柠檬酸钠对L–异亮氨酸发酵及代谢流量分布的影响
2010-12-10程立坤徐庆阳
马 雷,程立坤,徐庆阳,陈 宁
(1. 天津科技大学电子信息与自动化学院,天津 300222;2. 天津科技大学生物工程学院,天津 300457)
L–异亮氨酸(Ile)是人体8种必需氨基酸之一,又是3种支链氨基酸之一,因其特殊的结构和功能而在人类生命代谢中具有特别重要的地位.L–异亮氨酸除用于一般营养型复合氨基酸输液,还大量用于配制治疗型特种氨基酸输液如肝安、肾安氨基酸输液,对治疗各种肝脏疾病具有显著疗效[1].随着 L–异亮氨酸市场需求量的增加,人们对发酵生产 L–异亮氨酸进行了深入研究.代谢流分析(Metabolic Flux Analysis,MFA)定量地描述了代谢网络中各个支路的流量分配关系,有助于分析目的产物的合成过程的瓶颈,对于理解细胞的代谢调控机制有重要意义.
常高峰等[2]通过对黄色短杆菌生产 L–异亮氨酸的发酵代谢分析得出,在 L–异亮氨酸的发酵过程中有缬氨酸、丙氨酸、亮氨酸及乳酸等杂酸积累,副产物的生成造成了碳源的浪费.在生物合成过程中,当EMP途径通量超出 TCA循环的代谢能力时,会使EMP途径生成的丙酮酸积累,则会导致酸及副产物的生成.有研究表明[3],在发酵过程中添加柠檬酸钠,可适当减少 EMP途径的代谢流量,降低副产物的生成.本文考察了添加不同浓度的柠檬酸钠对 L–异亮氨酸发酵的影响,旨在确定柠檬酸钠最适的添加量;并采用代谢流分析方法,定量地描述了柠檬酸钠添加前后黄色短杆菌(Brevibacterium flavum)WYJ1合成L–异亮氨酸代谢流的分布.这有利于采取相应措施,改造菌种及优化发酵过程控制,为进一步提高 L–异亮氨酸的得率提供理论依据.
1 材料与方法
1.1 实验材料
1.1.1 菌株与培养基
黄色短杆菌(Brevibacterium flavum)WYJ1(Met-+AECr+α-ABr)为天津科技大学代谢工程研究室保藏菌种.
培养基组成参见文献[4].
1.1.2 主要仪器
发酵罐(5 L、30 L全自动发酵罐),上海保兴生物设备工程有限公司;SBA–40C型生物传感仪,山东科学院生物研究所;pH电极和溶氧电极,Mettler Toledo;Agilent 1200型高效液相色谱仪,Agilent Technologies;DZF–6020型真空干燥箱,上海博迅实业有限公司;FA2204B型电子天平,上海精密科学仪器有限公司.
1.2 培养方法
斜面菌种活化培养:取斜面保藏菌种划线接种于活化斜面,31,℃恒温培养24,h.
5,L种子罐培养:吸取适量无菌生理盐水于 5支活化斜面中,将所有菌悬液接入 5,L种子罐中.初始通气量1,L/min,搅拌转速 300~600,r/min,通过自动流加氨水控制 pH(7.0±0.2),培养温度 31,℃,以泡敌消泡,培养10,h后,按10%接种量接入发酵培养基中.
30,L发酵罐培养:按 10%接种量将种子液接入30,L发酵罐中,初始通气量 2,L/min,搅拌转速400~800,r/min,通过自动流加氨水控制 pH(7.0±0.2),培养温度31,℃,以泡敌消泡,发酵培养65,h.
1.3 分析方法
菌体生物量:菌体生物量以菌体干重(g/L)表示,取10,mL发酵液,10,000,r/min离心20,min,将菌体用蒸馏水洗涤 2次后置于真空干燥箱中,80,℃干燥至恒质量,用分析天平称量.
溶氧及pH:在线测定.
葡萄糖和乳酸浓度:采用 SBA–40C型生物传感仪测定.
L–异亮氨酸及其他氨基酸含量:L–异亮氨酸及其他氨基酸含量采用高效液相分析系统测定.色谱分离条件:Agilent C18(150,mm×4.6,mm,3.5,µm)为色谱分离柱,2,4–二硝基氟苯柱前衍生测定,乙腈与NaAc溶液进行梯度洗脱,流动相流量 1,mL/min,检测波长360nm.
