高黎贡山赧亢白眉长臂猿春秋季活动范围变化
2011-11-20邓忠坚
李 旭, 邓忠坚, 周 伟, 白 冰
(西南林业大学保护生物学院, 云南省森林灾害预警与控制重点实验室, 云南昆明 650224)
高黎贡山赧亢白眉长臂猿春秋季活动范围变化
李 旭*, 邓忠坚, 周 伟, 白 冰
(西南林业大学保护生物学院, 云南省森林灾害预警与控制重点实验室, 云南昆明 650224)
以云南省高黎贡山国家级自然保护区赧亢片区为研究地点,对1雌1雄1幼组成的白眉长臂猿家群展开为期4个月的持续跟踪.春季和秋季分别获得7个和5个全天跟踪数据,有效跟踪时间达176.0 h.白眉长臂猿有相对固定的夜栖区和取食区,春季活动区域面积仅占秋季活动区域面积的45.9%.春季白眉长臂猿的夜栖区和取食区集中在草果种植区内.秋季白眉长臂猿的夜栖区和取食区不与草果种植区重叠.人类草果种植没有影响白眉长臂猿的分布,但秋季草果采收却干扰白眉长臂猿取食和夜栖.
高黎贡山; 白眉长臂猿; 季节变化; 活动
白眉长臂猿 (Hoolockhoolock) 隶属于灵长目 (Primates) 长臂猿科 (Hylobatidae),为东洋界的缅甸-中国亚区的特有种,属国家I级重点保护动物.目前高黎贡山国家级自然保护区是国内目前确定有白眉长臂猿种群的唯一分布区,据2007年调查结果[1],其赧亢片区、白花岭片区和大塘片区现有白眉长臂猿野生种群15~20群、25~40只,中国白眉长臂猿种群数量持续下降.
白眉长臂猿为树栖性灵长类,偏好在树冠层分层明显的原始林中活动,且活动范围呈季节性变化[2-4].它们对生活环境具较高的依赖性[5],由于人类社区的发展,经济林种植和村庄的扩大,现长臂猿的栖息地受到极大干扰,并且被迫与人共同分享森林资源[6].至2006年开始,西南林业大学就对分布于赧亢片区的白眉长臂猿种群进行了系列生态学的专项研究,内容涉及栖息地利用及食物资源调查等[1,7-11].调查发现,该地区的种群对栖息环境具高度的选择性,尤其在人为干扰最大的秋季,当地草果种植是最主要的干扰模式.春秋食物资源的差异是导致两季猿群活动的海拔范围变化的原因之一[1,7-9].对白眉长臂猿游走行为研究尚属空白.本文研究白眉长臂猿的游走行为,可了解该物种对栖境的利用强度和活动模式,丰富该物种的行为学研究资料,有助于分析其活动范围的季节性变化与干扰和食物之间的关系,为白眉长臂猿的有效保护和食物的供给提供理论依据.
1 研究地点与时间
研究时间为2009年,为保障徒步跟踪调查过程中人员安全,仅选择气候较为温暖、干燥的春、秋两季展开跟踪;冬季气候寒冷,夏季多雨,不仅难以观察到白眉长臂猿实体,而且存在较大的安全隐患,暂未开展跟踪调查工作.选择中国云南省高黎贡山赧亢生境走廊带的惠坡 (最高海拔2 311 m)、大马河塘和栽山沟等地点为研究地,该地区分布有白眉长臂猿3~4群,6~8只.经过西南林业大学白眉长臂猿工作组2年的努力,已经成功与1雌1雄1幼组成的家群习惯化,保持一定距离的长时间观察不会影响该家群的行为模式,本项工作即选择该家群为跟踪对象.
