蠋蝽人工繁殖技术及田间释放控制研究
2012-10-17王皙玮张李香范锦胜王贵强
高 卓,王皙玮,,张李香,孙 元,范锦胜,付 雪,金 娜,王贵强
(1.东北林业大学 林学院,哈尔滨 150040;2.黑龙江大学 农业资源与环境学院,哈尔滨 150080;3.中国农业大学 农学与生物技术学院,北京 100094)
0 引 言
蠋蝽 (Arma chinensis Fallou)是一种捕食性天敌昆虫,能捕食鞘翅目、鳞翅目、同翅目的叶甲、刺蛾、蚧壳虫、蚜虫类40余种农林害虫[1]。该蝽在我国南北方十几个省区广有分布,适于人工饲养,且人工释放的蠋蝽在释放地生态环境适宜条件下可定殖形成自然种群,达到持续控制害虫的目的。国内曾有报道利用蠋蝽防治榆紫叶甲,其生物防治效果明显。路红研究了Arma custos的外部形态特征[2],但中国国内分布的蠋蝽为A.chinensis[3];徐崇华等对北京地区蠋蝽的生活史、生活习性及捕食量曾做初步调查,而后对蠋蝽生长发育与温度的关系、卵和若虫期的发育起点、发育历期等做了进一步研究[4];高长启等研究了蠋蝽人工饲养天然饲料和半人工饲料对蠋蝽生命力影响的差异[5];陈跃均则对蠋蝽卵发育的有效积温进行了探究[6];姜秀华等对蠋蝽的生物学特性和对榆蓝叶甲的捕食效率进行了深入研究[7]。对蠋蝽的生物学特性、发生规律、人工饲养及捕食功能等虽有报道[5,8],但有关蠋蝽人工繁殖饲养中不同的供栖植物、饲养密度以及人工饲料对其生活力与寿命的影响及田间防治效果,目前尚鲜有报道。
有研究者认为全球气候变化对昆虫分布、个体发育、行为、生殖及生存会产生重要的影响[9],尤其是对一些在冬季死亡率较高的昆虫而言,气候变暖将大幅提高其种群存活数量[10]。昆虫的耐寒性主要采取迁飞或隐藏来逃避极端温度的伤害[11-13],通过调节体内自由水与结合水的比率[14-15]、游离脂肪酸与结合脂肪酸比例[16]、氨基酸代谢水平[17-18],改变抗寒物质储量,抵御极端温度[19-21]。也有研究报道对温度敏感的昆虫,在短时间内给予一定的低温锻炼能够显著提高其存活率及繁殖能力等[22-26]。昆虫自身生理调节及代谢相联系的快速冷驯化现象,对降低昆虫在短时间内温度急剧变化的伤害具有重要保护作用[27]。但也有研究者更倾向于快速冷驯化是对某一温度范围的应急反应,而不是随着温度变化对自身生理代谢的预调整[24,28-29]。有冷驯化研究证实,驯化时间延长其体内某些小分子抗冻物质将明显增加[30-31],昆虫的冷驯化机理也将随着现代分子生物学技术的发展从基因层面得以进一步阐释[32-33]。
本研究旨在深入探明蠋蝽人工饲养繁殖技术,以显著降低生产成本、提高产品质量,为工厂化、规模化及商品化生产蠋蝽提供科学依据[34-37]。还有,试验人工释放蠋蝽防治农业害虫,以求降低或减少农药的用量,避免因过度施药造成的环境污染及对人体健康的威胁,探索既具有显著的经济效益、生态效益和社会效益的同时,也为害虫的可持续治理新技术开辟新途径。
1 材料与方法
1.1 不同温度对蠋蝽成活率及发育历期和生殖力的影响
