昌岭山油松年内径向生长特征及其对气候的响应
2024-04-28贾飞飞孙茹李鑫孙红月董燕王杰
贾飞飞, 孙茹, 李鑫, 孙红月, 董燕, 王杰
昌岭山油松年内径向生长特征及其对气候的响应
贾飞飞1,*, 孙茹1, 李鑫1, 孙红月1, 董燕1, 王杰2
1. 辽宁师范大学地理科学学院, 大连 116029 2. 祁连山国家级自然保护区管理局昌岭山自然保护站, 武威 733104
利用树木径向生长记录仪对甘肃昌岭山油松径向生长进行了连续三年动态监测, 通过平均值法提取了油松净径向生长数据, 利用Gompertz函数对油松径向生长曲线进行拟合, 分析了油松年内径向生长特征, 并结合气象资料分析了油松径向生长对温度和降水的响应, 探讨了油松径向生长开始时间与温度的关系以及气候影响油松径向生长的机制。结果表明: (1)油松年内径向生长速率先增大后减小, 最大值出现5月下旬到6月上旬, 主要生长时间集中在5—8月。两棵油松径向生长开始时间较一致, 出现在4月上旬和中旬, 但结束时间存在较大差异。(2)生长期油松径向生长与平均气温、最高气温和最低气温均呈负相关, 与降水呈正相关, 但相关性强弱存在年际差异。(3)昌岭山地区油松径向生长开始的平均气温阈值在7 ℃左右。生长季温度会通过影响水分条件间接对油松径向生长造成影响, 异常充沛的降水可能是2019年6月两棵油松净径向生长量偏多的主要原因。
昌岭山; 油松; 径向生长; 气候因子
0 前言
森林生态系统是各种生物赖以生存和发展的基础, 对改善生态环境, 维持生态平衡起着不可替代的作用[1]。树木作为森林生态系统的组成部分, 其生长变化直接关系到整个森林生态系统的发展。已有研究表明, 随着全球变暖, 高温干旱胁迫导致一些地区树木死亡率普遍上升, 森林生态系统脆弱性增强[2–3]。因此, 全球变暖的背景下, 理解树木生长动态过程以及对气候的响应, 对于认识区域乃至全球森林生态系统的发展趋势具有重要意义。