2 研究方法
白眉长臂猿种群数量极其稀少.国内还没有捕捉到野生白眉长臂猿,缺乏在其身上安装无线电或卫星发射器的先例,目前对白眉长臂猿开展的所有生态学研究均是以徒步调查的方式完成.白眉长臂猿在中国仅分布于云南省滇西地区的原始常绿阔叶林中,且偏好在树冠层间摆荡、跳跃,移动速度快,难于跟踪.在区域中设立5个听声站,工作组共4人,2人为1队,每天7:00开始进入听声站,听到白眉长臂猿晨鸣后,确定其活动方向和位置后徒步跟踪.在猿群两侧保持20~30 m距离持续跟踪,避免对猿群产生驱赶、堵截等惊扰行为.以30 min为时间间隔,在正点和半点 (例如7:30、8:00、8:30) 记录白眉长臂猿群体的位置.综合利用Magellan explorist 600型GPS接收机和1∶5 000的地形图上明显的地形特征,迅速且准确地记录猿群的活动位点.鉴于草果是赧亢地区唯一的农业作物,调查过程中特别勾绘草果种植区的边缘,便于辅助分析白眉长臂猿对生境的利用情况.
采用实地跟踪记录位点的方法,在视线可及的时间段收集家群位点数据,一旦家群脱离视线,则需要重新寻找、定位后再次进行数据收集.鉴于白眉长臂猿行动敏捷,跟踪难度较大,在后期数据整理时,仅选择持续跟踪的全天 (7:00—18:30) 和时间段较长的位点数据进行分析,以保证数据的代表性.运用AcrMap 9.3软件中的制图功能将1∶5 000的地形图按100 m×100 m的标准网格化,将猿群所有的活动位点在AcrMap 9.3中制成活动图层,按春、秋两季叠加.运用猿群位点在不同网格中出现的频次,分析猿群对不同区域的利用强度.借鉴C指数(C=b/a)检验相近2天猿群对区域利用的集中程度,b是相近2天中第2次猿群进入的新网格数,a是相近2天中第2次猿群进入的总网格数[12].C<0.77,代表研究对象对区域的利用相对集中;C≥0.77,代表研究对象对区域的利用相对分散[13-14].
3 结果与分析
3.1活动位点比较