将供试的第3代蠋蝽初产卵20粒放入15cm口径的培养皿中,培养皿底部垫滤纸。而后将卵分别放入20℃、25℃与30℃3种温度处理的人工气候箱中培养,控制条件为70%R.H,光照周期为16L:8D。每处理3次重复。每日检查记录卵及若虫的发育情况,更换食物与滤纸,及时清除虫尸。当成虫羽化,观测记录产卵时间及产卵数量。
1.2 不同寄主植物和饲养密度对蠋蝽生长发育和生殖力的影响
蠋蝽越冬成虫以柞蚕蛹为饲料,正常交配产卵后收集同批卵在人工气候箱内培养至初孵若虫。室内实验于黑龙江大学农业资源与环境学院昆虫饲养室自然光照条件下进行,温度控制在25~27℃,65%~70%R.H。
设室内盆栽的2a生榆树、当年扦插的杨树枝条及盆栽大豆幼苗3个不同寄主植物处理与空白对照,每处理3次重复。盆栽植物其上罩一高30 cm,直径20cm,与花盆口径相同的自制圆柱状尼龙网罩笼。每处理放20头初孵蠋蝽若虫,每笼放4个柞蚕蛹作饲料,每天更换新鲜蚕蛹,盆内定期浇水保持植物正常生长。蠋蝽饲养密度实验将同期孵化的1龄若虫按4、10、20、30、40、50头/盆设6个处理,每日观测检查发育状况、死亡数量,及时清除虫尸,每处理3次重复。成虫羽化后记载产卵数量、日期以及孵化数量与日期。
数据采用Excel统计计算,SPSS12.0软件方差分析及Sigma plot软件作图。不同饲养密度其幼虫存活率引用Weibull分布函数拟合幼虫存活率曲线,即S (t)=exp (-btc)[34],其中S (t)为t时刻的存活率;b为尺度参数 (scale parameter);c为形状参数 (shape parameter),用以判断存活率曲线的类型,若c>1则表示死亡率系时间的增函数;若c=1时表示死亡率系常数;若c<1时表示死亡率系时间的降函数[35]。
1.3 蠋蝽不同分离部位提取物对其生殖力的影响
将500只性成熟尚未交配的蠋蝽雌虫腹部,足的腿节、胫节、跗节剪下分别研磨捣碎,用75%丙酮溶剂冲洗干净,于5℃条件下浸泡72h后过滤,滤液定容至25mL放入冰箱备用。实验设6个处理,10次重复。每处理放入1对初次交配中的蠋蝽,每日用移液枪抽取0.5mL提取液以及对照蒸馏水,分别滴滤纸上,待溶剂挥发后将滤纸放入相应培养皿中。
1.4 不同冷藏期对蠋蝽成虫、卵及若虫生活力的影响
将新羽化成虫置放于养虫笼中,用柞蚕蛹集体饲养。处理实验先在25±1℃,70%~80%R.H的环境下饲养4d,再于15±1℃,40%~50%R.H条件下饲养4d。将经过上述预处理的与未经预处理的成虫分别移入直径15cm培养皿中,皿底铺单层滤纸并将其均匀打湿,每日更放新鲜榆树叶片,置放冰箱 (4±1℃)进行冷藏。每处理设10头成虫,冷藏间隔期为5d,3次重复,之后放入冰箱 (4±1℃)进行冷藏。
将初产下的新鲜卵10粒移入培养皿,设冷藏5、7、10、15d等不同冷藏时间处理及对照,测定冷藏对蠋蝽卵的孵化率、若虫存活率的影响,每处理3次重复。
1.5 蠋蝽室内捕食量测定及田间防控甜菜夜蛾效果调查
室内捕食量测定将饲养的F2代蠋蝽成虫及若虫与不同龄期的甜菜夜蛾 (Laphygma exigua Hubner)幼虫为供试材料,待蠋蝽饱食24h,再饥饿24h后进行室内捕食功能测定。实验设9个处理,每处理引入1头蠋蝽个体及10头甜菜夜蛾个体,3次重复。每日观测记录蠋蝽对甜菜夜蛾的捕食数量。
