杂交大豆光合特性与产量的关系
2013-09-15韩俊梅卫保国张海平
韩俊梅,卫保国,张海平
(1.山西大学生物工程学院,山西太原030006;2.山西省农业科学院农作物品种资源研究所,山西太原030031)
农业生产上,利用杂种优势对提高农作物产量有重要意义[1]。目前,杂种优势在水稻、玉米、棉花、高粱、蔬菜等作物上已广泛应用,大豆、小麦正在进入应用阶段[2-3]。王志新等提出大豆杂交种具有明显的杂种优势,可显著提高大豆的产量。王艳玲等[4]通过对不同组合F1的杂种优势分析表明,大豆部分形态性状、产量性状及品质性状具有较高的杂种优势。作物干物质的90%~95%来自作物的光合作用,光合作用是作物产量形成的基础。而有研究表明,光合速率与产量存在极显著的正相关关系[5]。杂交种在光合性能上存在多种形式的杂种优势。一般情况下,杂交种具有明显的超亲优势[6-7]。赵明等[7]研究发现,杂交种较其亲本在光合速率上表现为正优势,幅度在3.1%~23.4%之间,超高亲2.4%~9.3%,光合速率杂种优势与叶绿素含量有关。但也有报道研究表明,杂交种在光合速率上表现为无优势或负优势现象[8]。随着杂交大豆的问世,杂交大豆育种家逐渐开始关注杂交大豆的光合速率等相关生理性状的研究。近年来,国内外对作物光合速率、蒸腾速率和水分利用效率的研究十分活跃,主要集中在小麦、玉米等禾本科作物上[9-11]。对杂交大豆产量杂种优势形成的机理研究较少。
本研究以大豆的杂交种及其亲本为材料,对其苗期、盛花期、结荚期、鼓粒期、成熟期的净光合速率、蒸腾速率、水分利用率、气孔导度、胞间CO2浓度进行测定,以探讨大豆杂交种光合生理指标的变化规律,为大豆杂交种的优势组合选配提供理论依据。通过研究杂交种与其亲本的不育系、恢复系、保持系的产量与净光合速率的相关性,阐明其光合作用与产量的关系,同时从光合性能探讨其杂种优势的生理变化规律,为强优势组合选配和高产栽培提供理论依据。
1 材料和方法
1.1 试验材料
供试材料由山西省农业科学院农作物品种资源研究所卫保国研究员、张海平博士提供。用已培育出的杂交种SXNJCMS1A×中-21、恢复系中-21、不育系SXNJCMS1A、保持系SXNJCMS1B为试材。杂交种SXNJCMS1A×中-21经过2011,2012年的山西晋中和晋城高平、陕西延安、甘肃庆阳等多点试验,比对照品种晋豆19号增产15.20%~28.89%。
1.2 试验设计
于2011年进行预备试验,2012年在榆次东阳试验基地进行正式试验。每个组合按杂交种、恢复系、不育系、保持系的顺序进行种植,小区面积12 m2(6 m×2 m),各品种4行区,行距为50 cm,3次重复。生育期间进行常规的田间管理。成熟后按小区进行测产,包括生物学产量和经济学产量。
1.3 测定项目与方法
分别在苗期、盛花期、结荚期、鼓粒期、成熟期对每个品种的各个重复随机抽取6株,于晴天9:00—11:00采用 CB-1102便携式光合蒸腾仪测定其倒4复叶的中间叶片主要的光合指标(净光合速率、蒸腾速率、气孔导度、胞间CO2浓度),并采用Fischer and Tumer方法计算水分利用率。