春季野外跟踪工作时间为2009-03—04,对1雌1雄1幼组成的家群跟踪观察,发现猿群活动29天,获得7个全天 (7:00—18:30) 的跟踪数据;累积跟踪时间80.5 h,有效位点168个 (图1(a)).秋季野外跟踪工作时间为2009-10—11,跟踪对象与春季相同,发现猿群活动34天,其中有5个全天(7:00—18:30)的跟踪数据,以及相对完整的另外5个半天的跟踪数据;累积跟踪时间95.5 h,有效位点201个 (图1(b)).
高黎贡山赧亢生境走廊带的白眉长臂猿活动区可分为156个100 m×100 m的网格,对所有网格按A (1~13) -L (1~13) 编号 (图1).春季记录到白眉长臂猿活动的网格26个,经过活动但未记录位点的网格2个,总网格数28个 (图2(a));秋季记录到白眉长臂猿活动的网格38个,经过活动但未记录位点的网格23个,总网格数61个 (图2(b));白眉长臂猿春季活动区域明显小于秋季活动区域.

图1 白眉长臂猿活动位点分布图Figure 1 Map showing active distribution of the Hoolock hoolock
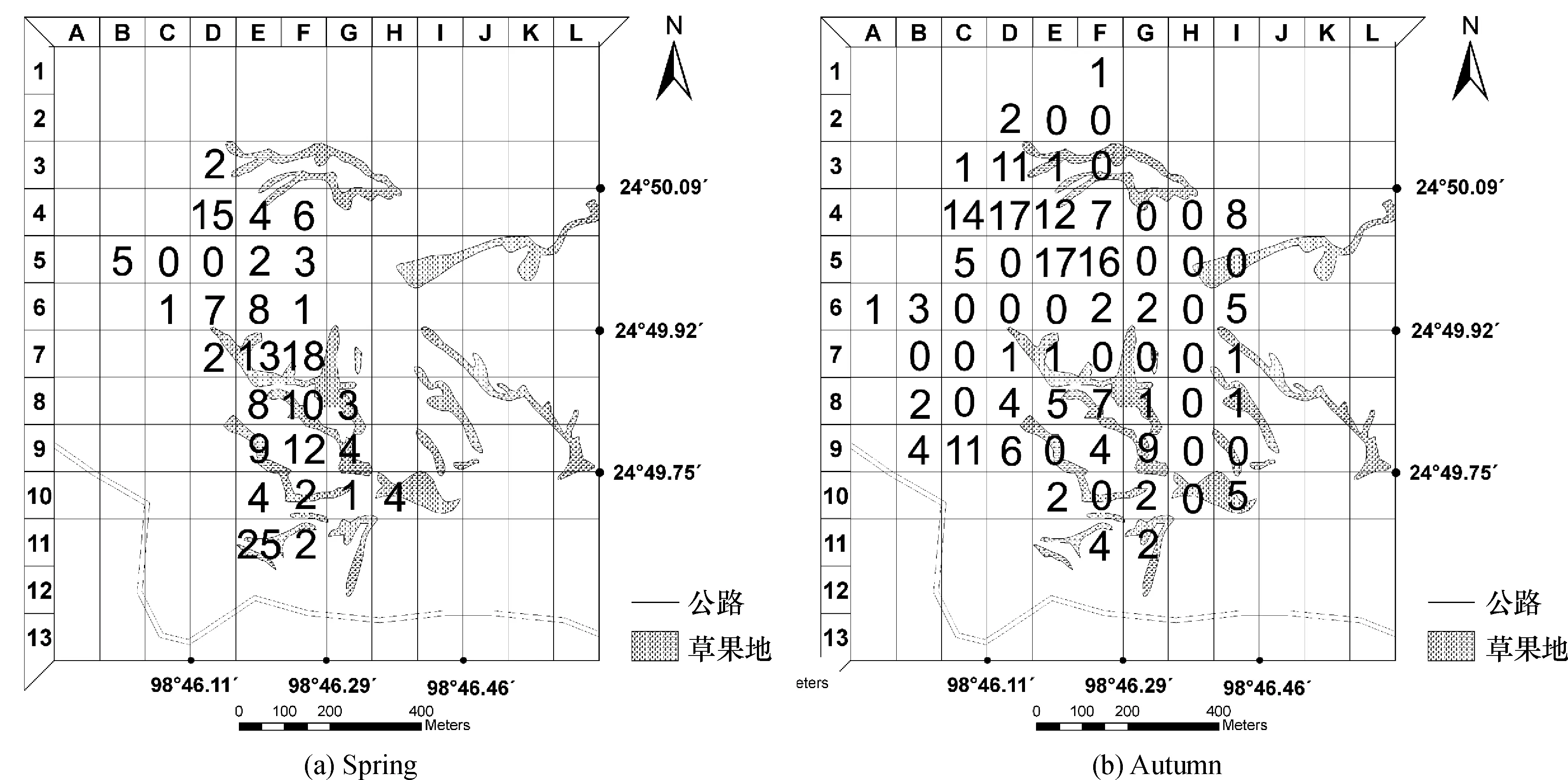
图2 白眉长臂猿活动位点统计Figure 2 Counting of active distribution of the Hoolock hoolock
白眉长臂猿春季利用的网格有28个,其中E11是利用频次最高的区域 (25次),C5和D5利用频次最低 (0次).初步按0~25利用频次等分为3个相对利用等级,0~8为低利用强度,区域有21个;9~16为中利用强度,区域有5个;17~25为高利用强度,区域有2个 (图2(a)).白眉长臂猿秋季利用的网格有61个,其中D4和E5是利用频次最高的区域 (17次),初步按0~17利用频次等分为3个相对利用等级,0~6为低利用强度,区域有50个;7~12为中利用强度,区域有7个;13~17为高利用强度,区域有4个 (图2(b)).
3.2区域利用分析
春季7个全天和秋季5个全天的跟踪数据被用来分析白眉长臂猿对区域利用的集中性.春季共6次C值计算结果表明,66.7%表现为对活动区域集中利用,33.3%表现为分散利用;秋季共4次C值计算结果,50%表现为对活动区域集中利用,50%表现为分散利用 (表1).