1.4 数据处理
发酵过程中菌体比生长速率 μ按照公式(1)计算:

式中:X、x均为菌体量;t为时间.用Origin绘图软件对实验数据进行微分计算,再用 Excel软件求解不同时刻的μ.
2 结果与分析
2.1 L–异亮氨酸生物合成途径分析
根据文献报道[5],在黄色短杆菌中L–异亮氨酸生物合成的中心代谢途径包括 EMP、TCA及 HMP途径,而乙醛酸途径封闭或微弱.L–异亮氨酸生物合成的代谢网络如图1所示.由图1可知,由葡萄糖合成L–异亮氨酸的途径较长且复杂.王健等[6]通过对 L–异亮氨酸的代谢途径分析得出:减弱 TCA循环和乙醛酸支路,可以使更多的碳架流转向异亮氨酸的合成.并且宋文军等[7]研究表明,在 L–异亮氨酸生物合成中,若 TCA 途径流量过大,会导致 L–异亮氨酸的代谢流减少.故减弱 TCA途径,可以提高发酵过程中 L–异亮氨酸产酸水平.Majewsi和 Domach[8]用限制性网络分析和相关酶活相关联的代谢流,指出在大肠杆菌和枯草芽孢杆菌中EMP途径相对于TCA循环要过量,当EMP途径通量超出TCA循环的代谢能力时,致使EMP途径生成的丙酮酸积累,且会通过其他途径进行代谢以缓解持续的“溢流”,导致酸及副产物的生成.有研究表明[3],在利用黄色短杆菌发酵生产 L–亮氨酸过程中,当 EMP途径流量超出 TCA及 L–亮氨酸生物合成的代谢能力时,丙酮酸代谢受阻,从而通过其他途径进一步代谢,导致副产物生成.因此,发酵生产L–异亮氨酸的过程中,减弱EMP途径代谢流量,降低副产物的生成及控制 TCA循环流量,是提高 L–异亮氨酸得率的关键.刘新星等[9]通过研究柠檬酸钠对枯草芽孢杆菌生产肌苷的影响得出,添加柠檬酸钠可以显著降低丙酮酸激酶和磷酸果糖激酶的活力,从而减弱 EMP途径通量.同时,柠檬酸钠会竞争性抑制柠檬酸合成酶的活性.因此,在L–异亮氨酸发酵过程中添加柠檬酸钠,可有效减弱EMP途径及TCA循环的代谢流量.
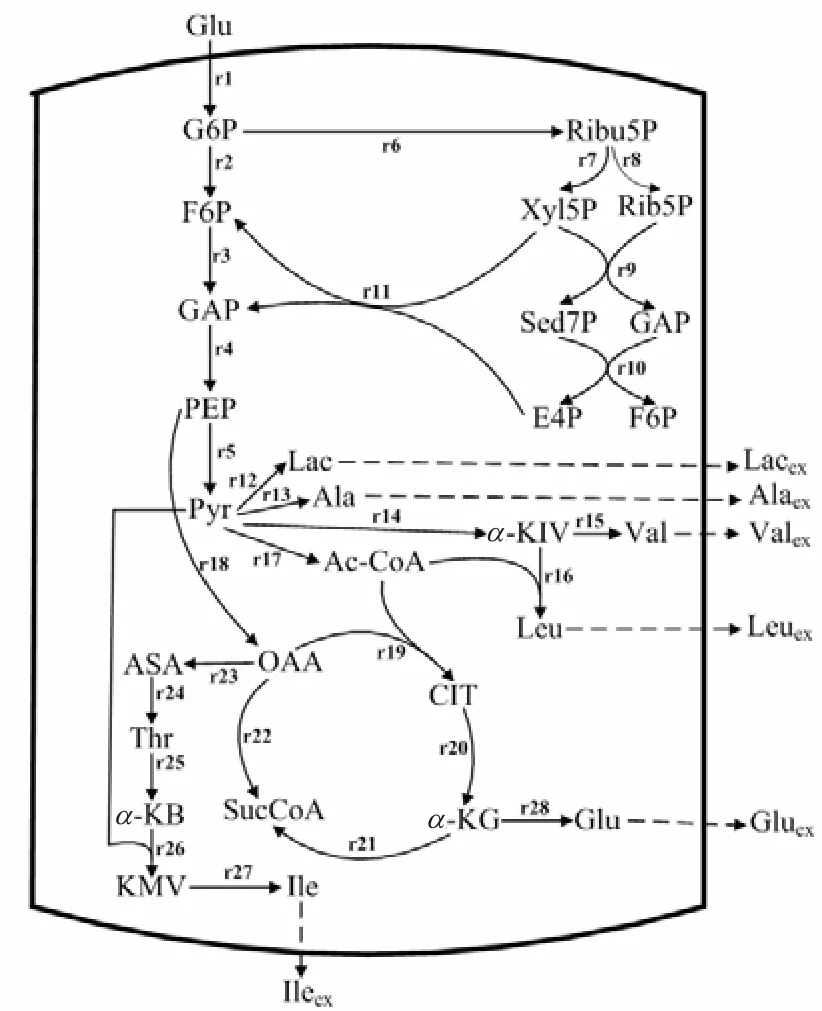
图1 黄色短杆菌L–异亮氨酸生物合成代谢网络Fig.1 Biosynthesis metabolism pathway of L-isoleucine in Fig.1 Brevibacterium flavum
