长链非编码RNA GHRLOS2在结直肠癌中的表达及作用机制
2024-05-11褚菲菲孙志宁吴慧丽
冯 娇, 褚菲菲, 李 璐, 张 勇, 姜 珊, 孙志宁, 吴慧丽
(1. 新乡医学院, 河南 新乡, 453003; 2. 郑州大学附属郑州中心医院 消化内科, 河南 郑州, 450007)
结直肠癌是临床常见的消化系统恶性肿瘤之一,术后局部复发和远处脏器转移是患者死亡的重要原因。有氧糖酵解代谢重编程是恶性肿瘤的特征之一,指肿瘤细胞在不利环境中为了满足生长需求通常会改变自身代谢方式,从而促进肿瘤的发生与发展[1]。即使在氧气充足的情况下,肿瘤细胞代谢方式也会从氧化磷酸化(OXPHOS)重新编程为有氧糖酵解,这种现象被称为“Warburg效应”[2]。糖酵解途径很大程度上依赖于限速酶和葡萄糖转运蛋白,因此肿瘤生长、侵袭和预后不良与这些基因的失调有关[3]。糖异生是一种逆糖酵解途径,若激活会抑制代谢重编程,导致癌细胞中能量失衡和代谢应激。PCK1是糖异生过程中的关键酶之一,其异常表达会激活致癌基因相关通路,促进肿瘤的发生与发展[4]。长链非编码RNA(lncRNAs)是指长度超过200个核苷酸单位且无蛋白质编码功能的RNA转录本,其转录失调可参与癌症典型特征的形成,进而影响肿瘤的发生发展及对治疗的敏感性[5]。lncRNA是多种肿瘤的关键调节因子,包括结直肠癌[6]。GHRLOS为生长素释放肽的反义转录物[7], 是一种候选的lncRNA, 其分型GHRLOS2在结直肠癌中的作用机制目前尚不明确。本研究分析lncRNA GHRLOS2在结直肠癌中的表达水平及相关功能,并探讨可能的调控机制,以期为探寻结直肠癌的潜在治疗靶点提供参考依据。
1 材料与方法
1.1 一般资料
收集2021年9月—2023年3月郑州大学附属郑州中心医院收治的30例经病理学检查确诊结直肠癌患者的癌组织和对应癌旁组织标本,患者术前均未接受任何辅助治疗,标本采集后储存于-80 ℃冰箱中待检。本研究经医院医学伦理审查委员会审核批准(伦理批件号202242), 且所有患者知情同意。
1.2 主要材料
正常结肠上皮细胞株(NCM460)、结直肠癌细胞株(HCT116、SW480、RKO、LOVO、COLO205)、DMEM培养基和胰蛋白酶购自普诺赛生命科技有限公司; 胎牛血清购自美国Gibco公司; Trizol试剂购自美国Invitrogen公司,实时荧光定量聚合酶链反应(qRT-PCR)相关试剂盒购自日本Takara公司, miRNA荧光定量PCR试剂盒购自天根公司。PCK1、U6、GAPDH引物购自上海华大基因科技有限公司; PCK1抗体购自美国Abcam公司,二抗购自北京博尔西科技有限公司。
1.3 方法
1.3.1 细胞培养: 将正常结肠上皮细胞株、结直肠癌细胞株加入含有10%胎牛血清和1%双抗的完全培养基中,培养于37 ℃、5%CO2的培养箱。每隔1~2 d用磷酸盐缓冲液(PBS)清洗细胞,并更换培养基。当细胞充分贴壁且融合度达到80%以上时,使用胰蛋白酶消化后进行传代培养。
1.3.2 细胞转染: 将处于对数生长期的细胞铺于6孔板中,待6孔板细胞充分贴壁后使用PBS换液,每孔加入2 mL培养基,然后依次加入助转染试剂及慢病毒,于5%CO2、37 ℃培养箱中培养72 h, 观察转染效率。