将室内饲养的F3代蠋蝽成虫及若虫与不同龄期的甜菜夜蛾幼虫为供试材料,在黑龙江大学呼兰校区甜菜试验基地设2块5m×5m试验地,其中1块地为对照区,两地间隔200m。逐一统计试验地内甜菜夜蛾的数量,按蠋蝽与甜菜夜蛾1∶15的比例释放2龄若虫。每日调查记录不同虫态甜菜夜蛾数量,连续观测30d。
2 结果及讨论
2.1 不同温度对蠋蝽成活率及发育历期和生殖力的影响
3种不同温度条件处理蠋蝽卵其孵化率均为83%左右 (图1),处理间的结果无明显差异 (p>0.05)。但温度不同对蠋蝽若虫的发育历期影响极显著 (p<0.01),发育历期随着温度的升高而缩短 (图2),温度控制在20℃时,初孵若蝽发育至成虫需42.3d,而温度在30℃仅为29d。

温度对蠋蝽成虫寿命的影响也表现出极大的差异 (p<0.01),在20℃条件下饲养,成虫可存活43d,而在30℃时寿命仅为28.4d(图3)。然而,不同温度处理实验对蠋蝽产卵量的影响差异不显著(p>0.05),单雌产卵量均为200粒左右 (图4)。处理结果证实蠋蝽能够应对环境中温度的变化,有着良好的适应性。但是,其体内是否具有应对低温胁迫环境的特异性蛋白,从而产生具有快速的应变能力,还有待于进一步研究。

2.2 不同寄主植物和饲养密度对蠋蝽生长发育和生殖力的影响
寄主植物为蠋蝽若虫提供了栖息及取食环境,但寄主植物不同其存活率差异明显 (p<0.05)。其中,以榆树的存活率最高,可达82.0%,大豆为61.3%,杨树相对较低,只有34.6%,而无寄主植物状态下的空白对照其存活率最低,仅为16.3% (图5)。但不同的寄主植物对蠋蝽若虫发育历期影响差异并不显著 (p>0.05) (图6)。相反,蠋蝽雌虫的生殖力受寄主植物的影响明显,不同的寄主植物环境处理下,无论是产卵量、产卵周期、产卵前期还是次代卵的孵化时间均呈现出显著差异 (p<0.05)(图7)。榆树的蠋蝽产卵量最高,平均每雌可达330.8粒,大豆略低于榆树为255.7粒,杨树更低为225.3粒,而无寄主植物的卵量最低,仅96.6粒,为榆树的29.2%。然而,榆树和杨树为寄主植物的蠋蝽产卵前期之间并无明显差异,对照和大豆的产卵前期却明显比榆树和杨树的长。不同的寄主植物环境条件,蠋蝽的产卵期也表现出与产卵量相同的趋势,即榆树的蠋蝽产卵期长达17.9d,而杨树和大豆的只有13.4d和6.8d,无寄主植物的仅为5.8d。同样,不同寄主植物的次代卵孵化时间也表现出与上述实验相同的结果,依序为榆树的时间最短,为5.1d,大豆和杨树的时间稍长于榆树,而对照用时7.4d并仅有50%的个体孵化。

蠋蝽若虫单笼罩养密度在4、10、20及30头处理时,其存活率差异不显著,均可达85%以上,但密度至40头其存活率降至53.3% ,50头的存活率仅为44.6% (图8)。6种密度下的蠋蝽累计存活率拟合的 Weibull模型,其相关性>0.871 6(表1)。在30头/罩的密度下蠋蝽存活率曲线的c<1,说明其若虫存活率曲线是时间的增函数,即随着时间的变化其存活率越来越高。表1结果还表明,不同的饲养密度下b值无明显差异,说明在同一密度下,各龄期存活率相对一致。

图8 不同饲养密度下的蠋蝽幼虫存活曲线Fig.8 Survivorship curve of Arma chinensis on different reared density
