亚砷酸钠致早期鸡胚胚胎发育毒性与基因甲基化的关系
2011-03-06夏红飞
宋 歌 崔 熠 夏红飞 马 旭*
1.北京协和医学院研究生院(100730);2.国家人口计生委科学技术研究所遗传优生中心
砷是一种普遍存在的环境污染物[1],砷化合物对胚胎发育毒性机制的研究是近年来的研究热点。胚胎发育是细胞和组织顺次迁移和分化的过程,各种不同类型细胞的细胞周期受到严格的调控,保证了各组织器官发育的协调。DNA甲基化状态可以改变 DNA的结构,影响顺式反应元件与转录因子的相互作用,进而影响基因的表达[2]。鸡胚是一种用于研究胚胎发育过程中重要基因功能的经典模型[3]。砷化合物对鸡胚胎有十分强的毒性,可引起胚胎早期死亡、影响胚胎生长发育直至引起畸形[4]。过去认为,砷的甲基化是一种解毒方式[5]和使致癌原失活的过程,近年来的研究则认为,砷在体内的甲基化过程可能对 DNA的甲基化造成干扰,继而引起致癌原对 DNA的损伤,所以砷的甲基化反应被认为是砷在体内的毒性效应的增强过程[6],目前对于无机砷致胚胎发育毒性与 DNA甲基化之间的关系尚存在很多争议[7,8]。在本研究以鸡胚为动物模型,利用亚砷酸钠(SA)诱导鸡胚发育畸形,分析SA对鸡胚基因组 DNA整体甲基化及受甲基化调节的相关基因的影响,通过补充甲基供体胆碱(CHO)进一步研究 SA致鸡胚发育畸形与 DNA甲基化的关系。
1 材料和方法
1.1 鸡胚处理
所用种鸡蛋为北京梅里亚维通实验动物技术有限公司生产的白来杭 SPF级种鸡蛋。为了模拟鸡胚胚胎发育的过程,将鸡蛋置于能自动倾斜的空气流通的孵箱里,孵化温度 37.8℃,湿度为 60%。鸡胚处理操作步骤见参考文献[9],发育时期见参考文献[10]。种鸡蛋钝端用打孔器打孔,用一次性无菌注射器于 Hhstage 6、8、12期分别注入 50μl生理盐水 (CON组 ),50μl(100M)SA(SA组 ),50μl(100μg/μl)SA和 50μl(100μg/μl)胆碱联合处理(SA+CHO组 ),50μl(100μg/μl)胆碱(CHO组)。收获 Hhstage 20期(72h)鸡胚,根据不同实验进行相应处理。
1.2 鸡胚生存能力检测
鸡胚生存能力间接通过 72h(Hhstage 19~20)鸡胚体重和鸡胚存活率来检测,收获时以心脏跳动为存活。独立处理 3批,每批 25只鸡胚,统计体重和存活率;对 72h鸡胚小心剥开蛋壳,挑取心脏跳动的鸡胚,剥离胚胎外血管网,胚胎用 Zeiss lumar V 12立体显微镜在白光下摄相,观察鸡胚形态学变化。
1.3 5-甲基胞嘧啶(5-mec)免疫荧光检测
收获 72h鸡胚置于 24孔培养板中,孔内加多聚
甲醛 -20°C固定过夜。第 2天将多聚甲醛换为100%甲醇,4°C过夜。第 3天用 75%、50%、25%梯度甲醇使胚胎复水,用 PBST洗净。10μg/ml蛋白酶K室温通透 25min后弃去蛋白酶 K,PBST洗 1次,时间 10min。接着用含 0.1%戊二醛的 4%多聚甲醛进行后固定,再用 PBST洗净,加 2%山羊血清封闭 30min后加入一抗(5-mec抗体用 PBS稀释 50倍,每孔 150μl),旋转摇床上孵育,4℃过夜。第 4天回收一抗,PBST清洗。加入二抗(FITC标记的山羊抗鼠 IgG用 PBS稀释 50倍),每孔 150μl。旋转摇床上室温孵育 2h,弃去二抗,PBST洗。胚胎用Zeiss lumar V 12立体显微镜 20×照相,488nm激发荧光,用 Axiovision Rel.4.8软件分析荧光强度。
1.4 Real-time PCR检测 bcl2、c-myc、cdkn2a、calb2基因 mRNA表达水平
1.4.1 cDNA合成 各处理组鸡胚采用 Trizol试剂提取总 RNA(详见试剂操作说明),紫外分光光度计测定总 RNA浓度。以总 RNA为模板,以寡核苷酸 Oligo(dT)为引物,按反转录试剂盒说明书进行反转录。
