验结果
2.1 AFLP 扩增带谱分析
本研究用 E-AAC/M-CAG、E-AAC/M-CCA、E-AAG/M-CCA、E-ACA/M-CTG、E-ATC/M-CTG五对引物组合, 在紫扇贝(PP)、海湾扇贝(II)及其正反杂交子代(PI和IP)4个群体中共得到了433个位点,平均每对引物得到86.6个位点。
AFLP在亲本和杂交后代 4个群体中的扩增带谱可以分为4种, 如图1: (I)双亲和正反杂交子代均有的扩增位点, 这种位点共有 169个, 占总位点数的 39.03%; (II)亲本一方与杂交子代共享位点,其中, 紫扇贝与正反交子代共享的位点数为 82个,海湾扇贝与杂交子代共享的位点数为 181个; (III)在亲本中出现, 而杂交子代中丢失的位点, 统计发现, 紫扇贝和海湾扇贝各有30和18个位点在子代中丢失; (Ⅳ)非亲位点, 即亲本中没有, 而杂交子代中出现的位点, 这类位点共有 12个(正交子代中5个、反交子代中 8个)。由此可见, 正反交子代97.23%扩增得到的位点来源于紫扇贝和海湾扇贝,且来源于海湾扇贝的位点数多于来源于紫扇贝的位点数。本研究在分子水平上证实了杂交扇贝成贝的杂种身份。

图1 紫扇贝、海湾扇贝及杂交子代AFLP选择性扩增结果—E33M54Fig. 1 Polyacrylamide gel electrophoresis result of selective amplification
2.2 紫扇贝(PP)、海湾扇贝(II)及其正反杂交子一代(PI和IP)的群体遗传结构分析
4个群体的多态位点比率、Nei’ s基因多样性、香农氏指数见表 2。4个群体中, 紫扇贝(PP)群体的遗传多样性最低; 海湾扇贝(II)群体最高。杂交子代群体的遗传多样性介于两亲本之间, 且反交群体高于正交群体。紫扇贝群体内遗传多样性较低可能是由于目前引种数目较少引起的。

表2 4个群体内遗传多样性参数Tab. 2 Genetic diversityparameters withinfour groups
从群体间的遗传距离(表 3)可以看出, 两个亲本(PP和II)间的遗传距离为0.5464, 正反杂交F1代与两亲本的遗传距离为 0.2186~0.2701, 表明两个亲本间、子代与亲本间的遗传差异较大。而正反交F1代间遗传距离小, 遗传相似度高, 表明正反交子代无群体分化。此外, 正反杂交F1代与母本的遗传距离均比与父本的遗传距离小, 表明杂交子代一定程度偏向母本, 从母本获得的遗传信息可能要多于从父本获得的遗传信息。
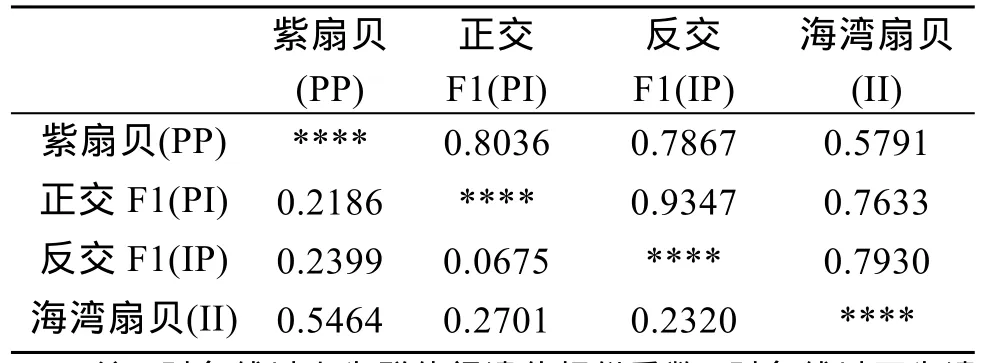
表3 4个群体间的遗传相似性和遗传距离Tab. 3 Genetic distances and similarities between four groups
应用分子方差分析(AMOVA)对4个群体两两间的遗传多样性来源进行分析(表 4), 两亲本群体间遗传差异最大, 两亲本间和亲本与杂交子代间遗传分化显著; 正反杂交子代间遗传差异最小, 没有明显的遗传分化。