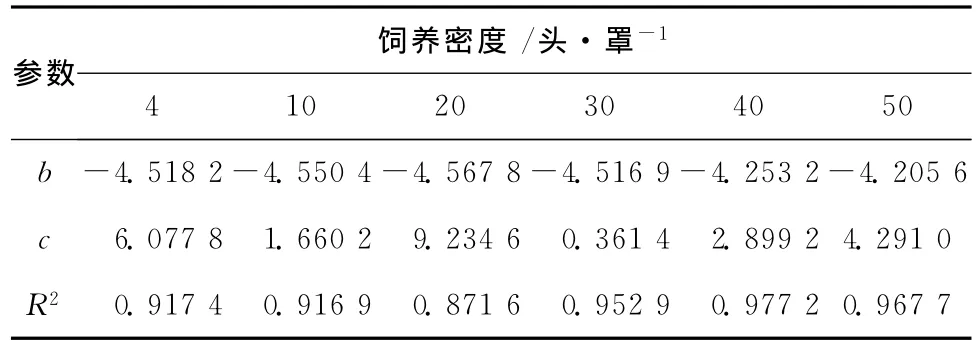
表1 不同饲养密度下蠋蝽若虫存活率曲线的Weibull模型参数值Table 1 Survivorship curve of parameter Weibull Model of nymph on different reared density
虽然饲养密度对蠋蝽1龄和5龄若虫的发育历期影响显著 (p<0.05),对其它龄期若虫的影响差异不明显,但处理间的总发育历期存在明显差异(p<0.05),即4头/罩>10头/罩>20头/罩>30头/罩>40头/罩>50头/罩 (表2)。
不同密度环境下人工饲养的蠋蝽,其产卵前期、产卵期及产卵量的差异明显,密度过高或过低其生殖能力均显著降低 (图9)。饲养密度为4头/罩和50头/罩时,其产卵前期最长为12.1d和13.6d。而对产卵期的影响则相反,密度过高或过低其产卵历期都不足8d,密度适宜时其产卵期均>11d。而对产卵数量影响的趋势与产卵历期相类似,表现为低密度和高密度的产卵量较少,只有100粒左右,而其它密度下的均在200粒左右。
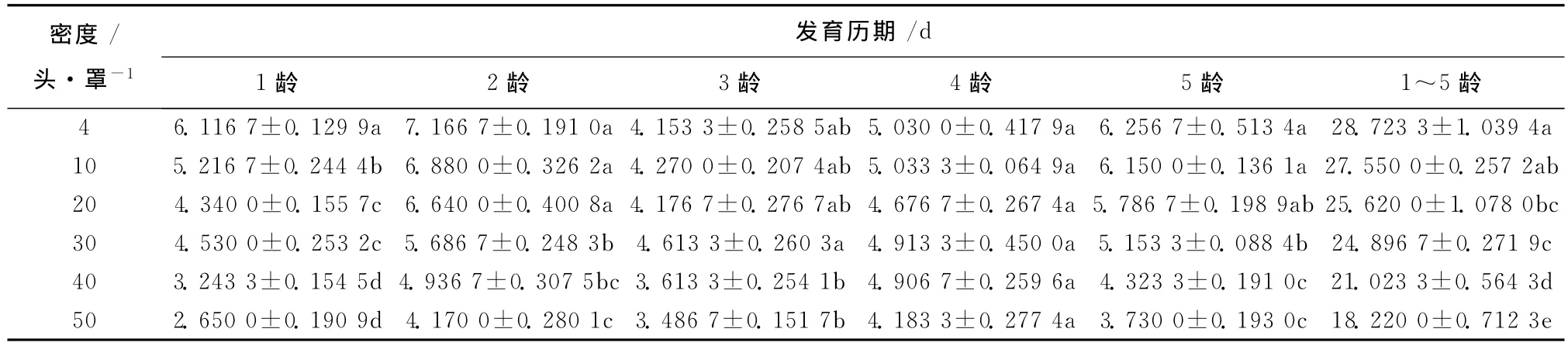
表2 不同饲养密度下蠋蝽幼虫各龄期发育历期Table 2 Developmental periods of nymph on different density

图9 不同饲养密度下的蠋蝽生殖力Fig.9 Fecundity of adult on different reared density
蠋蝽若虫饲养在以榆树幼苗为寄主植物环境下,可获得较多的成虫量及产卵量,但大豆幼苗的则略低于榆树的,而杨树幼苗比榆树和大豆幼苗的都低,其结果是因榆树叶片中粗蛋白、粗脂肪的量要比大豆和杨树叶片所含的量略高。另外,杨树幼苗能挥发出一种苦味油性多酚化合物,该类物质对某些昆虫具有拒食作用,因此杨树为寄主植物的蠋蝽其存活率和生殖力都明显低于榆树的。而大豆叶片中的粗脂肪含量比榆树和杨树的低,大豆为寄主植物饲养的蠋蝽其产卵期则比榆树和杨树的要短。榆树虽易获得,但培养周期较长,易落叶枯萎,成本较高。而大豆幼苗饲养蠋蝽虽然存活率和生殖力略低于榆树,但培养周期短,豆苗不易死亡,且可随时更换,成本较低。因此,建议在人工大量饲养蠋蝽时可首选大豆幼苗为寄主植物。在同一生活周期内,种群的同龄个体随时间推移而减少的现象,可以用存活曲线表示。目前国内外大多数学者均采用Weibull频数分布模型拟合动植物种群存活率与时间的关系描述种群存活率与时间变化的关系[37-38]。然而,该方法只能表示在某一时间正在死亡的种群个体数在原始观察时整个种群个体数所占的百分比,对于昆虫个体生长阶段的不同龄期,其死亡率差异很大,而研究者真正关心的是在某一时间段正在死亡的个体数占存活个体数的百分比。人工饲养蠋蝽在群体密度较高时由于空间及食物竞争,极易自相残杀,其个体活动异常活跃,存活率显著降低,生殖力也明显下降,既使种群恢复适宜密度,其生殖力在一段时期内依然维持在较低水平。有学者认为这是由于昆虫为了调节自身种群数量而释放了某种抑卵信息素后而导致的滞后作用。