1.4.2 荧光定量 PCR 荧光定量 PCR反应体系(20μl)如下,分别用 bcl2、c-myc、cdkn2a、calb2和内参 GAPDH的引物 :cDNA 1μl,引物 1μl,SYBRGreen real-time PCRMasterMix 12.5μl,ddH2O 5.5μl。用ABI 7000 real-time PCR仪分析鸡胚 bcl2、c-myc、cdkn2a、calb2基因 mRNA在不同处理组中的表达情况。荧光定量PCR反应条件为:95℃10min;95℃15s;60℃30s共 40个循环。用 7500 Software v2.0.1软件测定每一个样品的 Ct值(cycles threshold),用相对定量法分别分析 bcl2、c-myc、cdkn2a、calb2基因 mRNA的表达水平。以 bcl2基因为例,具体方法如下:先计算每个样品的ΔCt值,ΔCt=Ct(bcl2)-Ct(GAPDH);以 CON为对照,计算不同样品与对照的ΔΔCt值,然后再用2-ΔΔCt方法计算各处理组与对照之比的 bcl2基因的相对表达量。

表 1 引物和扩增片段大小
1.5 统计分析
2 结果
2.1 SA及其与 CHO联合处理对鸡胚生存力的影响
如表 2所示,与 CON相比,SA组胚胎存活率和胚胎体重明显减少(P<0.01)。AS组与 AS+CHO组相比,CHO组与 CON相比,胚胎存活率和胚胎体重差异均无统计学意义。
2.2 各组鸡胚表型
由图 1可见,SA可诱发鸡胚发育畸形,主要表现为体型变小、后脑发育迟缓、小头等畸形。补充甲基供体 CHO,未明显改变鸡胚的发育异常。与 CON相比,单独给予 CHO未引起胚胎发育畸形。这些结果提示补充胆碱不能明显恢复胚胎的发育异常。
表 2 各组鸡胚存活率及体重情况(±s)
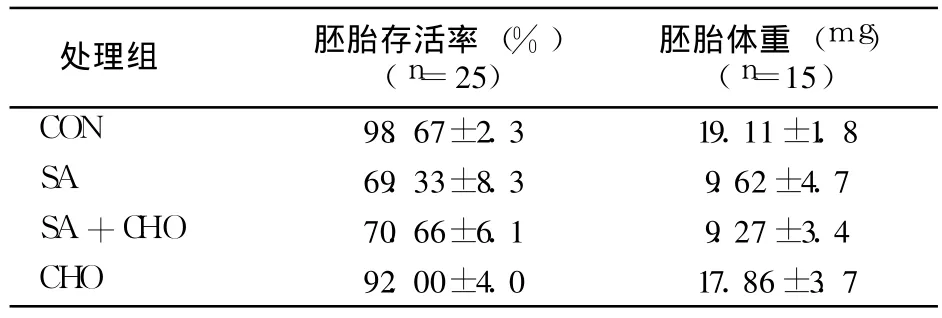
表 2 各组鸡胚存活率及体重情况(±s)
处理组 胚胎存活率 (%)(n=25)胚胎体重 (mg)(n=15)CON 98.67±2.3 19.11±1.8 SA 69.33±8.3 9.62±4.7 SA+CHO 70.66±6.1 9.27±3.4 CHO 92.00±4.0 17.86±3.7
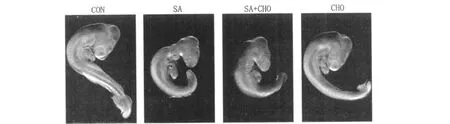
图1 不同处理组鸡胚形态学的变化
2.3 各组鸡胚基因组 DNA整体甲基化情况
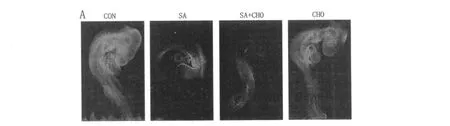
图 2 whole-mount免疫荧光检测 5-mec基因的表达情况
5 -mec含量是基因组 DNA整体甲基化水平的体现。利用全胚的免疫荧光来检测不同处理组鸡胚5-mec的水平。如图 2、图 3所示,与 CON相比,SA组 5-mec的含量明显减少(P<0.01)。SA组与CHO+SA组相比,CON与 CHO组相比,5-mec的表达无显著差异,提示补充 CHO不能明显拮抗砷抑制鸡胚基因组 DNA整体甲基化的作用。
2.4 不同处理方式对受甲基化调节基因表达的影响
为了研究鸡胚整体甲基化水平改变是否影响基因的表达模式,用荧光定量 PCR来检测受甲基化调节基因 bcl2、c-myc、cdkn2a和 calb2的表达水平。在利用 real-time PCR检测不同处理组受甲基化调节基因的表达前,首先检测了这些基因的溶解曲线和扩增曲线。这些基因的溶解曲线和扩增曲线均较特异,提示当前反应条件下,各基因均可得到很好的扩增。
由图 4可见,与 CON相比,SA组 bcl2(P<0.05)和 c-myc的 mRNA水平显著降低 (P<0.01),cdkn2a和 calb2mRNA水平没有明显变化(P>0.05)。与 SA组相比,CHO+SA组 bcl-2的表达水平增加(P<0.01),calb2的表达水平明显减少(P<0.05),c-myc和 cdkn2a的表达降低(P<0.01)。CON与 CHO组 bcl2、c-myc、cdkn2a和calb2的表达水平差异无统计学意义(P>0.05)。这些结果提示,尽管补充胆碱对砷抑制基因组 DNA整体甲基化水平没有明显的补偿作用,但在一定程度上可以抑制部分受甲基化调节基因的表达。
