20个青菜品种的遗传多样性分析
2012-06-29马晓静李广平郑晓静林冬枝董彦君
马晓静 ,李广平 ,郑晓静 ,林冬枝 ,2,董彦君 ,2
(1.上海师范大学生命与环境科学学院,200234;2.上海师范大学遗传研究所)
青菜(Brassica chinensisL.)属十字花科芸薹属青菜种,别称小白菜、青梗白菜等,是我国常见的绿叶蔬菜之一[1],在我国长江三角洲地区占有着重要地位,市场需求量大[2,3]。目前对青菜种质资源的鉴别主要依据形态学,而在DNA分子水平对其遗传多样性研究不多。李红斌等[4]利用AFLP分子标记技术对浙江省主栽的10个青菜品种进行了遗传多样性及聚类分析。近年来,更多DNA分子标记的发展及应用[5~11],为种质资源遗传多样性的研究提供了更多手段,其中SSR分子标记技术广泛应用于种质资源遗传多样性方面的研究[12~15],但未见利用SSR分子标记技术对青菜进行遗传多样性分析的报道。本研究利用10对SSR分子标记技术对来自中国和日本的20个青菜品种进行遗传多样性分析,探索其亲缘关系,以期为青菜种质改良和创新提供分子水平上的理论依据。
1 材料与方法
1.1 试验材料
供试材料为来自我国长江三角洲地区主栽的8个青菜品种以及从日本引进的12个青菜品种,具体见表1。2012年4月1日将其播种于上海师范大学奉贤校区生物基地内,30 d后取其植株的新鲜幼嫩叶片,-70℃保存备用。

表1 供试青菜品种
1.2 DNA模板的提取
用改良的CTAB法[16]提取青菜叶片基因组DNA,-20℃保存。
1.3 引物筛选
本研究选用88对随机引物对样品进行PCR扩增,筛选出谱带清晰、位点数量及产物大小适中且适用于20个青菜品种遗传多样性分析的引物,对样品进行正式扩增。
1.4 PCR扩增及聚丙烯酰胺凝胶电泳
PCR反应在eppendorf扩增仪上进行,反应体系为 10 μL, 其中 1 μL 10×Buffer with Mg2+,0.2 μL 2.5 mmlo/L dNTP,0.25 μL 1 μmol/L 引 物 ,0.25 μL 5 U/μL Taq 酶,5 μL 模板 DNA,最后用无菌水补足至 10 μL。 反应条件如下:94℃预变性 4 min,94℃变性 30 s,50℃退火 30 s,72℃延伸 60 s,35 个循环;最后72℃延伸10 min,产物4℃保存备用。扩增产物在8%的聚丙烯酰胺非变性凝胶上电泳分离3.5 h(恒定功率30 W),以银染法检测电泳结果。
1.5 数据统计与分析
PCR扩增产物在银染过的聚丙烯酰胺凝胶上都表现为谱带的“有”和“无”,分别将其赋值,有带的记为1,无带的记为0,建立所用材料分子标记的0、1矩阵。根据表征矩阵,统计扩增产物的条带总数和多态性条带数量。
利用NTSYS 2.10e软件分析材料间的Nei's遗传相似系数。根据遗传相似系数矩阵,用非加权类平均法(UPGMA法)进行聚类分析,建立聚类图。
2 结果与分析
2.1 引物筛选
从88对随机引物中筛选出谱带清晰、位点数量及产物大小适中且适用于遗传多样性分析的10对引物(表2)。其扩增产物的条带总数和多态性条带结果见表3。由表3可知,其扩增的条带总数为26条,其中多态性条带有20条,占扩增条带总数的76.9%,说明供试青菜品种间具一定的遗传差异。
2.2 聚类分析
由图1可知,以遗传相似系数0.708为界,可将20个青菜品种划分为3类,其中第Ⅰ类包括CR夏专科、夏八仙、长江;第Ⅱ类包括大叶黑大头、嘉兴矮脚青、上海五月慢、特矮苏州青、上海矮抗青;第Ⅲ类包括长阳、上海四月慢、夏三味、敦煌、夏赏味2号、NiHAO 夏、阳夏、一江、华王、阳太、抗热 605、紫冠一号(表 4)。