然而,在低密度饲养的蠋蝽其存活率虽差异不显著,但生殖力也显下降。有研究报道表明:单对饲养的棉铃虫产卵量和产卵期都比多对群养的低60%[39]。同样结果也在美洲棉铃虫饲养上得到证实,单雌饲养处女蛾的产卵量仅为10对饲养的1/3[40]。此种现象也同样表现在蠋蝽人工饲养过程之中。目前尚不清楚是否由于昆虫群体饲养过程中具有相互促进产卵作用,或是促卵与抑卵作用同时并存,究竟昆虫种群密度如何进行自身调节,仍有待深入研究,但上述研究结果表明人工繁殖饲养密度应以26头/m2为宜。
2.3 蠋蝽不同分离部位活性物质对产卵量的影响
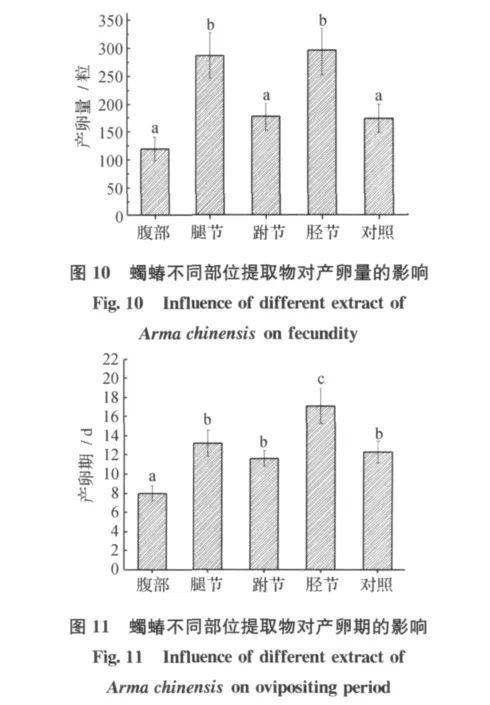
蠋蝽腿节和胫节的提取物对雌虫有明显的促排卵作用 (图10),经腿节和胫节提取物处理过的其产卵量分别比对照提高70.1%和64.3%。虽该促卵活性物质的成分以及作用机理尚不清楚,但用蠋蝽体驱不同体节提取物处理行产卵中的雌虫后,处理间的产卵历期也差异明显 (图11)。用胫节处理过的雌虫产卵天数明显比对照延长约5d(p<0.05),而用腹部处理过的雌虫,其产卵天数显著缩短,仅为8d。
2.4 不同冷藏期对蠋蝽成虫、卵及若虫生活力的影响
蠋蝽成虫寿命随着冷藏处理时间的延长,寿命明显缩短 (图12)。当冷藏时间延长至85d,成虫表现全部死亡。但在冷藏前对蠋蝽成虫进行逐渐降温预处理后再实施冷藏的,可显著延长其寿命40d左右。蠋蝽卵在较短的低温冷藏期内其孵化率无明显变化,但随着冷藏时间的延长,孵化率则显著降低 (p<0.05)。当冷藏时间延长至15d时,其孵化率仅为25.6% (图13)。温度处在14~30℃的范围内,蠋蝽卵均可保持较高的孵化率。温度提高卵的孵化时间缩短,在30℃时仅要3d,而在15℃时则需10d。在4℃低温条件下蠋蝽卵停止发育,若冷藏时间过长,卵会失去活力不能正常孵化。蠋蝽若虫也随着低温冷藏时间的延长,其存活率显著下降 (p<0.05)。当冷藏时间达15d时,其存活率比对照低60% (图14)。低温冷藏实验蠋蝽2龄若虫其耐饥及抗寒能力与成虫有明显差异。冷藏期内对存活率影响最大的是其耐饥能力,在对比实验中,对照组不进行饲喂,5d后其死亡率高达80%,而处理组中冷藏5d的,其死亡率仅为7%左右,冷藏反而提高了处饥饿条件下的存活率。这可能由于低温使其活动减弱体能消耗减少,从而提高其存活率和延长存活时间。

2.5 蠋蝽室内捕食量测定及田间防控甜菜夜蛾效果调查
蠋蝽成虫及4~5龄若虫对甜菜夜蛾1~2龄幼虫的捕食量最高,平均可达2.1头/d和2.0头/d,而对甜菜夜蛾5龄幼虫取食量最低,只有0.4头/d和0.3头/d(表3)。蠋蝽对甜菜夜蛾的田间捕食效应试验结果也显示相同结果,成蝽及4~5龄若蝽对甜菜夜蛾1~2龄幼虫的捕食量最高,捕食达1.9头/d和1.1头/d,而对甜菜夜蛾成虫则少有捕食,只有0.2和0.1头/d (表3)。蠋蝽室内捕食猎物的数量明显高于田间的捕食量,室内温度和环境条件干扰性小,而田间的昼夜温差及环境干扰会对取食产生一定的影响。
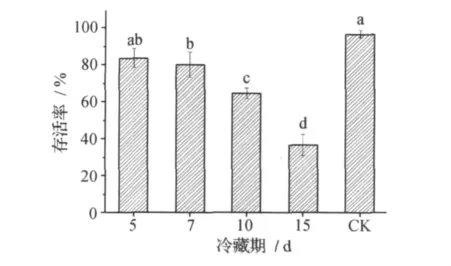
图14 不同冷藏期对蠋蝽若虫存活率的影响Fig.1 4 Nymph survival rate influenced by different cold storage