1.3.3 数据挖掘: ① 从TCGA、GEO数据库下载结直肠癌组织和正常结直肠组织RNA测序数据; 应用R edgeR软件包(version 3.30.3)对所有测序数据和差异表达基因(DEGs)进行归一化分析。② 功能富集分析,从GEO数据库中下载编号为GSE134834的全基因组测序数据,该数据集共有10个样本(5例结直肠癌组织样本和5例癌旁组织样本)。利用GEO2R工具进行差异性分析,设置P<0.05, |log2(FC)|>1, 筛选得到DEGs(104个上调的DEGs, 82个下调的DEGs)。通过MetascaPe在线数据库对186个DEGs进行京都基因与基因组百科全书(KEGG)通路富集分析。③ 通过生物信息学软件(Targetscan)预测微小RNA-33b-5p(miR-33b-5p)与PCK1的结合位点。④ 通过人类蛋白质图谱(HPA)数据库分析PCK1蛋白表达水平。
1.3.4 qRT-PCR检测GHRLOS2和miR-33b-5p表达: 收集细胞,使用Trizol试剂从细胞或组织中提取总RNA。测定RNA浓度后,使用反转录试剂盒合成cDNA。参照SYBR Green qRT-PCR试剂盒和miRNA荧光定量PCR试剂盒的说明书配置反应液,然后放入PCR仪器中,设置反应的温度曲线,完成DNA扩增。qRT-PCR引物序列见表1, 采用2-△△Ct法计算相对表达量。

表1 qRT-PCR引物及序列
1.3.5 划痕实验检测过表达GHRLOS2对HCT116、SW480细胞迁移能力的影响: 转染后,将细胞接种于6孔板中。当细胞充分贴壁且贴壁面积达到80%~90%时,使用无菌的10 μL枪头以均匀压力垂直于6孔板底部划线。用PBS洗去悬浮细胞,更换无血清的新鲜培养液,随后将6孔板置于37 ℃、5%CO2培养箱中。0、12、24 h时,分别于显微镜下拍摄划痕区域照片,并应用Image J软件分析划痕距离。
1.3.6 Transwell实验检测过表达GHRLOS2对HCT116、SW480细胞迁移和侵袭能力的影响: 在无血清培养基中饥饿处理转染后的HCT116细胞和SW480细胞。待贴壁面积达到80%以上,收集细胞,使用离心机以1 000 r/min离心3 min, 弃上清液。使用无血清培养基重悬细胞,制备细胞悬液,计数后调整细胞浓度为1×105个/mL。将细胞均匀接种于Transwell小室的上室中(避免产生气泡),向下室中加入含有10%胎牛血清的培养基600 μL, 静置15 min后将细胞放置于培养箱中孵育48 h。孵育结束后,取出Transwell小室,吸走小室中的培养基,并用PBS小心清洗,用棉签擦除多余细胞。使用4%多聚甲醛固定细胞30 min, 以PBS洗涤2次后将小室底面浸入0.1%结晶紫中染色5~10 min。晾干后,使用显微镜观察并拍照,使用Image J软件进行计数。侵袭实验则需要将60 μL基质胶均匀涂布于Transwell上室底部,然后于培养箱中孵育3 h进行水化处理,其余步骤与迁移实验相同。
1.3.7 葡萄糖含量检测试剂盒检测过表达GHRLOS2细胞内葡萄糖含量: 收集转染后细胞的上清液,按一定比例加入蒸馏水,以超声波破碎细胞后置于沸水浴中煮沸10 min, 冷却后取上清液备用。参照说明书依次设定空白管、标准管和测定管,涡旋混匀,在恒温培养箱反应15 min, 于505 nm波长处检测吸光度。