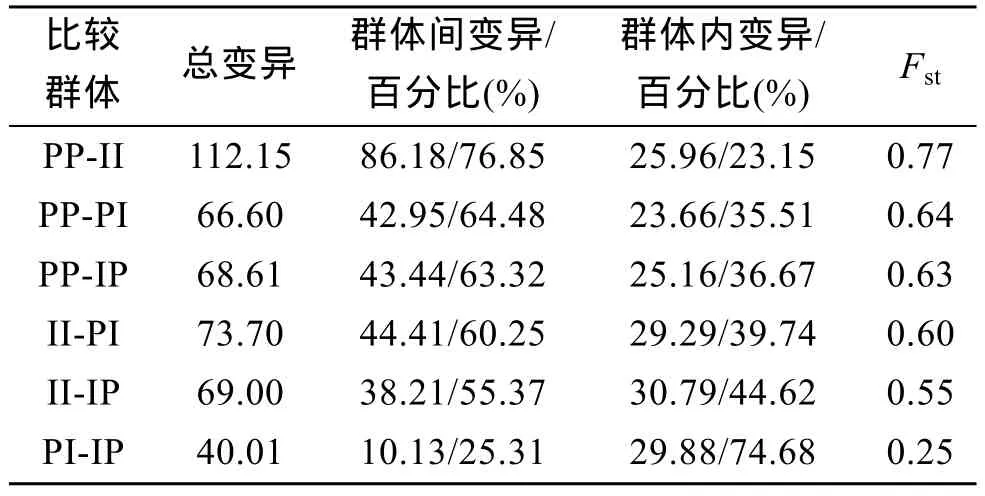
表4 4个群体总遗传变异来源及遗传分化指数Tab. 4 The source of variation and Fstabout four scallop groups
3 讨论
紫扇贝与海湾扇贝同属于海湾扇贝属(Argopecten), 进化关系较近, 是当今世界上两种重要的海水经济养殖扇贝[13]。虽然紫扇贝在生产上做出了相当大的贡献, 但针对其分子生物学的研究很少, 基因组信息相对于其他经济养殖扇贝而言十分有限。AFLP标记对基因组的信息背景没有要求, 通过改变限制性内切酶和选择性碱基的种类与数目,可产生大量的理论上可以覆盖整个基因组标记数目,该标记还具有丰富的多态性, 对遗传关系相近的材料也可达到较强的多态分辨能力。本研究选用AFLP分子标记技术, 能够从基因组的水平对紫扇贝、海湾扇贝及其正反杂交F1代4个群体之间的遗传关系进行分析, 从而确证杂交子代的遗传组成, 并为研究杂种优势产生提供可能的遗传基础。
杂交作为一种改良生产性状的重要途径, 在海洋养殖扇贝的育种工作已有很多报道。扇贝种内不同地理种群之间杂交的研究较多也较为成熟, 常亚青和刘小林对栉孔扇贝中国种群与日本种群及与俄罗斯群体的杂交组合的研究表明, 杂交后代表现出了不同程度的杂交优势[14-15]。种间杂交方面, 国内在栉孔扇贝(Chlamys farreri)、海湾扇贝(A. irradians irradians)、虾夷扇贝(Patinopecten yessoensis)和华贵栉孔扇贝(Mimachlamys nobilis)等不同种间杂交已经开展了很多工作[16-18]。但是, 种间杂交由于亲本间遗传差异较大, 常出现杂交不亲和、幼体不成活等“远缘杂交综合征”, 导致在许多贝类的种间杂交中出现低幼虫孵化率、幼虫低成活率和高胚胎畸形率的现象[17,19]。仅在杨爱国等开展的栉孔扇贝与虾夷扇贝杂交研究中, 获得的杂交子一代, 并在抗逆性能方面明显优于亲贝, 而通过细胞遗传学分析发现, 该两种扇贝的杂种身份在幼虫时期是可以证实的, 在幼虫发育过程中染色体组成上发生了一定的变异,成体时期的杂种身份仍需验证[20-21]。王春德等[8-9]于2007和2008年将紫扇贝引进中国, 并开展了紫扇贝和海湾扇贝种间杂交研究, 成功获得了杂交扇贝成体, 且杂交扇贝表现出一定的杂种优势, 这在已有的扇贝杂交育种研究当中是鲜有报道的, 借助细胞遗传学和分子生物学等技术手段, 作者已经证明杂种子代的确由双亲的遗传物质组成, 证明的杂交一代的杂种身份[22]。