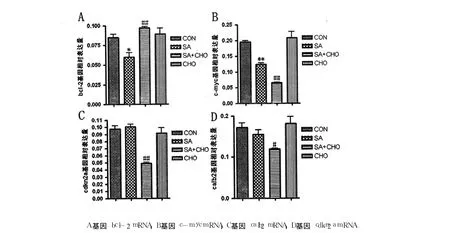
图 4 基因 bcl2、c-myc、cdkn2a和 calb2mRNA表达模式
3 讨论
在本研究中,SA能够显著降低鸡胚体重和存活率,并可诱发鸡胚各种发育畸形,主要表现为体型变小、后脑发育迟缓、小头等畸形。李勇等[11]研究了不同剂量砷诱发大鼠畸胎形成。Hood[12]给小鼠一次腹腔内注射砷酸钠,引起胚胎死亡,残存仔鼠体重减轻并出现多种畸形,以露脑为多见。本研究在鸡胚组织中的检测结果与这两篇文献在大鼠和小鼠中的报道基本一致。
无机砷在环境和机体内均有甲基化代谢的转换过程[13],砷暴露是否引起 DNA甲基化模式的改变,进而产生致畸效应,已引起人们极大的关注。本实验中,SA组鸡胚基因组 DNA整体甲基化水平低于对照组,说明砷处理组鸡胚基因组 DNA整体甲基化水平降低。这与 Nohara等[14]在小鼠中的研究结果是一致的,砷能够引起小鼠肝脏整体 DNA甲基化水平减少。此外,为了研究砷诱导的胚胎整体低甲基化与基因表达的关系,我们选择受甲基化调节,且在砷致胚胎毒性和细胞周期调控中发挥重要作用的基因 bcl2、c-myc、calb2和 cdkn2a[15,16]为研究对象,利用荧光定量 PCR分析这些基因在砷处理鸡胚中的表达模式,发现 bcl2和 c-myc表达下调,calb2和 cdkn2a没有明显变化,这些结果表明砷诱导的鸡胚基因组 DNA整体甲基化水平降低,同时,部分受甲基化调节的基因表达水平也具有下调趋势。
CHO是一种含丰富甲基的营养物质,做为甲基供体参与体内甲基代谢反应[17]。它能够影响胚胎发育,动物模型研究结果和人类流行病学研究表明,在妊娠期间补充 CHO对于胎儿中枢神经系统的发育和功能的正常是非常重要的[18]。叶酸的供甲基作用还能够参与维持 DNA甲基化状态,影响基因的表达,Ramirez等[19]报道指出叶酸能够抵抗砷的毒性。因为胆碱和叶酸都是一碳代谢的供体,已有研究证实 CHO缺乏导致基因甲基化的改变,并且间接影响细胞周期调控因子 cdkn、calb2的表达,以此改变胚胎大脑的发育[16];而 CHO能否拮抗砷的毒性还尚未见报道。为了研究 CHO是否可以拮抗砷的胚胎毒性,对 SA处理的鸡胚补充 100μg/μl CHO,观察鸡胚生存力和表型及甲基化水平的变化。研究发现,与砷处理组相比,SA和 CHO联合处理鸡胚不能明显改善鸡胚生存力和发育情况,鸡胚基因组DNA整体甲基化水平亦没有明显变化。检测受甲基化调节基因的表达发现 c-myc、calb2和 cdkn2a表达水平显著降低,这些结果提示尽管补充 CHO对砷抑制基因组 DNA整体甲基化水平没有明显的补偿作用,但可以在砷化物存在时抑制受甲基化调节的基因 c-myc、calb2和 cdkn2a表达,其机制有待进一步研究。
从以上结果可见,砷通过抑制基因组 DNA整体甲基化水平引起鸡胚发育异常,同时,部分受甲基化调节的基因表达水平存在反向关系。甲基化供体CHO不能明显恢复砷所致胚胎异常,但能抑制 cmyc、calb2和 cdkn2a的表达。对砷及其与 CHO联合处理对鸡胚甲基化影响分子机制的深入研究将有助于阐明基因甲基化在 SA致鸡胚胚胎毒性中的作用机制。
1 Coppin JF,Qu W,Waalkes MP.Interplay between cellular methyl metabolism and adaptive efflux during oncogenic transformation from chronic arsenic exposure in human cells[J].J Biol Chem,2008,283:19342-19350.