2.2 柠檬酸钠对L–异亮氨酸发酵的影响
2.2.1 柠檬酸钠对发酵副产物的影响
宋翔等[10]研究表明,在谷氨酸发酵中添加柠檬酸钠可适当减少 EMP途径的代谢流量,降低副产物乳酸、L–丙氨酸、L–赖氨酸的生成.L–异亮氨酸属天冬氨酸族,又是分支链氨基酸,其生物合成存在严格的细胞内反馈调节抑制,不可能过量积累,且复杂的调控机制使得 L–异亮氨酸发酵产物中存在有机酸及其他氨基酸等副产物.发酵过程中副产物的生成造成了碳源的浪费及 L–异亮氨酸合成流量的减少.因此,减少副产物的生成可有效提高 L–异亮氨酸产率.分别向发酵培养基中添加 0.0、1.0、2.0、3.0、4.0 g/L 的柠檬酸钠进行 L–异亮氨酸分批发酵,发酵过程中副产物含量如图2所示.由图2可知,在L–异亮氨酸发酵过程中添加柠檬酸钠可有效减少副产物的生成,且随着柠檬酸钠添加量的增加,副产物含量逐渐降低,但是降低幅度逐渐减小.
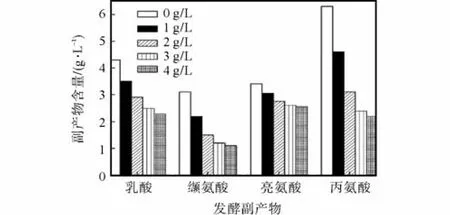
图2 柠檬酸钠对L–异亮氨酸发酵副产物的影响Fig.2 Effect of sodium citrate on byproducts of Fig.欢会 L-isoleucine fermentation
2.2.2 柠檬酸钠对菌体比生长速率及生物量的影响
柠檬酸钠对生物合成途径中关键酶如磷酸果糖激酶、丙酮酸激酶及柠檬酸合成酶等存在抑制作用,则在发酵过程中添加柠檬酸钠会抑制菌体生长[9].在L–异亮氨酸分批发酵中,柠檬酸钠对菌体生长的影响如图3所示.
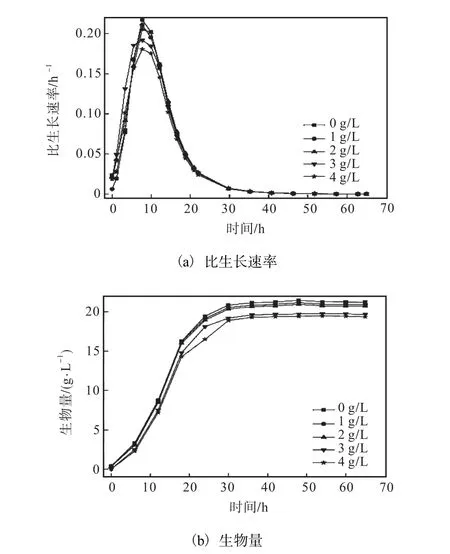
图3 柠檬酸钠对菌体比生长速率及生物量的影响Fig.3 Effect of sodium citrate on the specific growth rate Fig.3 and biomass
由图 3可知,柠檬酸钠添加量为 1.0、2.0,g/L与未添加时,菌体比生长速率及生物量差异不大.而添加量为3.0,g/L时,菌体比生长速率及生物量降低,且随着添加量的增加逐渐降低.