树木径向生长记录仪可以对树木茎干生长进行动态监测, 具有分辨率高、连续性强的特点, 已经被广泛用于记录短期树木径向生长变化[4–7]。近年来, 在我国利用树木径向生长记录仪在树轮生态学和树轮气候学领域已经开展了许多研究, 并取得了一些成果。熊伟等[8]和管伟等[9]分别对六盘山地区南侧和北侧的华北落叶松的径向生长进行了监测和研究, 认为温度和降水是影响该区域树木径向生长的主要气候因子。牛豪阁等[10]和路明等[11]分别对祁连山东部的青杄和祁连圆柏径向生长动态研究, 发现影响树木径向生长的主要限制性因子是水分。肖生春等[12–13]利用树木径向生长记录仪对黑河下游胡杨生长季内径向生长进行了监测, 并结合环境气象、水文因子同步监测资料分析认为, 地下水和与之相关的土壤水分是制约黑河下游地区胡杨生长的最本质因素。受到树木种类以及立地条件的影响, 在不同环境下不同气候因子的重要性不同, 对树木径向生长的影响也有所差异。本文利用树木径向生长记录仪对昌岭山油松年内径向生长变化进行动态监测, 并利用监测数据和气象资料分析油松年内径向生长特征及其与气候因子的响应关系, 最后探讨气候因子对树木径向生长的影响。这不仅可以了解昌岭山地区树木生长规律, 对本区森林管理和保护也具有重要科学价值。
1 研究区概况
昌岭山位于甘肃省古浪县, 处于祁连山自然保护区昌岭山自然保护站内(图1), 属于祁连山脉的东部余脉。昌岭山总面积50.14 km2, 地势东高西低, 海拔2250—2900 m, 主峰海拔2954 m, 与基带高差达400—700 m, 北距腾格里沙漠仅8 km[14]。本区属于西北干旱区, 多年年平均气温为8.85 ℃, 气温日较差大, 多年平均降水量为188 mm, 降水主要集中在6—9月。林下土壤为灰褐森林土, 因海拔高度不同, 山地植被呈现出明显的垂直地带性, 由下至上依次为荒漠草原带, 落叶阔叶林带, 针叶林带以及灌丛带, 针叶林的主要建群种为油松(.)、祁连圆柏(.)和青海云杉()。
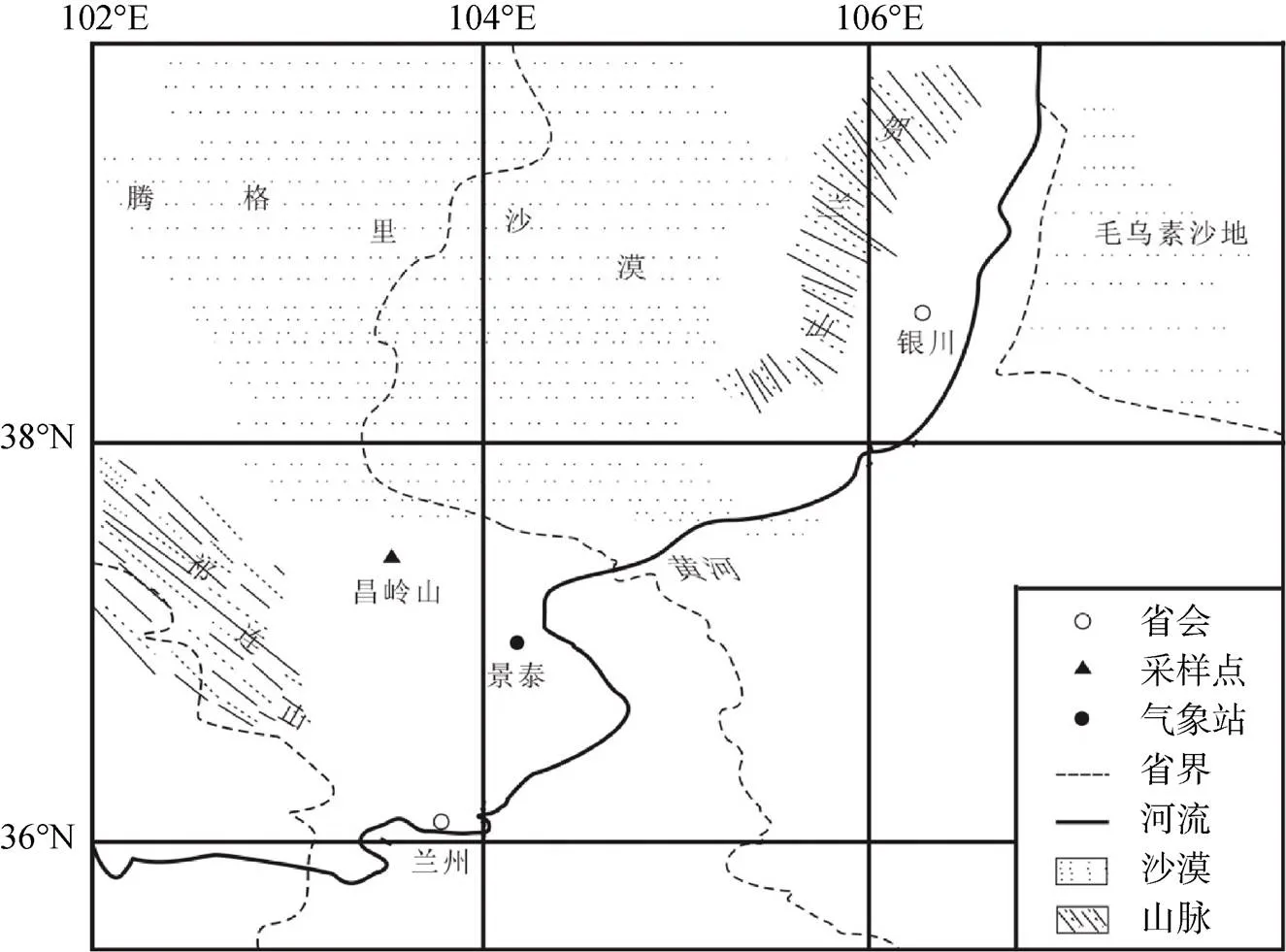
图1 昌岭山位置图
Figure 1 Location of Changling Mountain
2 数据和研究方法
2.1 油松径向生长监测