为了进一步了解杂种子代的遗传组成, 本研究应用 AFLP技术对紫扇贝、海湾扇贝及正反杂交子代的遗传结构进行了分析, 计算出两亲本遗传距离0.5464, 遗传分化指数Fst为0.77。Nei[23]估计亚种间的遗传距离值约为 0.02~0.20; 种间遗传距离的变幅为 0.10~2.00, Wright[24]提出,Fst>0.25 说明群体间的遗传分化很大。由此可见, 本研究中两亲本扇贝群体遗传分化显著, 属于两个不同的物种, 它们的杂交属于种间杂交。但是扇贝种间杂交子代, 尤其是杂交成贝, 可能有复杂的遗传组成。本研究对杂交子代遗传结构的分析表明, 正反交子代中的位点大部分来源于紫扇贝和海湾扇贝, 来源于海湾扇贝的扩增位点略多于来源于紫扇贝的扩增位点, 原因可能是海湾扇贝群体的遗传多样性高于紫扇贝; 此外,正反交F1与双亲的Fst指数都>0.25, 表明分化显著,且F1与双亲的遗传相似度和遗传距离显示其为介于双亲间的中间种; 虽然杂交子代与紫扇贝和海湾扇贝的遗传关系上并不完全对等, 略偏向于母本一方,但并不表现出明显的倾向性。本研究结果为紫扇贝和海湾扇贝种间杂交子代的遗传组成提供了分子水平的证据。
紫扇贝和海湾扇贝杂交获得的子代, 与海湾扇贝相比, 杂交子代成贝总质量增加了 125.9%~138.9%,肉柱质量增加了 145.4%~156.2%, 具有明显的杂种优势[9]。杂种优势作为生物界普遍存在的现象, 在改良生产性状和种质品质中得到了广泛的应用, 但其理论基础的研究却相对薄弱, 理论研究的滞后也制约其在生产中的进一步应用。因此, 杂种优势的遗传机理研究一直是近百年来遗传和育种学家研究的热点和难点。早在 20世纪初, Bruce[25]和 Shull[26]就基于单基因遗传效应相继提出了杂种优势的显性假说和超显性假说。随着研究手段的改进和研究内容的深入, 认为杂种优势的遗传实质应是各种基因效应的综合,特别是上位效应和连锁效应在杂种优势遗传基础中将占有重要地位。杂种优势对包括贝类在内的海洋经济动物进行遗传改良研究和应用的历史较短, 关于其杂种优势遗传机理的研究报道也很少[27]。AFLP作为一种显性标记应用于紫海杂交贝的遗传结构分析, 可以为海洋贝类杂种优势产生的机理的探讨提供一定的线索。在杂交过程中子代出现了非亲遗传的位点, 这些位点的产生可能是由于亲本双方的同源染色体相互作用对酶切位点引起的修饰或隐蔽, 或者个别酶切位点附近碱基突变等, 这些杂交子代中出现的种种非孟德尔遗传变化可能与其在生产上表现出的杂种优势有一定关系, 需要进一步深入研究加以证实。
[1]张福绥, 何义朝, 刘祥生, 等. 海湾扇贝(Argopecten irradians)引种、育苗及试养[J].海洋与湖沼, 1986,17(5): 367-374.
[2]张福绥, 何义朝, 杨红生. 海湾扇贝引种工程及其综合效应[J]. 中国工程科学, 2000, 2(2): 30-35.
[3]Guo X. Use and exchange of genetic resources in molluscan aquaculture [J]. Reviews in Aquaculture,2009, 1(3-4): 251-259.
[4]张国范, 刘述锡, 刘晓, 等. 海湾扇贝自交家系的建立和自交效应[J]. 中国水产科学, 2003, 10(6):441-445.