2.2.3 柠檬酸钠对L–异亮氨酸产量的影响
在发酵培养基中添加柠檬酸钠可减少副产物生成,即副产物的代谢流量减少可使 L–异亮氨酸的代谢流量增加.有研究表明[10],向L–谷氨酸发酵过程中添加柠檬酸钠,可增加 L–谷氨酸生物合成的代谢流,提高 L–谷氨酸产酸水平.而谷氨酸是异亮氨酸合成的前体物,则谷氨酸的增加利于 L–异亮氨酸的合成.发酵过程中添加柠檬酸钠对 L–异亮氨酸产量的影响如图4所示.
图4 柠檬酸钠对L–异亮氨酸产量的影响Fig.4 Effect of sodium citrate on the yield of L-isoleucine
由图4可知,当柠檬酸钠添加量为2.0,g/L时,产酸水平最高,为25.63,g/L,与未添加相比提高了 7.87%.但是柠檬酸钠添加量高于 3.0,g/L时,L–异亮氨酸产酸水平逐渐下降,这可能是柠檬酸钠对生物合成途径中关键酶的抑制作用造成的.
综上所述,当柠檬酸钠添加量为 2.0,g/L,可有效减少发酵过程中副产物的生成,且对 L–异亮氨酸生产菌的生长影响较小,同时使 L–异亮氨酸的产量得到提高.故在 L–异亮氨酸发酵中,柠檬酸钠的最适添加量为2.0,g/L.
2.3 柠檬酸钠添加前后代谢流量分析
在发酵培养基中添加或未添加 2.0,g/L柠檬酸钠,分别对其发酵中后期进行代谢流量分析.由图 3可知,L–异亮氨酸发酵过程中 36,h后菌体干重基本没有变化,说明发酵进入中后期.测定36~60,h葡萄糖、4 种氨基酸(L–异亮氨酸、L–缬氨酸、L–亮氨酸、L–丙氨酸)和乳酸的含量,并对其变化速率和代谢流进行计算,结果见表1.
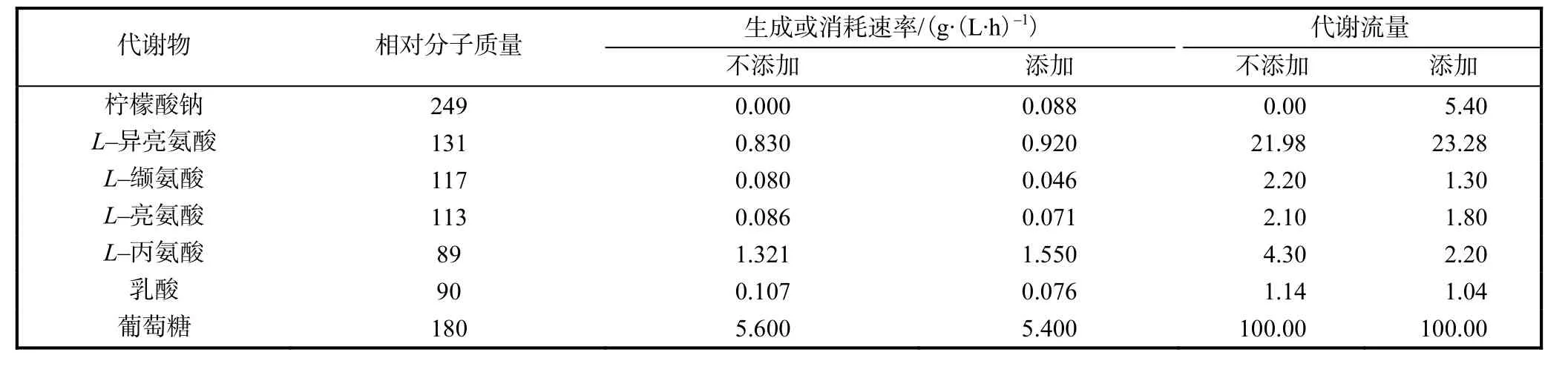
表1 柠檬酸钠添加前后代谢产物变化速率及代谢流量Tab.1 Variation rate and metabolic flux of metabolites without or with sodium ciltrate
由表 1可以看出,添加柠檬酸钠后,降低了副产物 L–缬氨酸、L–亮氨酸、L–丙氨酸及乳酸的代谢流量,提高了L–异亮氨酸的代谢流量.同时,加入柠檬酸钠以后,葡萄糖的消耗速率降低,这与文献[9]所得出的结论“柠檬酸钠降低了枯草芽孢杆菌的耗糖速率”相一致.但是,L–异亮氨酸的生成速率增加,这说明加入柠檬酸钠可以提高 L–异亮氨酸发酵中的糖酸转化率.