昌岭山大沟东北坡分布大片油松—青海云杉林,林中乔木以油松和青海云杉为主, 郁闭度0.7, 灌木层主要有铺地柏()、金露梅()等, 草本植物有冰草()、醉马草()等。通过年轮资料分析, 此处油松平均年龄在100年左右。2017年9月在昌岭山大沟东北坡海拔2400 m左右高度处选取生长状况良好的两棵油松(YS1和YS2), 在树干1.3 m处安装带状树木径向生长记录仪, 仪器型号为DRL26C, 可以同步记录气温数据。YS1和YS2树高分别为11 m、10 m, 胸径分别为93 cm、83 cm。树木径向生长记录仪每小时记录一次数据, 一天共记录24个数据, 监测数据时间区间为2017年9月24日11时到2020年10月31日10时。前期由于仪器故障, 记录的YS1径向生长数据出现异常, 因此YS1所使用的有效数据时段为2018年7月到2020年10月。
2.2 监测数据处理
树木径向生长记录仪记录的树木径向生长变化包括细胞吸水膨胀和失水收缩的可逆变化以及细胞分裂生长的不可逆变化, 为了获取树木净径向生长量, 需要从监测数据中剔除由于细胞吸水和失水等可逆变化带来的影响。常见的提取树木日净径向生长量的方法有平均值法[15]、最大值法[16]和周期法[17]。已有研究结果表明, 这三种方法提取的日净径向生长量具有较好的一致性[18]。因此本文选用其中的平均值法来提取油松日净径向生长量, 即求出每天24个数据的平均值, 然后用后一天数据均值减去前一天数据均值, 获取树木日径向变化量, 当所得数值为正时, 则认为是油松在这一天的日净径向生长量, 如果该数值为负值或零, 则认为当日树木未出现径向生长, 日净径向生长量标记为零。
Gompertz函数因其灵活性和形状的不对称性, 在描述树木径向生长模式方面具有独特的优势, 可以用来确定树木径向生长开始和结束的时间以及最大生长速率等[19–21]。本文利用Gompertz函数对两棵油松径向生长变化进行拟合, 并通过对函数求导得到油松生长速率曲线, 将生长速率大于4 μm·d–1的时间段作为油松的径向生长期[22], 进而确定两棵油松径向生长开始、结束以及最大生长速率所对应的时间。Gompertz函数表达式为:
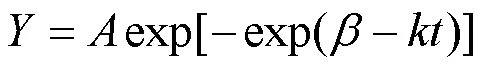
式中为树干径向生长的累积和;为以天为单位的时间;为内秉生长率;为与初始值有关的参数;为上渐近线。
2.3 气象数据
本文树木径向生长与气候的响应分析所使用的气象数据来自距昌岭山最近的景泰气象站(104.03°N, 37.11°E), 气候因子主要包括平均气温(Tmean)、最高气温(Tmax)、最低气温(Tmin)、降水(P)。图2为景泰气象站1957—2020年的多年月平均气温和降水情况。
3 结果与分析
3.1 油松径向生长特征
通过分析2018—2020年两棵油松各月净径向生长量可以发现, 不同年份的同一月份两棵油松净径向生长量差异较小, 说明监测期间两棵油松生长较为稳定, 未出现异常生长现象。同一年不同月份油松净径向生长量存在差异, 其中5—8月净径向生长量较大, 占年净径向生长量的60%以上(图3)。1月、11月和12月三个月份油松树干还存在净径向生长, 这可能是因为数据中还残留一些由于细胞吸水和失水带来的可逆信号, 但这几个月份净径向生长量值很小, 低于年净径向生长量的10%, 基本可以忽略这些可逆信号, 认为这几个月份油松树干未增长。两棵油松净径向生长量都在2019年6月出现了最大值, 生长量分别高达1760 μm和1923 μm (图3)。
由于2018年YS1监测数据时间序列不完整, 因此本文利用Gompertz函数仅对2019—2020年两棵油松的径向生长情况进行拟合, 拟合方程R2均达到了95.8%以上, 表明Gompertz函数能很好地表达研究区油松的径向生长情况。通过分析两棵油松径向生长速率, 并结合前文对树木径向生长开始和结束时间的定义, 发现不同年份两棵油松径向生长起止时间存在一定差异, 生长速率也不同(图4)。两棵油松径向生长开始时间出现4月上旬和中旬, 但结束时间差异较大, 2020年YS1径向生长结束时间最早, 在8月下旬(儒略日DOY239天)基本停止生长, 2019年YS2径向生长结束时间最晚, 在10月上旬(儒略日DOY280天)基本停止生长(表1)。