1.3.8 蛋白质印迹法(Western blot)检测PCK1蛋白表达水平: 使用含有苯甲磺酰氟的RIPA裂解缓冲液裂解细胞,并提取每个样品的总蛋白。使用BCA试剂盒定量蛋白浓度。检测浓度和纯度后,将每个样品的总蛋白于100 ℃下变性10 min, 然后通过12%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分离等量蛋白质,并转移至聚偏二氟乙烯膜。于5%脱脂奶粉中封闭2 h后,在4 ℃条件下使用抗人PCK1一抗(1∶40 000)和β-actin一抗(1∶1 000)对膜进行探针检测。冲洗后,于室温下使用辣根过氧化物酶(HRP)标记的二抗处理膜2 h。使用ECL试剂(Beyotime)进行蛋白条带显影,并应用Image J软件进行定量分析。
1.4 统计学分析

2 结 果
2.1 GHRLOS2在结直肠癌组织及细胞中低表达
GEO数据库GSE39582数据集分析结果显示, GHRLOS2在结直肠癌组织中的表达低于正常结直肠组织,差异有统计学意义(P<0.05), 见图1A; GHRLOS2低表达与更高的肿瘤分期相关(P<0.05), 且提示更差的预后(P<0.05, HR>1), 见图1B、1C。
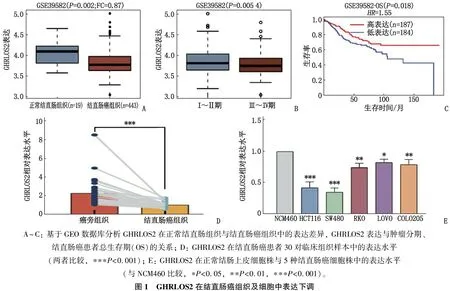
qRT-PCR检测结果显示,结直肠癌组织样本中的GHRLOS2表达水平低于癌旁组织,差异有统计学意义(P<0.001), 见图1D; 与正常结肠上皮细胞株NCM460比较, 5种结直肠癌细胞株中的GHRLOS2表达水平均降低(尤其是SW480、HCT116细胞),差异有统计学意义(P<0.05), 见图1E。本研究进一步分析30例结直肠癌患者的临床病理特征后发现, GHRLOS2表达与Grade分级、Stage分期、淋巴结转移均相关(P<0.05), 见表2。

表2 30例结直肠癌患者GHRLOS2表达与临床病理特征的相关性
2.2 构建稳定过表达GHRLOS2的结直肠癌细胞系
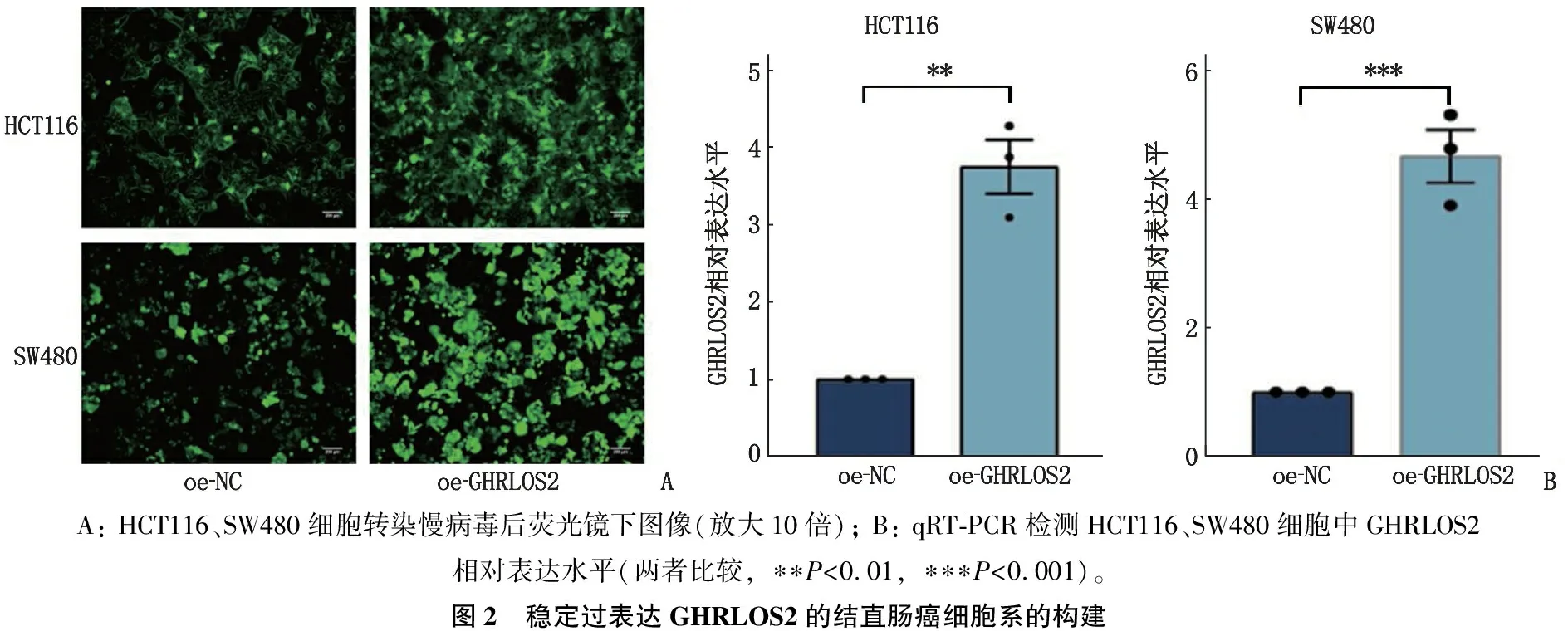
本研究选取GHRLOS2表达水平低的HCT116、SW480细胞,利用慢病毒载体构建稳定过表达GHRLOS2的细胞系。qRT-PCR检测结果显示,过表达GHRLOS2(oe-GHRLOS2)的HCT116细胞、SW480细胞的GHRLOS2相对表达水平均高于阴性对照(oe-NC)细胞,差异有统计学意义(P<0.05), 见图2。
2.3 过表达GHRLOS2抑制 SW480、HCT116细胞的迁移和侵袭能力
基于上述研究推测, GHRLOS2可能作为一种肿瘤抑制性lncRNA发挥作用。细胞划痕实验结果显示,与oe-NC细胞相比, oe-GHRLOS2细胞的迁移能力降低,差异有统计学意义(P<0.05), 见图3。Transwell实验结果显示,与oe-NC细胞相比, oe-GHRLOS2细胞的迁移能力、侵袭能力均降低,差异有统计学意义(P<0.05), 见图4。