利用MATLAB软件中的Linprog函数计算得到柠檬酸钠添加前后 L–异亮氨酸发酵中后期的代谢流量分布,其结果分别如图5和图6所示.
由图 5和图 6可知,添加柠檬酸后生成副产物Val、Leu、Ala及 Lac的代谢流均有所降低,分别降低了40.91%、14.26%、48.84%、8.78%.而进入L–异亮氨酸生物合成途径的代谢流较未添加前提高了 5.91%.由于分支链氨基酸的合成需要谷氨酸提供氨基,则谷氨酸合成的代谢流量较大.未添加柠檬酸钠时进入HMP、EMP的代谢流量分别为 36.66、63.34,添加后HMP途径的代谢流量为 53.46,EMP代谢流量为46.54.则可知,添加柠檬酸钠可以减弱 EMP途径代谢流量,增加 HMP途径代谢流量,这与文献[3]报道的柠檬酸钠对产 L–亮氨酸黄色短杆菌代谢流分布的影响作用一致.HMP途径的主要作用是L–异亮氨酸合成途径提供还原力 NADPH.HMP途径流量增加,从而还原力 NHDPH 生成量增加,有利于 L–异亮氨酸的合成.柠檬酸钠添加后,TCA循环的代谢流量有所降低,由于柠檬酸钠合成酶是TCA循环的限速酶,添加柠檬酸钠会对柠檬酸钠合成酶活性产生抑制作用.因此,在 L–异亮氨酸发酵过程中添加柠檬酸钠,可有效减弱EMP途径及TCA循环的代谢流量,减少副产物的生成及提高 L–异亮氨酸合成的代谢流量,有利于L–异亮氨酸发酵.
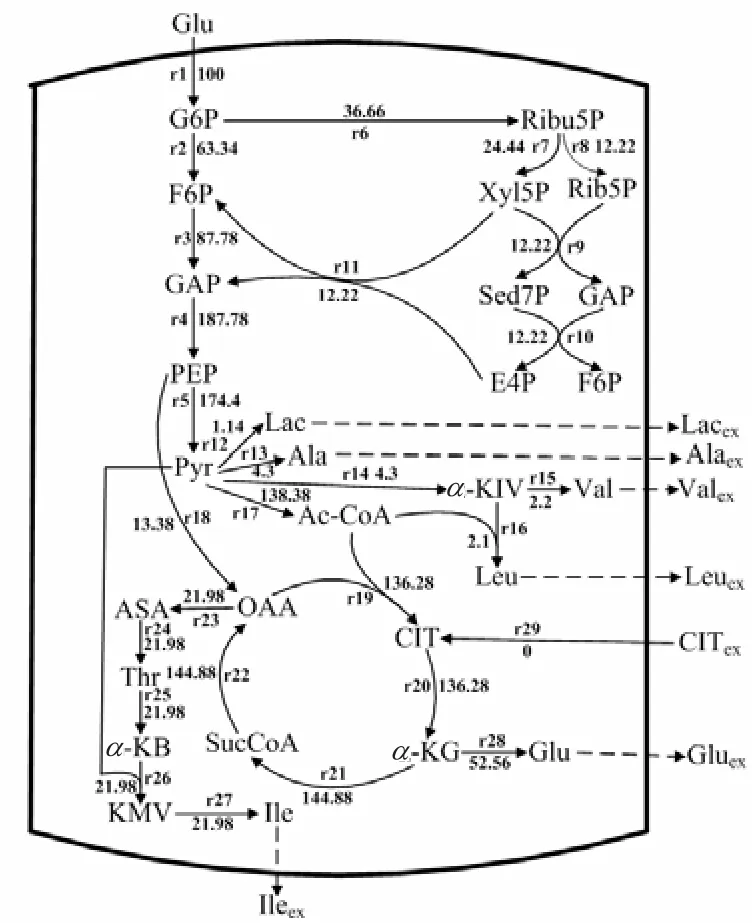
图5 未添加柠檬酸钠时L–异亮氨酸合成代谢流量分布Fig.5 Metabolic flux distribution of L-isoleucine fermen-Fig.5 tation without addition of sodium citrate
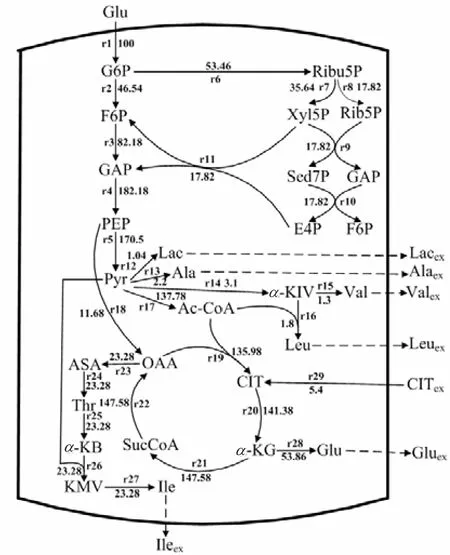
图6 添加柠檬酸钠后L-异亮氨酸合成代谢流量分布Fig.6 Metabolic flux distribution of L-isoleucine fermen-Fig.6 tation with addition of sodium citrate
3 讨 论
代谢网络是一个复杂的反应体系,要提高目的产物的产率,中心代谢途径与产物合成途径所组成的代谢网络的限速反应是关键.王健等[6]对 L–异亮氨酸的代谢途径进行分析,通过对基本模型的分析,确定了 L–异亮氨酸生物合成最优的代谢途径.通过引导最优途径中各个关键酶成比例的过量表达,在不破坏整个代谢途径的情况下,可提高 L–异亮氨酸产率,防止碳源的浪费.