蠋蝽田间防治甜菜夜蛾试验以1∶15的比列释放蠋蝽2龄若虫。释放后的第13d其防治效果达63.8%,至第30d调查其虫口减退率达38.5%。试验结果表明,以1∶15或1∶10的比例进行释放,可对甜菜夜蛾种群增长起到良好的抑制作用。如果释放地其栖息环境及生态条件适宜,人工释放的蠋蝽可在释放地定居繁殖并形成自然种群,达到可持续控制害虫的目的。

表3 蠋蝽对甜菜夜蛾日平均捕食量调查Table 3 Predation number of Arma chinensis on Laphygma exigua /头·d-1
3 结 论
人工饲养繁殖蠋蝽其室内温度应控制在23~28℃,相对湿度在70%左右。饲料可用柞蚕蛹、黄粉甲幼虫或柞蚕幼虫等活体,但以柞蚕蛹作为饲料效果最佳。养虫室宜采用塑钢或铝合金的装饰材料,避免虫笼、墙壁等处发生霉变及细菌侵染。饲料要保证干净、新鲜,尤其不能以腐烂变质的饲料喂养蠋蝽。养虫室要经常通风,保证室内空气新鲜。
蠋蝽人工繁殖其种蝽的采集可在每年春、秋两季进行,一般以秋季的11月初冰雪封地前和春季的4月初田间化冻后为宜。采集地点通常选择位于于榆、杨混交林交界处越冬场所的鱼鳞坑内或者树根基部的落叶层下。采到的蠋蝽成虫放置存有10~15cm湿沙的养虫盒内,虫体平放其上复盖树叶,于4~6℃低温保藏120~150d,其成活率仍可达90%左右。也可在室外向阳避光处挖一深度为20~30cm的土坑,坑内先填15~20cm潮湿的细沙,将蠋蝽放置其上复盖些落叶,坑面用纱网罩好防止天敌侵害。此方法保藏蠋蝽成虫90~150d后,其存活率仍可达90%左右。取出越冬蠋蝽在室温15℃条件下1~2d缓缓复苏后,再将其移到环境条件为25℃,70%R.H养虫室内的30cm×30cm×40cm大小的养虫笼中。笼内需放水培杨树带叶枝条,每笼通常饲养30~50对,以柞蚕蛹为饲料,40~50d可繁殖一代。
蠋蝽若虫要严格区分龄期分别饲养,不同龄期若虫混养非但不便于管理,而且若虫相互残杀现象严重。另外,笼内要严格控制饲养密度。虽然养虫笼设计的大小有不同,养虫的数量差异较大,但原则上还是要根据若蝽是否取食方便,以及残杀现象是否严重而定。
饲养过程中,每天需清除虫笼中的粪便、虫尸及变质饲料,更换寄主植物时勿将虫卵或若虫带出。采卵应按收集时间不同分别培养,以保持虫龄发育的一致性。卵孵化后,将若蝽移放养虫笼内,在虫笼顶部与底部各放4~6个柞蚕蛹供其食用。蠋蝽人工大量饲养可选用大豆幼苗作为栖息植物,以提高蠋蝽的成活率并降低生产成本。
蠋蝽捕食量不仅与其虫龄高低有关,而且与猎物虫龄及体躯大小相关程度明显。蠋蝽成虫平均可捕食甜菜夜蛾1~2龄幼虫2.1头/d,老熟幼虫0.4头/d。田间释放蠋蝽防治甜菜夜蛾试验以蝽蛾1∶15的比率放蝽,13d后防治效果达63.8%,30 d后的虫口减退率达38.5%。
探明蠋蝽的生物学与生态学特性及田间释放控制技术,能够提高其人工饲养繁殖的数量及质量,实现天敌昆虫的人工生产养殖规模化、产业化及商品化,开创合理利用天敌资源推广生物防治与农林害虫可持续治理的新途径。
[1]萧刚柔.中国森林昆虫 [M].北京:中国林业出版社,1992:304-306.