2.4 过表达GHRLOS2抑制 SW480、HCT116细胞的糖酵解水平
应用GEO2R工具对GEO数据库结直肠癌相关数据集进行差异性分析,共发现186个DEGs(104个上调, 82个下调)。对186个DEG进行KEGG通路富集分析发现,糖酵解/糖异生信号通路相关基因显著富集,见图5A。葡萄糖含量检测结果显示, oe-GHRLOS2细胞的葡萄糖含量高于oe-NC细胞,差异有统计学意义(P<0.001), 见图5B。

2.5 GHRLOS2在结直肠癌中充当miR-33b-5p的分子海绵
lncRNAs的生物学功能高度依赖于其在细胞中的定位,本研究应用在线生物信息学工具(iLoc-LncRNA)分析后发现, GHRLOS2主要定位于细胞质(图6A),表明GHRLOS2参与基因表达的转录后调控。细胞质lncRNAs常作为竞争性内源性RNA(ceRNA)调节靶基因的水平,为了探讨GHRLOS2是否通过ceRNA模式调控CRC的进展,本研究基于starBase v3、LncBase v2、miRTarBase数据库分别进行miRNA-lncRNA及miRNA-mRNA互作关系分析,共得到8个候选的miRNA,见图6B。由于GHRLOS2可能通过ceRNA机制发挥功能,本研究进一步筛查GHRLOS2、8个候选miRNA和61个糖酵解相关基因的表达关系,最终识别出miR-33b-5p。此外,结直肠癌组织样本的miR-33b-5p表达水平高于对应的癌旁组织,差异有统计学意义(P<0.001), 见图6C。

2.6 PCK1是miR-33b-5p的下游基因
Targetscan软件预测结果显示,PCK1是miR-33b-5p的靶标,见图7A。TCGA数据库和GEO数据库(GSE21510、GSE39582数据集)分析结果显示,与正常结直肠组织相比,PCK1在结直肠癌组织中的表达降低,差异有统计学意义(P<0.05), 见图7B。结直肠癌患者30对临床样本检测结果显示,PCK1在肿瘤组织中的相对表达水平低于癌旁组织,差异有统计学意义(P<0.001), 见图7C。基于HPA数据库分析正常结直肠组织和肿瘤组织样本的PCK1蛋白表达情况,发现正常结直肠组织样本中PCK1蛋白呈中度染色,肿瘤组织样本中PCK1蛋白呈低度染色,提示结直肠癌组织中PCK1蛋白表达减少,见图7D。

2.7 GHRLOS2通过海绵化miR-33b-5p调控PCK1的表达
qRT-PCR、Western blot检测结果显示,与oe-NC细胞相比, oe-GHRLOS2细胞的PCK1表达水平升高,差异有统计学意义(P<0.05), 见图8A、8B。qRT-PCR检测结果显示,与oe-NC细胞相比, oe-GHRLOS2细胞的miR-33b-5p表达水平降低,差异有统计学意义(P<0.05)见图8C。由此提示, GHRLOS2通过miR-33b-5p调控PCK1的表达,进而影响糖酵解及细胞的侵袭、迁移能力,见图9。


3 讨 论