2 结果与分析
2.1 不同生育时期光合指标的变化
2.1.1 不同生育时期净光合速率的变化 由图1可知,苗期杂交种及其亲本三系净光合速率都比较低,各品种之间差异不明显。随着植株的生长,净光合速率也逐渐增加,在鼓粒期达到最大值,鼓粒之后急剧下降。光合速率的整体变化规律与杨文杰等[12]的研究结果相似。在盛花期、结荚期、鼓粒期,杂交种的净光合速率均高于其亲本三系,但结荚期和鼓粒期的净光合速率变化不大,可能是由于杂交种在结荚期后植株高大,植株之间相互遮盖,叶片得不到足够光照。结荚期杂交种的净光合速率显著高于其亲本三系(P<0.05),即在生殖生长时期,杂交种保持较高的光合能力和积累更多的有机产物,使有机产物能源源不断地运往生殖器官,使产量增加。

不育系在整个生育期内净光合速率变化平稳,在盛花期生殖生长开始,净光合速率高有利于生殖生长。但由于大部分花朵没有授粉,花荚脱落,使大豆没有足够的库,源库比过大,降低维管束通畅程度,增加同化物运输的阻力,减慢同化物的运输速度,同化物大量积聚在叶片中,则会大大降低叶片的同化能力[13-14],使得不育系结荚期和鼓粒期光合速率不高。
2.1.2 不同生育时期蒸腾速率的变化 由图2可知,从苗期到鼓粒期,随着光合能力的增强,蒸腾速率有不同程度的增强。杂交种在结荚期达到最大值,且与其亲本三系差异显著,在鼓粒期有所下降。而恢复系、不育系、保持系的变化与净光合速率的变化趋势一致。杂交种结荚期蒸腾速率高,能给水分、营养物质的运输提供足够的动力,保证了流的畅通。

2.1.3 不同生育时期水分利用率的变化 由图3可知,结荚期和鼓粒期杂交种的水分利用效率高于其亲本三系,特别是结荚期呈显著水平(P<0.05)。从图3可以看出,从营养生长到生殖生长水分利用率不断升高。杂交种在结荚期水分利用率高,说明此时期水分是影响光合作用的重要因素,同时保证了其有机物质的运输。因此,此时期充足的水分供应对于产量形成极为重要,这与一些学者的研究结论一致[15-16]。

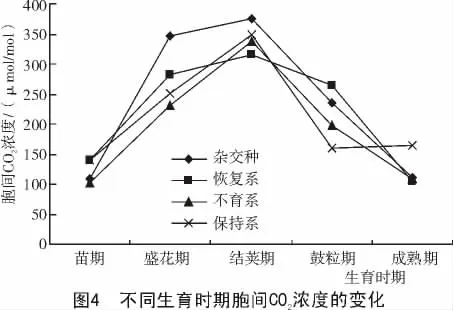
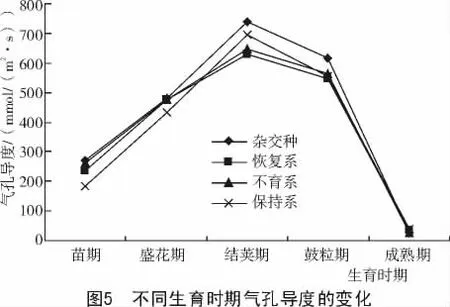
2.1.4 不同生育时期胞间CO2浓度的变化 由图4可知,在整个生育期内,从苗期开始胞间CO2浓度逐渐升高,到结荚期达到最大值,与净光合速率的变化规律一致。而杂交种在盛花期和结荚期均高于其亲本三系,足够的CO2供应使得光合作用有充足的原料供给,提高了光合作用,为籽粒提供了足够的营养物质。在大豆生育后期鼓粒期,叶片互相遮盖,空气不流通,可利用的CO2浓度低,成为光合作用的限制因子,增加CO2浓度有利于增加产量。2.1.5 不同生育时期气孔导度的变化 由图5可知,在大豆的各个生育期,气孔导度的变化趋势与光合速率的变化趋势基本一致,即净光合速率高,则气孔导度也大。杂交种在整个生育期内均高于其亲本三系,气孔是控制叶片内外水蒸汽和CO2扩散的门户,影响着蒸腾与光合进程。结荚期光合速率高,CO2供应充足,气孔导度也大,到鼓粒期由于植株繁茂,植株之间的空间减少,CO2浓度降低,气孔导度随之降低。鼓粒后期随着植株趋于成熟,光合速率下降,气孔导度也随之下降。