在代谢途径分析的指导下,通过优化培养基及控制工艺条件,使 L–异亮氨酸产酸水平得到提高.经酶学性质研究表明[9],柠檬酸钠的添加降低了EMP途径中关键酶PFK和PK的活力,从而引起糖酵解过程酶反应速率减慢,通量减少,同时 HMP途径的关键酶 6–磷酸葡萄糖脱氢酶的活力有所增加.则在 L–异亮氨酸发酵中添加柠檬酸钠可以减弱EMP途径,且可以增加还原力NADPH的供应,有利于提高 L–异亮氨酸的产率.本文利用代谢流分析法分析 L–异亮氨酸发酵中后期的代谢流量分布,可为菌株构建及发酵工艺研究提供理论依据,对 L–异亮氨酸规模化生产有一定指导意义.
[1] 宋文军,陈宁,魏春,等. 基于代谢流导向与分析的 L–异亮氨酸发酵条件优化[J]. 天津轻工业学院学报,2003,18(2):15-19.
[2] 常高峰,陈宁. L–异亮氨酸发酵代谢分析[J]. 生物技术通讯,2003,14(6):502-505.
[3] 陈宁,刘辉. 柠檬酸钠对 L-亮氨酸发酵代谢流分布的影响[J]. 高校化学工程学报,2008,22(3):478-482.
[4] 陈宁,常高峰,张克旭. L–异亮氨酸发酵培养基的响应面法优化[J]. 食品与发酵工业,2004,30(2):33-37.
[5] Pharkya P,Burgard A P,Maranas C D. Exploring the overproduction of amino acid using the bilevel optimization framework opt knock[J]. Biotechnol Bioeng,2003,84(7):887-899.
[6] 王健,王志诚,田梁逦霞,等. 产生 L–异亮氨酸的黄色短杆菌的代谢途径分析[J]. 生物技术通讯,2006,17(4):593-596.
[7] 宋文军,张克旭,张坤生,等. 不同供氧条件对 L–异亮氨酸合成代谢流的影响[J]. 中国食品学报,2003,3(1):1-6.
[8] Majewsi R A,Domach M M. Simple constrained-optimization view of acetate overflow in E. coli[J]. Biotechnology and Bioenineering,1990,35(7):732-738.
[9] 刘新星,陈双喜,储炬,等. 柠檬酸钠对枯草芽孢杆菌生长代谢及肌苷积累的影响[J]. 微生物学报,2004,44(5):627-630.
[10] 宋翔,谢希贤,徐庆阳,等. 柠檬酸钠对 L-谷氨酸发酵代谢流迁移的影响[J]. 天津科技大学学报,2009,24(2):5-8.