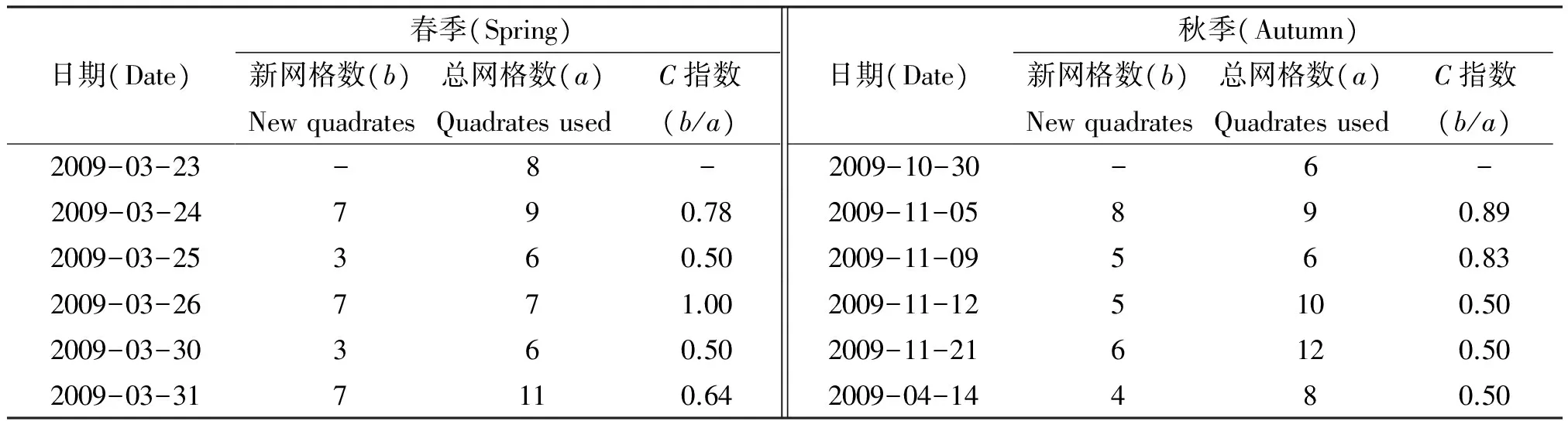
表1 白眉长臂猿区域利用值 (C values) 计算Table 1 C values computed for the Hoolock hoolock
4 讨论
4.1白眉长臂猿春季和秋季夜栖区存在差异
白眉长臂猿没有固定的夜栖树,却具有固定的夜栖区.春、秋两季13个全天的跟踪发现,他们清晨晨鸣的树木即是前夜休息的夜栖树,并且晨鸣后还在该树上持续活动较长时间.另外,黄昏猿群也多在夜栖区内游走.这就是调查组于春季黄昏和清晨多次在高强度利用区E11区中发现猿群活动 (图2(a))的原因,高黎贡山国家级自然保护区赧亢管理站工作人员也经常在E11区中听见白眉长臂猿晨鸣,可以确定E11区是白眉长臂猿春季的主要夜栖区.D4区是春季中强度利用区和秋季高强度利用区,E5是秋季高强度利用区 (图2),调查组在确定白眉长臂猿夜栖后的次日,在相同区域中发现猿群晨鸣;D4区是白眉长臂猿长年固定的夜栖区,E5是猿群秋季主要夜栖区.
4.2白眉长臂猿秋季取食区较春季分散
食物资源和人为干扰的季节性变化导致白眉长臂猿春季取食集中在草果种植区,而秋季取食避开草果种植区,取食点较为分散.白眉长臂猿春季日活动时间分配中,休息仅占35%,取食和移动分别占32%和33%,长时间在漫游过程中觅食是白眉长臂猿取食特点[9].猿群春季对区域的利用相对集中 (表2),除固定的夜栖区E11和D4外,猿群对E7、E9、F8、F9等区域呈中强度利用,对F7区呈高强度利用 (图2(a)),白眉长臂猿多在E7、E9、F8、F9和F7等5个区域中反复游走觅食.虽然春季可选择的果实仅3种,但仍是猿群取食频次最高的食物[10],由于果实种类少且分布范围窄,形成了白眉长臂猿对该5个栅格的反复利用,同时春季无人进山看守草果地,干扰程度小,亦使白眉长臂猿能放心地在草果地种植区觅食.而秋季猿群对区域的利用较春季分散,集中分布比率由66.7%下降到50%(表2).虽然白眉长臂猿的活动区仍然与草果种植区重叠,但主要的取食区已经避开草果种植区,集中在C4、E4、F5、C9等非草果种植区取食.秋季果实类食物增至11种[11],种类较多且分布分散,白眉长臂猿选择喜食食物的地域增加,是导致其秋季集中分布比率下降的原因.这种现象与婆罗洲杂交长臂猿 (灰长臂猿与黑掌长臂猿的杂交种Hylobatesmuelleri×agilis) 的食性研究结果相似,当食物资源丰富时,动物总是寻找几种喜食食物[15].另外,秋季正是草果采摘季节,由于人为干扰强度高,白眉长臂猿在觅食过程中避开草果种植区,取食点较为分散.