2019年两棵油松径向生长开始时间都比2020年早, 结束时间比2020年晚, 生长期持续时间超过了170天。油松径向生长速率变化曲线成单峰型, 2019年两棵油松径向生长速率显著大于2020年, 年平均生长速率是20 μm·d–1, 最大生长速率均超过了40 μm·d–1。2019年和2020年两棵油松最大生长速率都出现在儒略日DOY160天左右, 即每年的5月下旬到6月上旬。
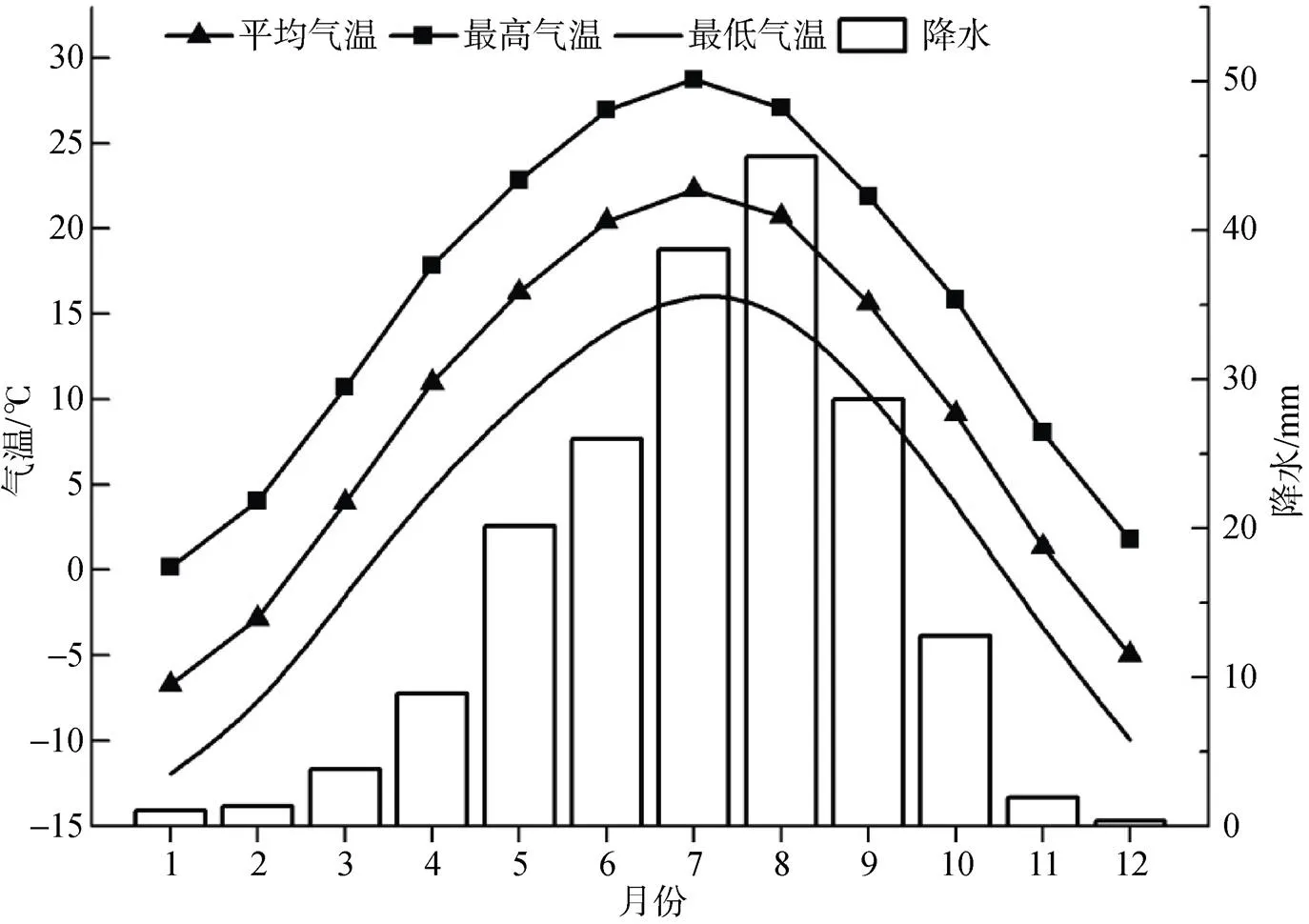
图2 景泰气象站1957—2020年月平均气温和降水分布
Figure 2 The mean monthly temperature and precipitation of Jingtai weather station from 1957 to 2020
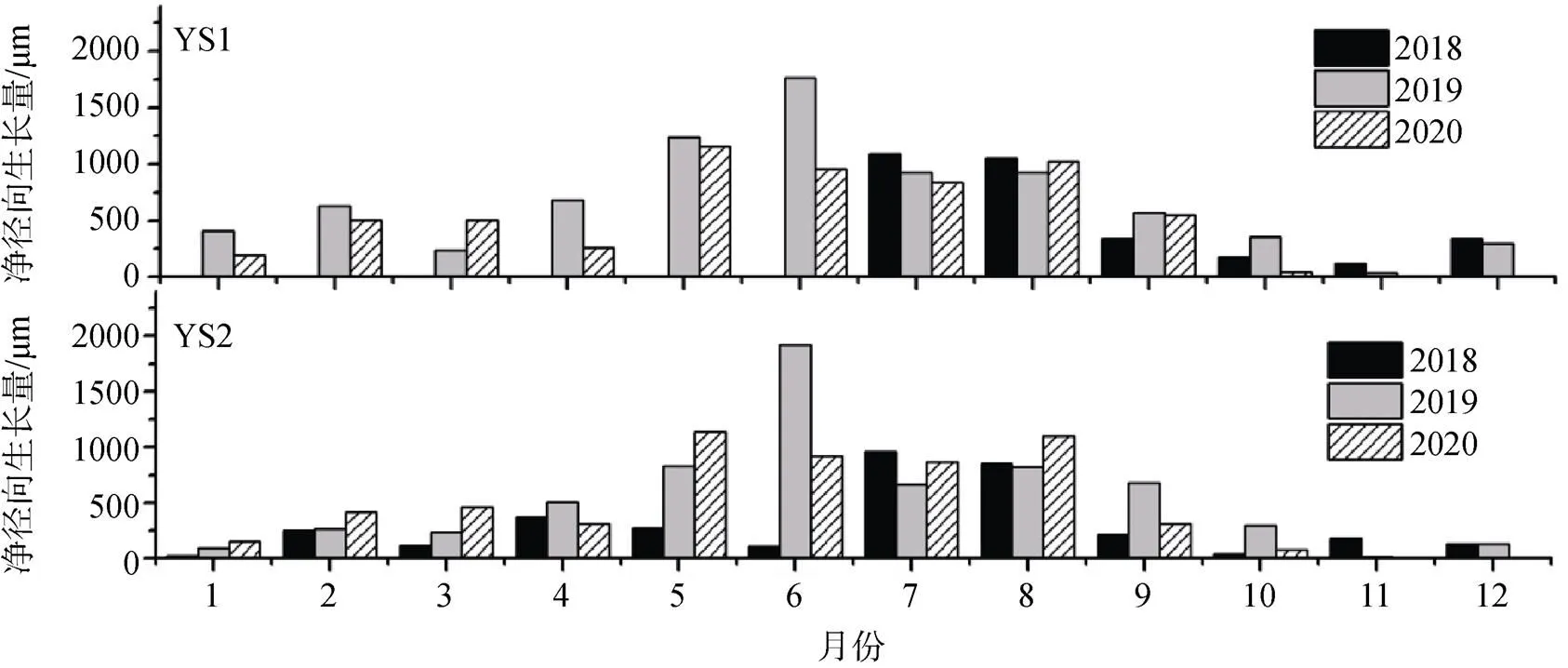
图3 2018—2020年油松各月净径向生长量
Figure 3 Monthly net growth offrom 2018 to 2020
3.2 油松径向生长对气候因子的响应
通过对两棵油松主要生长期(4—9月)日净径向生长量与气候因子的相关分析表明, 连续两个生长期油松日净径向生长量对气候因子的响应关系基本一致, 与平均气温、最高气温和最低气温均呈负相关, 而与降水呈正相关(图5)。2019年油松日净径向生长量与各气象因子的相关性明显高于2020年, 而且都通过了0.01的显著性水平, 特别是与降水的相关性, 2019年相关系数为0.355, 但2020年有所降低, 仅为0.124。2019年昌岭山地区年降水量多达到283 mm, 显著高于多年平均年降水量, 说明在湿润年份(2020年), 油松径向生长与降水的相关性会明显增强。
Figure 4 Radial growth fitting curves ofand growth rate in main growing season
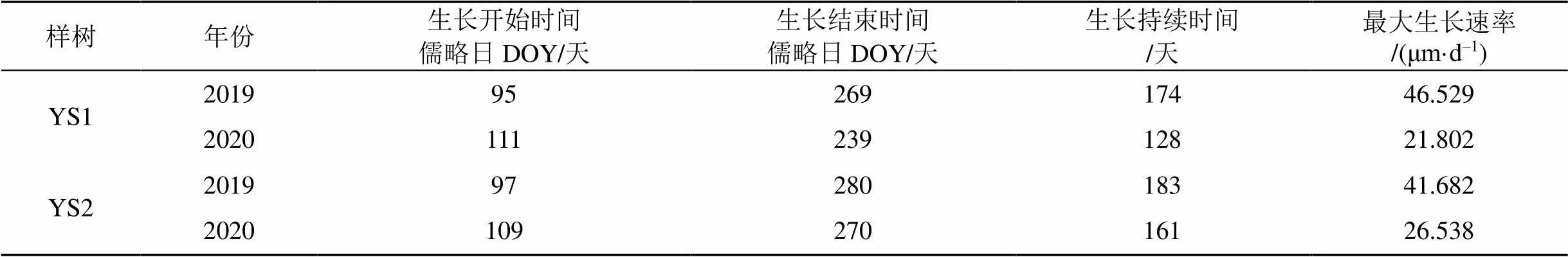
表1 2019—2020年油松生长情况信息表