[5]Zheng H, Zhang G, Liu X, et al.Inbreeding depression for various traits in two cultured populations of the American bay scallop,Argopecten irradians irradiansLamarek (1819)introduced into China[J].Journal of Experimental Marine Biology and Ecology,2008,364(1):42-47.
[6]Brand E, Merino G E, Abarca A, et al. Scallop fishery and aquaculture in Chile [J]. Developments in Aquaculture and Fisheries Science, 2006, 35: 1293-1314.
[7]燕敬平, 孙慧玲, 方建光, 等. 日本盘鲍与皱纹盘鲍杂交育种技术研究[J]. 海洋水产研究, 1999, 20(1):35-39.
[8]王春德, 刘保忠, 李继强, 等. 紫扇贝与海湾扇贝种间杂交的研究[J]. 海洋科学, 2009, 33(10): 84-87.
[9]Wang C, Liu B, Li J, et al. Introduction of the Peruvian scallop and its hybridization with the bay scallop in China[J]. Aquaculture, 2011, 310(3): 380-387.
[10]Sambrook J, Russell David W. Molecular cloning: a laboratory manual. Vol. 3[M]. Cold Spring Harbor Laboratory Press, 1989.
[11]Vos P, Hogers R, Bleeker M, et al. AFLP: a new technique for DNA fingerprinting[J]. Nucleic acids research, 1995, 23(21): 4407-4414.
[12]潘洁, 包振民, 赵洋, 等. 扇贝 AFLP分析中的引物选择[J]. 青岛海洋大学学报, 2002, 32(Sup.): 310.
[13]Gonzalez M L, Perez M C, Lopez D A. Breeding cycle of the northern scallop,Argopecten purpuratus(Lamarck,1819)in southern Chile [J]. Aquaculture Research, 2002,33(11): 847-852.
[14]常亚青, 刘小林, 相建海, 等.栉孔扇贝中国种群与日本种群杂交子一代的早期生长发育[J]. 水产学报,2002, 26(5): 385-388.
[15]刘小林, 常亚青, 相建海, 等. 栉孔扇贝不同种群杂交效果的初步研究.I中国种群与俄罗斯种群的杂交[J]. 海洋学报, 2003, 25(1): 93-99.
[16]张玉勇, 常亚青, 宋坚. 杂交育种技术在海水养殖贝类中的应用及研究进展[J]. 水产科学, 2005, 24(4):39-41.
[17]刘宪杰, 常亚青. 扇贝种间杂交的可行性及幼体早期发育的初步研究[J]. 大连水产学院学报, 2006, 4: 35-39.
[18]刘广绪, 包振民, 胡景杰, 等. 栉孔扇贝、华贵栉孔扇贝及其种间杂交子代、种内交配子代的 ISSR分析[J]. 中国海洋大学学报, 2006, 36(1): 71-75.
[19]张国范, 刘晓. 关于贝类遗传改良几个问题的讨论[J].水产学报, 2006, 30(1): 130-137.
[20]吕振明, 杨爱国, 王清印, 等. 栉孔扇贝和虾夷扇贝杂交子代的细胞遗传学研究[J].高技术通讯, 2006,16(8): 853-858.
[21]吕振明, 杨爱国, 王清印, 等. 栉孔扇贝和虾夷扇贝杂交子代的GISH鉴定及其免疫学特性[J]. 中国水产科学, 2006, 13(4): 587-602.
[22]Hu L, Huang X, Mao J, et al. Genomic Characterization of Interspecific Hybrids between the ScallopsArgopecten purpuratusandA. irradians irradians[J]. PLOS ONE, 2013, 8(4): e62432.
[23]郭平仲. 种群间遗传距离与进化趋异[J]. 北京师范学院学报. 1992, 13(2): 63-71.
[24]Weight S. Evolution and the Genetics of Population Variability Within and Among Natural Population[M].Chicago: University of Chicago Press, 1978.
[25]Shull G H. The composition of a field of maize[J].Journal of Heredity, 1908(1): 296-301.
[26]兰进好, 张宝石, 周鸿飞. 作物杂种优势遗传基础研究进展[J]. 中国农学通报2005, 21(1): 114-119.
[27]张国范, 刘晓, 阙华勇, 等. 贝类杂交及杂种优势理论和技术研究进展[J]. 海洋科学2004, 28(7): 54-60.