4.3秋季草果采收干扰白眉长臂猿取食和夜栖
草果的种植没有影响白眉长臂猿分布,但秋季草果的采收却干扰了白眉长臂猿取食和夜栖.调查结果发现,该片区白眉长臂猿主要在草果种植区活动 (图 1),长期的人-猿共同利用同一区域,猿群似乎已习惯这种无害化干扰.然而从分布区域上来看,猿群春季活动区域的面积仅占秋季活动区域面积的45.9% (图1),虽然食物变化是其中的原因之一,但是人为干扰却是不可忽视的因素.这从秋季猿群的夜栖区和取食区均不再与草果种植区重叠的现象可以证明,这种改变有草果采收干扰的作用.类似的相关研究结果也证明,受草果采收影响,黑冠长臂猿的生境利用行为和正常活动受到干扰[16].该片区社区居民通过社区共管参与保护白眉长臂猿,使得人-猿能够形成和谐的共存关系,但夜栖区和取食区仍是白眉长臂猿不希望受人类干扰的“私人空间”.
[1] 张兴勇,白冰,艾怀森,等. 云南高黎贡山自然保护区白眉长臂猿种群及数量现状初报[J]. 四川动物,2007,27(2):856-858.
ZHANG Xingyong,BAI Bing,AI Huaisheng,et al. Population and quantity of Hoolock Gibbon(Hylobateshoolock) in Gaoligongshan Nature Reserve[J]. Sichuan Journal of Zoology,2007,27(2):856-858.
[2] ALFRED J,SATI J P. Survey and census of the Hoolock Gibbon in West Garo Hills,Northeast India [J]. Primates,1990,31(2):299-306.
[3] ISLAM M A,FEEROZ M M. Ecology of Hoolock Gibbon of Bangladesh [J]. Primates,1992,33(4):451-464.
[4] SATI J P,ALBIDE J. Hoolock Gibbon (Bunopithecushoolock) [J]. Wildlife and Proteceted Areas,2001,1(1):60-67.
[5] BRIEN T,KINNAIRD M F,NURCAHYO A,et al. Abundance and distribution of sympatric gibbons in a threatened Sumatran rain forest [J]. International Journal of Primatology,2004,25,267-284.
[6] MUZAFFAR S B,ISLAM M A,FEEROZ M M,et al.Habitat characteristics of the endangered Hoolock Gibbons of Bangladesh:The role of plant species richness [J]. Biotropica,2007,39(4):539-545.
[7] 白冰,周伟,艾怀森,等. 高黎贡山赧亢白眉长臂猿春季栖息地利用[J]. 动物学研究,2007,28(2):179-185.
BAI Bing,ZHOU Wei,AI Huaisen,et al. Habitat use of the Hoolock Gibbon (Hoolockhoolock) at Nankang,Mt. Gaoligong in spring [J]. Zoological Research,2007,28(2):179-185.
[8] 白冰,周伟,艾怀森,等. 高黎贡山大塘白眉长臂猿春季栖息地利用[J]. 四川动物,2008,27(4):626-630.
BAI Bing,ZHOU Wei,AI Huaisen,et al. Habitat use of Hoolock Gibbon (Hoolockhoolock) at Datang,Mt. Gaoligong in spring [J]. Sichuan Journal of Zoology,2008,27(4):626-630.
[9] 张兴勇,吴建普,周伟,等. 云南高黎贡山赧亢白眉长臂猿春季食谱及活动时间分配初探[J]. 四川动物,2008,27(2):193-204.