4 讨论
4.1 油松径向生长开始时间与温度
对祁连山东部余脉昌岭山油松径向生长特征的研究发现, 昌岭山地区油松径向生长开始具体日期存在差异, 但总体都出现在4月上中旬。张军周[23]通过监测2013—2015年祁连山东部油松径向生长, 也发现油松径向生长开始时间出现在4月中旬。在贺兰山地区, 史江峰等[24]利用V-S模型判定了油松的主要生长期, 认为该区油松径向生长开始时间大约在4月22日左右。这些证据说明局域性油松径向生长开始时间比较一致, 大致出现在4月。
已有研究表明, 温度是影响温带树木生长周期主要因素[25]。关于树木生理学和物候学的研究也证实了温度在冬季休眠后形成层的再激活和木质部生长中的重要性[26–27]。当温度超过一定的阈值时, 树木形成层细胞才开始真正的生长, 即存在控制树木开始径向生长的温度阈值。Gao等[28]研究了贺兰山油松年内径向生长模式, 发现贺兰山油松在平均气温达到9.9 ℃时, 木质部开始生长。郭斌德等[29]在川西高原的研究结果显示, 7—9.5 ℃可能是川西高原林线附近冷杉形成层活动的温度阈值。Körmer和Paulsen[30]对世界各地林线位置的温度阀值进行了研究, 发现温带地区控制树木生长的平均气温阀值为7—8 ℃。昌岭山地区油松径向生长开始的时间是4月上中旬, 树木径向生长记录仪记录的这个时段本区平均气温在7 ℃±1.5 ℃, 这与其他地区的树木径向生长开始时间温度阈值接近, 说明虽然在不同地区树木的生长期长短存在差异, 但却存在较为接近的控制树木开始径向生长的温度阈值。
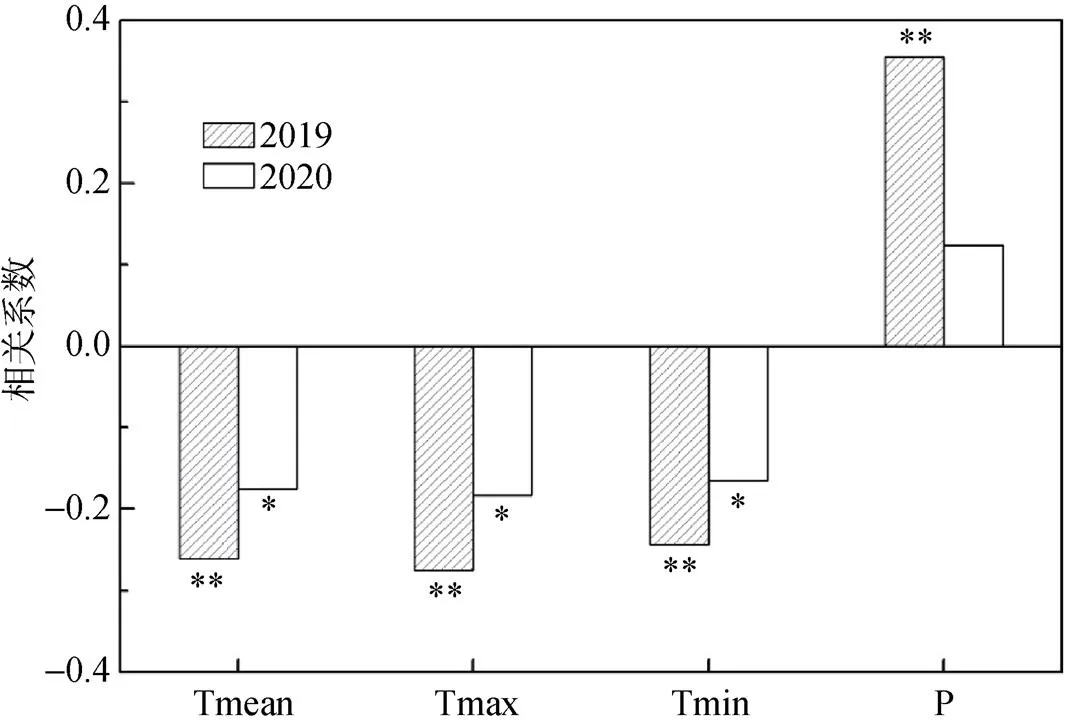
注:*表示达到0.05显著性水平, **表示达到0.01显著性水平。
Figure 5 The correlation coefficient between the radial growth ofand the climatic factors in the growing season
4.2 油松径向生长对气候响应机制