[2]路 红,徐 伟,宋桂茹.蠋敌生物学的初步研究[J].吉林农业大学学报,1999,21 (1):33-34.
[3]郑乐宜.中国的蠋蝽属ArmaHahn(半翅目:蝽科)[J].昆虫天敌,1981,3 (4)28-32.
[4]徐崇华,严静君,姚德富.温度与蠋蝽 (Arma chinensis Fallou)发育的关系 [J].林业科学,1984,20(1):96-99.
[5]高长启,王志明,余恩裕.蠋蝽人工饲养技术的研究[J].吉林林业科技,1993,103 (2):16-18.
[6]陈跃均,乐国富,粟安全.蠋敌卵的有效积温研究初报 [J].四川林业科技,2001,(3):29-32.
[7]姜秀华,王金红,李振刚.蠋敌生物学特性及其捕食量的试验研究 [J].河北林业科技,2003,(3):7-8.
[8]陈 静,张建萍,张建华,等.蠋敌对双斑长跗萤叶甲成虫的捕食功能研究 [J].昆虫天敌,2007,29(4):149-154.
[9]Butterfield,A.Buse,J.C.Coulson,et al.Herbivory in global climate change research:direct effects of rising temperatures on insect herbivores [J].Global Change Biology,2002,(8):1-16.
[10]Harrington,R.,R,Fleming,I.P.Woiwood.Climate change impacts on insect management and conservation in temperate regions:can they be predicted?[J].Agricultural and Forest Entomology,2001 (3):233-240.
[11]马春森,马 罡,杜 尧,等.连续温度梯度下昆虫趋温性的研究与展望 [J].生态学报,2005,25(12):3 390-3 397.
[12]Hayward S A L,Worland M R,Convey P,er al.Temperature preferences of the mite,A laskozetes antarcticus,and the collembolan,Cryptopygus antarctius from the maritime [J].Antarctic Physiological Entomology,2003.28:114-121.
[13]Danks HV.Insect adap tations to cold and changing environments[J].Can.Entom ol.,2006,138:1-23.
[14]BlockW,Zettel J,Activity and dormancy in relation to bodywater and cold tolerance in a winter2active sp ringtail[J].Eur.J.Entom ol.,2003,100:305-312..
[15]马延龙,候 凤,马 纪.荒漠昆虫光滑鳖甲的耐寒性季节变化及其生理机制 [J].昆虫学报,2009,52(4):372-379.
[16]强承魁,杜予州,于玲雅,等.水稻二化螟越冬幼虫耐寒性物质的动态变化 [J].应用生态学报,2008,19 (3):599-605.
[17]韩瑞东,孙绪艮,许永玉,等.赤松毛虫越冬幼虫生化物质变化与抗寒性的关系 [J].生态学报,2005,25 (6):1 352-1 356.
[18]Nicholas M.Teets,Michael A.Elnitsky,Joshua B.Benoit,et al.Rapid cold-hardening in larvae of the Antarctic midge Belgica antarctica:cellular cold-sensing and a role for calcium [J].Am J Physiol Regulatory Integrative Comp Physiol,2008,294:1 938-1 946.