表2 引物筛选结果
3 结论与讨论
本研究所筛选出的10对SSR分子标记中绝大多数的多态性比率较高(表3),20个青菜品种的遗传相似系数在0.461 5~0.923 1,变异范围较大,说明其遗传多样性较高,同时也说明,SSR分子标记可作为青菜分子育种中有力的辅助工具。
另外,利用10对SSR分子标记的聚类分析结果表明,供试的20个青菜品种可分为三大类 (表4),其中第Ⅰ类中CR夏专科、夏八仙、长江均来自日本;第Ⅱ类中大叶黑大头、嘉兴矮脚青、上海五月慢、特矮苏州青、上海矮抗青则均来自我国;而第Ⅲ类中除上海四月慢、抗热605、紫冠一号以外,其余均为日本品种。本试验中将来自中国的青菜品种五月慢和抗热605归属为不同类群,与李红斌等[4]的结果一致。总之,本试验结果显示绝大部分供试青菜品种,来自同一国家的聚在一起,说明其亲缘关系较近,而来自不同国家的青菜品种之间的亲缘关系则较远,这可能是因为长久缺乏种质交流而使绝大部分供试青菜品种呈现明显的地域性。由此可见,地域限制可能是促成青菜种质资源遗传分化的主要原因之一。因此,我们可以引入外来种质,将其与国内青菜品种进行杂交,实现性状的改良,以满足青菜品种的市场需求。

表3 10对引物的扩增结果
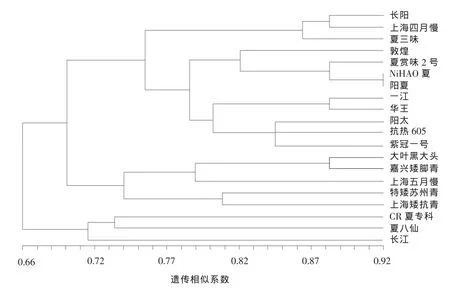
图1 20个青菜品种的UPGMA聚类分析

表4 供试材料聚类结果
[1]郝春燕.上海地区青菜品种应用现状及潜力品种推荐[J].长江蔬菜,2009(7):9-11.
[2]骆义成,戚立刚,沈玉兴,等.青菜新品种夏帝引种示范试验项目总结[J].上海蔬菜,2010(6):10.
[3]龚静,朱玉英,吴晓光,等.青菜新品种“新矮青”的选育[J].上海农业学报,2002,18(1):39-42.
[4]李红斌,王世恒,王宏.青菜品种亲缘关系AFLP分析 [J].北方园艺,2008(8):39-40.
[5]Thomson M J,Septiningsih E M,Suwardjo F,et al.Genetic diversity analysis of traditional and improved Indonesian rice(Oryza sativaL.)germplasm using microsatellite markers[J].Theoretical and Applied Genetics,2007,114(3):559-568.
[6]Ma C Y,Ma H Y,Xu Z,et al.Isolation and characteristics of polymorphic microsatellite markers inPennahia argentatus[J].Biochemical Systematics and Ecology,2012,43:159-162.
[7]王娟,郭大龙,侯小改,等.不同花型牡丹品种亲缘关系的SRAP 分析[J].中国农学通报,2011,27(28):167-171.
[8]魏玉杰,张金文,何庆祥,等.不同生态区罂粟种质的遗传多样性 ISSR 分析[J].植物遗传资源学报,2012,13(2):239-243.
[9]王爱华,季祥彪,文晓鹏.16份野生半夏的遗传多样性分析[J].贵州农业科学,2012,40(1):15-19.
[10]刘丽,杨新龙,刘楚吾,等.南海海域常见龙虾的遗传多样性分析[J].水产科学,2012,31(3):160-164.
[11]孙立夫,裴克全,张艳华,等.中国与欧洲高卢蜜环菌的遗传多样性[J].生物多样性,2012,20(2):224-230.
[12]徐建欣,王云月,姚春,等.利用SSR分子标记分析云南陆稻品种遗传多样性 [J].中国水稻科学,2012,26(2):155-164.
[13]倪胜利,李兴茂,杨德龙,等.国内外冬小麦品种(系)的遗传多样性分析[J].西北农业学报,2012,21(2):20-25.
[14]刘龙洲,翟文强,陈亚丽,等.设施用厚皮甜瓜品种SSR标记遗传多样性分析 [J].植物遗传资源学报,2012,13(3):381-385.
[15]曾维英,梁江,陈渊,等.广西野生大豆SSR标记的遗传多样性研究[J].江苏农业科学,2012,40(3):22-25.
[16]Murray M G,Thompson W F.Rapid isolation of high molecular weight plant DNA[J].Nucleic Acid Res,1980,8:4 321-4 326.