2.2 结荚期主要光合性能参数之间的相关分析
大豆的产量与光合作用特别是结荚期间的CO2同化作用有密切关系。Wells等[17]认为,大豆结荚期和生理成熟期间的表观冠层光合作用曲线与种子产量呈正相关。对5个生育期间各个参数的比较和分析发现,杂交种在结荚期均高于其亲本三系,所以,对结荚期光合性能指标之间进行相关分析,以探明影响光合作用的重要因素。
由表1可知,结荚期净光合速率与蒸腾速率、气孔导度、胞间CO2浓度均呈正相关,而与水分利用率呈极显著正相关,说明结荚期水分是影响光合速率的重要因素。净光合速率与气孔导度的相关系数达到0.722,而气孔张开越大,胞间CO2浓度越高,有利于提供足够的光合原料,从而提高光合速率。
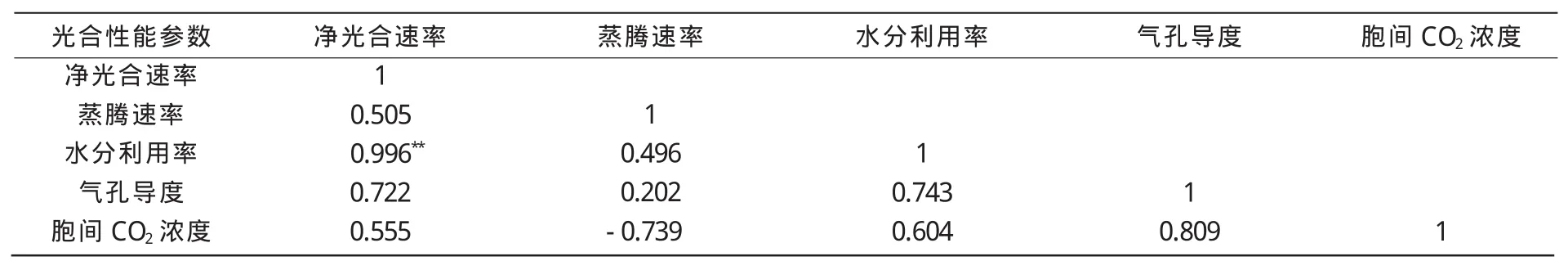
表1 结荚期主要光合性能参数之间的相关分析
2.3 光合速率与产量的关系
方差分析结果(表2)表明,参试杂交大豆同其亲本相比具有明显的产量优势,二者之间差异达极显著水平,F值为21.413。
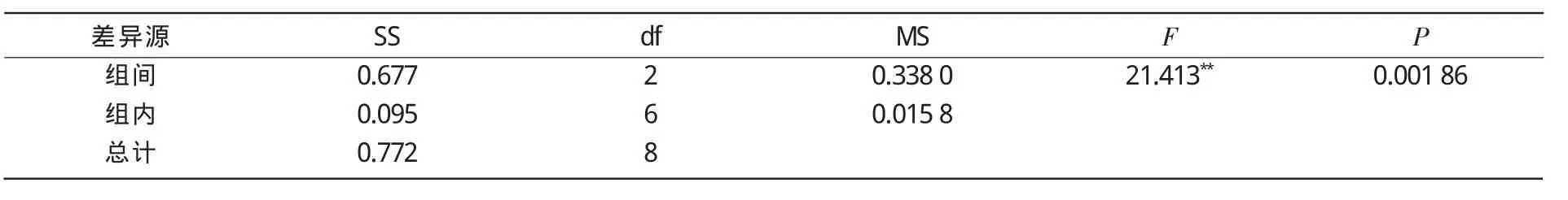
表2 杂交种、恢复系、保持系产量方差分析
对杂交大豆的产量与光合速率之间的关系进一步研究表明,大豆结荚期净光合速率与产量的相关系数为0.997 4,说明结荚期净光合速率与产量呈显著正相关。邹冬生等研究也认为,大豆在结荚鼓粒期间叶片的光合速率与籽粒产量呈显著正相关。有研究表明,大豆在生殖生长阶段的光合速率对籽粒产量影响最大。光合作用与产量密切相关,这种相关性从大田所获得的资料与水培所获得的资料一致[18]。
3 结论与讨论
(1)本研究对大豆生长的整个生育期的净光合速率、蒸腾速率、水分利用率、胞间CO2浓度、气孔导度测定结果表明,大豆杂交种SXNJCMS1A×中-21的净光合速率在盛花期、结荚期、鼓粒期高于亲本三系,特别是在结荚期显著高于其亲本三系。杂交种的蒸腾速率在结荚期显著高于恢复系、不育系、保持系,保证了源与库之间的贯通。杂交种的气孔导度和胞间CO2浓度在结荚期显著高于其他品种,可见,结荚期是杂交种整个生育期中最重要的一个时期。
(2)结荚期光合指标之间的相关性分析结果表明,净光合速率与蒸腾速率、气孔导度、胞间CO2浓度均呈正相关,与水分利用率呈极显著正相关。说明结荚期水分是影响光合速率的重要因素,此时期充足的水分有助于大豆产量的增加。
(3)方差分析结果表明,杂交种SXNJCMS1A×中 -21、恢复系中 -21、保持系 SXNJCMS1B这3个品种产量之间呈显著性差异,结荚期净光合速率与产量呈显著正相关。
杂种优势是生物界的普遍现象,它是指2个遗传组成不同的亲本杂交产生的杂种第1代,在生长势、生活力、繁殖力、抗逆性、产量和品质上比其双亲优越的现象。本研究表明,杂交种在光合性能和产量上存在优势,而且净光合速率与产量呈正相关。由此可以推测,杂交种的选育可以与高光效品种相结合,为杂交种的选育提供理论依据。