lncRNAs是重要的调控分子,与癌症的发生进展相关,可作为癌症诊断和预后评估的重要生物标志物[8-9]。lncRNA GHRLOS(生长素释放肽基因GHRL的反义链上发现的基因)编码的生长素释放肽具有抗炎作用,有助于预防肿瘤发生[7]。GHRLOS通过激活AMPK信号通路拮抗“Warburg效应”,进而抑制胃癌细胞的增殖、侵袭和迁移能力[10]。GHRLOS是miR-346的分子海绵,通过协同调节APC表达而抑制非小细胞肺癌的进展[11]。此外, GHRLOS还被证实为肿瘤转移的生物标志物及大肠癌的预后指标[12]。本研究发现, GHRLOS2在结直肠癌组织中的表达显著低于癌旁非肿瘤组织,且GHRLOS2表达与Grade分级、Stage分期和淋巴结转移相关。细胞功能学实验结果显示,过表达GHRLOS2可显著抑制结直肠癌细胞的侵袭、迁移及葡萄糖代谢,提示GHRLOS2在结直肠癌中可能发挥抑癌基因作用。
ceRNA假说认为, mRNA、lncRNA和环状RNA(circRNA)等RNA分子可通过共享的miRNA应答元件(MREs)靶向miRNA, 从而调控miRNA下游靶基因的表达[13-14]。在骨肉瘤中, KCNQ1OT1通过海绵化miR-34c-5p发挥ceRNA的作用,促进肿瘤进展和能量代谢重编程[15]。LINC00958通过海绵化miR-490-3p上调AURKA表达,从而促进膀胱癌的发生发展[16]。lncRNA的作用模式很大程度上与其亚细胞定位有关,其中细胞质lncRNAs通过充当miRNAs的分子海绵发挥作用。本研究基于在线生物信息学软件发现, GHRLOS2主要定位于细胞质中。此外, qRT-PCR检测结果显示, miR-33b-5p在结直肠癌组织中的表达水平较高,与GHRLOS2表达呈负相关,进一步支持了两者存在反向调控关系的推测。
研究[17]发现,在无线粒体缺陷的低氧条件下,乳酸发酵速率显著增加,这一现象被称为“Warburg效应”。“Warburg效应”可使细胞产生更多的乳酸,造成细胞外基质微环境酸中毒,为肿瘤细胞提供生长条件,并保护肿瘤细胞免受免疫系统的攻击,其被认为是癌症发生发展的关键驱动因素[18]。肿瘤从无氧糖酵解中获得能量,因此,作为糖酵解逆反应的糖异生或可成为有效的治疗靶点。PCK1是糖酵解逆反应糖异生的关键酶,不仅发挥代谢功能,如调节糖异生、甘油生成、丝氨酸生物合成及氨基酸代谢,还在糖尿病、肥胖症、癌症、心血管疾病、神经系统疾病中发挥重要作用[19]。PCK1在结直肠癌中低表达,并通过抑制UBAP2L蛋白丝氨酸454位点的磷酸化和增强自噬,抑制结直肠癌的进展[20]。本研究借助TargetScan软件成功预测了miR-33b-5p与PCK1之间的靶向作用,随后通过临床组织样本及相关细胞系验证发现, PCK1在结直肠癌中表达下调。本研究结果显示, GHRLOS2在结直肠癌中的表达水平与miR-33b-5p呈负相关,推测miR-33b-5p靶基因PCK1的表达可能受到GHRLOS2的间接调节。本研究基于过表达GHRLOS2实验发现, miR-33b-5p表达显著下调, PCK1表达显著上调,进一步提示GHRLOS2可能作为miR-33b-5p的分子海绵,介导对PCK1表达的调控作用。
综上所述, GHRLOS2作为一种新型的抑癌lncRNA,在结直肠癌组织及细胞中表达下调,其可通过发挥miR-33b-5p的分子海绵作用,上调PCK1表达水平,从而抑制结直肠癌的进展。GHRLOS2有望作为结直肠癌的新型生物标志物和潜在治疗靶点,为该疾病的临床筛查及靶向治疗提供新思路。