ZHANG Xingyong,WU Jianpu,ZHOU Wei,et al. Diet and time budget of Hoolock Gibbon (Hoolockhoolock) at Nankang,Gaoligongshan Mountains in spring [J]. Sichuan Journal of Zoology,2008,27(2):193-204.
[10] 张兴勇,周伟,吴建普,等. 高黎贡山赧亢白眉长臂猿春季食物选择[J]. 动物学研究,2008,29(2):174-180.
ZHANG Xingyong,ZHOU Wei,WU Jianpu. Food selection of Hoolock Gibbon (Hoolockhoolock) at Nankang,Mt. Gaoligong in spring [J]. Zoological Research,2008,29(2):174-180.
[11] 吴建普,周伟,周杰珑,等. 高黎贡山赧亢白眉长臂猿食性及日取食量[J]. 动物学研究,2009,30(5):539-544.
WU Jianpu,ZHOU Wei,ZHOU Jielong. Diet and daily feeding amount of Hoolock Gibbon (Hoolockhoolock) at Nankang,Mt.Gaoligong [J]. Zoological Research,2009,30(5):539-544.
[12] BUZZARD P J. Ranging patterns in relation to seasonality and frugivory amongCercopithecuscampbelli,C.petaurista,andC.Dianain the Tai Forest [J]. International Journal of Primatology,2006,27(2):559-573.
[13] KIM R M,ANTION A,FIRMANN A,et al. Influence of forest season ablity on Gibbon food choice in the rain forests of Baritoulu,Central Kalimantan [J]. International Journal of Primatology,2003,24 (1):19-32.
[14] 倪永庆,马世来. 滇南、滇东南黑冠长臂猿分布与数量. 动物学研究,2006,27 (1) :34-40.
NI Yongqing,MA Shilai. Population and distribution of the black crested Gibbons in southern and southeastern Yunnan [J]. Zoological Research,2006,27 (1) :34-40.
[15] STRIER K B. Ranging behavior of woolly spider monkeys,or muriquis,Brachytelesarachnoids[J]. International Journal of Primatology,1987,8:575-591.
[16] KAPLIN B A. Ranging behavior of two species of guenons (CercopithecuslhoestiandC.mitisdoggetti) in the Nyungwe Forest Reserve,Rwanda [J]. International Journal of Primatology,2001,22(4):521-548.
Keywords: Mt. Gaoligong; Hoolock gibbon; seasonal changes; action
【责任编辑 成 文】
SEASONALCHANGESOFRANGINGAREAOFTHEHOOLOCKHOOLOCK
ATNANKANG,MT.GAOLIGONGBETWEENSPRINGANDAUTUMN
LI Xu*, DENG ZhongJian, ZHOU Wei, BAI Bing
(Faculty of Conservation Biology, Southwest Forestry University; Key Laboratory of Forest Disaster Warning and Control in Yunnan Province, Kunming, Yunnan 650224, China)
Nankang District of GaoliGong National Nature Reserves was taken as research place, and four-month duration tracing was carried out for the activity area of a Hoolock gibbon family group composed of a female, a male and a child. Seven full-day tracing data were obtained in spring, 5 full-day tracing data were obtained in autumn, and effective tracing duration achieved 176.0 hours. Gibbon had relatively fixed feeding area and roosting area, and activity region area of Hoolock gibbon in spring was only 45.9% of activity region area in autumn. The night roosting area and feeding area of Hoolock gibbon in spring were centered in the grass and fruit planting area. The night roosting area and feeding area of Hoolock gibbon in autumn were not centered in the grass and fruit planting area and are more scattered, which indicates that human strawberry cultivation does not affect distribution of Hoolock gibbon, but grass and fruit picking in autumn disturbs feeding and roosting of Hoolock gibbon.
2010-11-15
云南省教育厅科学研究基金项目(08Y0193);云南省野生动植物保护与利用重点学科支持项目(XKZ200904);西南林学院重点科研基金项目(200608Z)
*通讯作者,lixu.swfc@yahoo.com.cn
1000-5463(2011)02-0108-05
Q951
A