2019年和2020年油松主要生长期日净径向生长量均与平均气温、最高气温、最低气温呈负相关, 而与降水呈正相关。这与牛豪阁[31]在祁连山东部对青扦(Mast.)、油松和祁连圆柏三种针叶树种的研究结果一致。生长季温度过高或过低, 都会对树木的径向生长产生消极的抑制作用。温度过高会导致蒸腾作用加强以及光合作用效率降低[32],影响树木生长。最低温一般出现在夜晚, 最低气温过高会增强树木的呼吸作用, 从而加剧养分的消耗, 抑制油松的径向生长。白天最高气温过高也会使蒸发增强, 从而加剧干旱胁迫[33–34], 影响树木生长。生长季降水不仅可以为树木生长直接提供水分, 还可以增加土壤水分含量, 补充了因高温造成的水分缺失, 促进细胞分裂和增大, 对树木径向生长具有明显的促进作用[35]。2019年昌岭山地区降水量比正常年份降水多50%, 特别是6月降水量达到111 mm, 这可能是两棵油松净径向生长量最大值都出现在2019年6月的主要原因。
5 结论
本文利用树木径向生长记录仪对甘肃昌岭山两棵油松径向生长进行动态监测, 监测数据结果显示, 两棵油松年内生长趋势比较一致, 5—8月是它们的主要生长时期。受温度的控制, 两棵油松径向生长开始的时间也比较接近, 都出现在4月, 但生长结束的时间差异较大, 这与影响树木径向生长结束时间的因素更为复杂有关。研究区生长期油松径向生长量与降水和温度有相反的相关关系, 过高的温度会抑制油松的径向生长, 而丰沛的降水有利于油松的快速生长, 特别是主要生长期内的降水对油松年径向生长有重要贡献。基于以上研究结论, 可以初步推测, 全球变暖气候背景下, 极端降水或高温事件频发, 可能会造成研究区油松径向生长变得不稳定, 生长期各月径向生长量差异会增大, 这一认识对于保护区未来油松林管理以及预测油松林发展趋势具有很好地指导作用。
致谢: 感谢辽宁师范大学巩世钰和张建伟硕士在野外数据采集工作中的帮助, 特别感谢祁连山国家级自然保护区管理局昌岭山自然保护站工作人员在仪器维护工作中给予的帮助。
[1] 赵同谦, 欧阳志云, 郑华, 等. 中国森林生态系统服务功能及其价值评价[J]. 自然资源学报, 2004, 19(4): 480–491.