[ 1] Sinha S K,Khanna R.Physiological,biochemical and genetic basisof heterosis[J].Advancein Agronomy,1975,27:123-174.
[2]李海渤,袁美.作物杂种优势预测研究进展[J].河南农业科学,2004( 12):30-33.
[3]杨春玲,郭瑞林,关立.我国小麦杂种优势利用现状及存在的问题[J].河南农业科学,2002( 9):14-15.
[4]王艳玲,奚广升,王丕武,等.大豆不同杂交组合杂种优势分析[J].吉林农业科学,2008,27( 5):760-763,772.
[5]朱桂杰.大豆生理生态特性及其与产量的关系 [D].北京:中国科学院研究生院,2006.
[6]李少昆,赵明.玉米杂交组合光合特性研究[J].新疆农业科学,1999( 1):8-15.
[7]赵明,王树安,王美云.我国常用玉米自交系光合特性的聚类分析 [J].作物科学,1999,25( 6):733-741.
[8]王金陵.大豆 [M].哈尔滨:黑龙江科学技术出版社,1982:138.
[9]王雄健,薛文多,侯海鹏,等.不同密度春小麦群体的光合作用特性研究[J].天津农业科学,2008,14( 6):28-31.
[10]王小兵,李元清,崔国惠,等.不同类型小麦品种(系)光合生理特性差异研究[J].内蒙古农业科技,2011( 2):31-32,35.
[11]徐新娟,王伟,黄中文.大豆品种主要光合指标日变化的研究[J].河南农业科学,2011,40( 11):49-52,56.
[12]杨文杰,苗以农.大豆光合生理生态的研究:第2报 野生大豆和栽培大豆光合作用特性的比较研究 [J].大豆科学,1983,2( 2) :83-92.
[13]徐正进,陈温福,曹洪任.水稻穗颈维管束数与穗部性状关系的研究[J].作物学报,1998,24( 1):47-54.
[14]黄璜.水稻穗颈节间组织与颖花数的关系 [J].作物学报,1998,24( 2):113-120.
[15]高振福,庞振义,武长利.不同水分条件下大豆干物重增长的数学摸型与应用[J].大豆科学,1986,5( 4):299-304.
[16]常耀中,宋英淑.大豆需水规律与灌溉增产效果研究[J].大豆科学,1983,2( 4):277-285.
[ 17] Wells R,SchulzeL L,Ashley D A.Cultivar difference canopy apparent photosynthesis and their relationship to seed yield in soybeans[J].Crop Sci,1982,22:886-890.
[18]郝乃斌,戈巧英,杜维广,等.大豆高光效育种光合生理研究进展[J].植物学通报,1991,8( 2):13-20.