[19]赵 静,于令媛,李 敏,等.异色瓢虫成虫耐寒能力的季节性变化 [J].昆虫学报,2008,51 (12):1 271-1 278.
[20]Johannes Overgaard,Anders Malmendal,Jesper G.Sorensen,et al.Metabolomic profiling of rapid cold hardening and cold shock in Drosophila melanogaster[J].Journal of Insect Physiology,2007,53:1 218-1 232.
[21]Robert M.Michaud,David L.Denlinger.Shifts in the carbohydrate,polyol,and amino acid pools during rapid cold-hardening and diapause-associated coldhardening in flesh flies (Sarcophaga crassipalpis):a metabolomic comparison [J].J Comp Physiol,2007,177:753-763.
[22]Lee R E,Chen C P,Denlinger D L.A rapid coldhardening process in insects[J].Science,1987,238:1 415-1 417.
[23]Kelty J D,Lee R E.Rapid cold-hardening of Drosophila melanogaster(Diptera:Drosophilidae)during ecologically based thermoperiodic cycles [J].Journal of Experimental Biology,2001,204:1 659-1 666.
[24]Overgaard J,Sorensen J G,Petersen S O,et al.Changes in membrane lipid composition following rapid cold hardening in Drosophila melanogaster [J].J Insect Physiol,2005,51:1 173-1 182.
[25]Kostal V,Vambera J,Bastl J.On the nature of prefreeze mortality in insects:water balance,ion homeostasis and energy charge in the adults of Pyrrhocoris apterus[J].Journal of Experimental Biology,2004,207:1 509-1 521.
[26]Shreve S M,Kelty J D,Lee R E.Preservation of reproductive behaviors during modest cooling:rapid cold-hardening fine-tunes organismal response [J].Journal of Experimental Biology,2004,207:1 797-1 802.
[27]Koveos D S.Rapid cold hardening in the olive fruit fly Bactrocera oleae under laboratory and field conditions[J].Entomologia Experimentalis et Applicata,2001,101:257-263.
[28]Michaud M R,Denlinger D L.Oleic acid is elevated in cell membranes during rapid cold-hardening and pupal diapause in the flesh fly,Sarcophaga crassipalpis [J].J Insect Physiol 2006,52:1 073-1 082.
[29]Storey J M,Storey K B.Regulation of cryoprotectant metabolism in the overwintering gall fly larva Eurosta solidaginis:temperature control of glycerol and sorbitol levels[J].J Comp Physiol,1983,149:495-502.
[30]Chen C P,Lee R E,Denlinger D L.Cold shock and heat shock:a comparison of the protection generated by pretreatment at less severe temperatures [J].Physiol Entomol,1991,16:19-26.
[31]Yoshihiro Fujiwara,David L.Denlinger.MAPK is a likely component of the signal transduction pathway triggering rapid cold hardening in the flesh fly Sarcophaga crassipalpis [J].The Journal of Experimental Biology,2007,210:3 295-3 300.
[32]Stephen R.Misener,Cheng-Ping Chen,Virginia K.Walker.Cold tolerance and proline metabolic gene expression in Drosophila melanogaster [J].Journal of Insect Physiology,2001,47:393-400.
[33]景晓红,康 乐.昆虫耐寒性研究 [J].生态学报,2002,22 (12):2 202-2 207.
[34]William H G..Econometric analysis [M].New York:New York University,1997:221-222.
[35]Pinder J E,Winer J G,Smith M H.The Weibull distribution:A new method of summarizing survivorship data [J].Ecology,1978,59 (1):175-179.
[36]刘 怀,赵志模,邓永学,等.温度对竹盲走螨实验种群数量消长的影响 [J].动物学研究,2002,23(4):356-360.
[37]周 蕊,陈 力.饲养密度对洋虫生长发育的影响[J].昆虫知识,2006,43 (3):389-391.
[38]李国清,郭慧芳.不同饲养密度对棉铃虫蛾寿命和生殖力的影响 [J].南京农业大学学报,1998,21(1):46-50.
[39]Rellae L.A,Peter E A,James H T.Age and crowing affects the amount of sex pheromone and the oviposition rates of virgin and mated females if Helicouerpa zea(Lepidoptera:Noctuidae)[J].Annals of the Entomological Society of America,1994,87:350-354.