2 Doerfler W.DNA methylation and gene activity[J].Annu Rev Biochem,1983,52:93~ 124.
3 林艳青,耿建国.鸡胚早期发育过程中细胞周期调控的研究进展[J].广东药学院学报,2010,26(6):648-652.
4 万伯健,王长才.亚砷酸盐对早期鸡胚的毒性及其致畸作用的实验研究[J].环境与健康杂志,1984,1(4):3-4.
5 蔡林,王革娇.抗砷性微生物及其抗砷分子机制研究进展[J].微生物学报,2009,36(8):1253-1259.
6 Thomas DJ,Miroslav S,Shan L.TheCellular Meatbelism and Systemic Toxieity of Aesrnic[J].Toxicol Appl Pharmacol,2001,176(2):127-144.
7 Zhao CQ,MatthewR,Diwan BA,et al.Association of arsenic-inducedmalignant transformation with DNA hypomethylation and aberrant gene expression[J].Proc Natl Acad Sci USA,1997,94:10907-10912.
8 Mass MJ,Wang L.Arsenic alters cytosinemethylation patterns of the promoter of the tumor suppressor gene p53 in human lung cells:a model for amechanism of carcinogenesis[J].Mutat Res,1997,386(3):263-277.
9 Rosenquist TH,Ratashak SA,Sehlub J.Homocysteine induces congenital defects of the heart and neural tube:effect of folic acid[J].Proc Natl Acad Sci USA,1996,93:15227-15232.
10 Hamburger V,Ham ilton HL.A series of normal stages in the development of the chick embryo[J].JMorph,1951,88:49-58.
11 李勇,俞在芳.砷诱发细胞凋亡与大鼠畸胎形成的关系研究[J].卫生研究,1998,27(2):91-94.
12 Hood RD.Developmental effects of methylated arsenic metabolites in mice[J].Bull Environ Contam Toxicol,1998,61(2):231-238.
13 Vahter M,Marafante E.Intracellular interaction and metabolic fate of arsenite and arsenate inm ice and rabbits[J].Chem Biol Interact,1983,47:29-44.
14 Nohara K,Baba T,Murai H,et al.GlobalDNAmethylation in the mouse liver is affected by methyl deficiency and arsenic in a sexdependentmanner[J].Arch Toxicol.2010Oct 27.http://www.ncbi.nlm.nih.gov/pubmed/20978746
15 王文恭,童坦君.细胞凋亡过程中 bcl-2基因的甲基化[J].中国生物化学与分子生物学报,1998,14(3):309-313.
16 Niculescu MD,Craciunescu CN,Zeisel SH.Dietary choline deficiency altersglobal and gene-specific DNA methylation in the developing hippocampus ofmouse fetal brains[J].The FASEB Journal,2006,20(1):43-49.
17 Zeisel SH.Dietary choline:Biochem istry,physiology,and pharmacology[J].Annu Rev Nutr,1981,1:95-121.
18 Zeisel SH.Choline:Critical role during fetaldevelopment and dietary requirements in adults[J].Annu Rev Nutr,2006,26:229-250.
19 Ram irez T,Stopper H,Fischer T,etal.S-Adenosyl-L-methionine counteractsm itotic distrbance and cytostatic effects induced by sodium arsenite in HeLa cells[J].Mutat Res,2008,637:152-160.