[2] PENG Changhui, MA Zhihai, LEI Xiaodong, et al. A Drought-induced pervasive increase in tree mortality across Canada's boreal forests[J]. Nature Climate Change, 2011, 1(9): 467–471.
[3] ALLEN C D, MACALADY A K, CHENCHOUNI H, et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests[J]. Forest Ecology and Management, 2010, 259(4): 660–684.
[4] DESKAYRIERS A, MORIN H, URBINATI C, et al. Daily weather response of((L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada)[J]. Trees, 2003, 17: 477–484.
[5] 王章勇, 杨保, 秦春, 等. 树木径向生长机制监测和模拟研究进展[J]. 中国沙漠, 2011, 31(3): 780–787.
[6] 李秧秧, 石辉, 张安邦, 等. 黄土丘陵区几种林木茎干径向生长的日变化及其对环境因素的响应[J]. 水土保持学报, 2007, 21(6): 170–173.
[7] 江源, 杨艳刚, 董满宇, 等. 芦芽山林线白杄与华北落叶松径向生长特征比较[J]. 应用生态学报, 2009, 20(6): 1271–1277.
[8] 熊伟, 王彦辉, 于澎涛, 等. 六盘山南坡华北落叶松()树干直径生长及其对气象因子的响应[J]. 生态学报, 2007, 27(2): 432–441.
[9] 管伟, 熊伟, 王彦辉, 等. 六盘山北侧华北落叶松树干直径生长变化及其对环境因子的响应[J]. 林业科学, 2007, 43(9): 1–6.
[10] 牛豪阁, 张芬, 于爱灵, 等. 祁连山东部青杄年内径向生长动态对气候的响应[J]. 生态学报, 2018, 38(20): 7412–7420.
[11] 路明, 勾晓华, 张军周, 等. 祁连山东部祁连圆柏()径向生长动态及其对环境因子的响应[J]. 第四纪研究, 2015, 35(5): 1201–1208.
[12] 肖生春, 肖洪浪, 司建华, 等. 胡杨()径向生长日变化特征分析[J]. 冰川冻土, 2010, 32(4): 816–822.
[13] 肖生春, 肖洪浪, 彭小梅. 黑河下游胡杨季节尺度径向生长变化研究[J]. 冰川冻土, 2012, 34(3): 706–712.
[14] 高尚玉, 鲁瑞洁, 强明瑞, 等. 140年来腾格里沙漠南缘树木年轮记录的降水量变化[J]. 科学通报, 2006, 51(3): 326–331.
[15] GRUBER A, ZIMMERMANN J, WIESER G, et al. Effects of climate variables on intra-annual stem radial increment in(L.) along the Alpine treeline Ecotone[J]. Annals of Forest Science, 2009, 66(5), 503.
[16] BOURIAUD O, LEBAN J-M, BERT D, et al. Intra-annual variations in climate influence growth and wood density of Norway Spruce[J]. Tree Physiology, 2005, 25(6): 651–660.
[17] DOWNES G M, BEADLE C, WORKEDGE D. Daily stem growth patterns in irrigatedandin relation to climate[J]. Trees, 1999, 14(2): 102–111.
[18] DESKAYRUERS A, ROSSI S, ANFODILLO T. Dendrometer and intra-annual tree growth: what kind of information can be inferred?[J]. Dendrochronologia, 2008, 25(2): 113–124.
[19] 李兴欢, 刘瑞鹏, 毛子军, 等. 小兴安岭红松日径向变化及其对气象因子的响应[J]. 生态学报, 2014, 34(7): 1635–1644.
[20] ZEIDE B. Analysis of growth equations[J]. Forest Science, 1993(3): 594–616.
[21] DUCHESNE L, HOULE D, D'ORANGEVILLE L. Influence of climate on seasonal patterns of stem increment ofin a boreal forest of Quebec, Canada[J]. Agricultural and Forest Meteorology, 2012(162/163): 108–114.
[22] WANG Zhangyong, YANG Bao, DESLAURIERS A, et al. Intra-annual stem radial increment response of Qilian Juniper to temperature and precipitation along an altitudinal gradient in Northwestern China[J]. Trees, 2015, 29(1): 25–34.
[23] 张军周. 祁连山树木形成层活动及年内径向生长动态监测研究[D]. 兰州: 兰州大学, 2018.
[24] 史江峰, 刘禹, 蔡秋芳, 等. 油松()树轮宽度与气候因子统计相关的生理机制——以贺兰山地区为例[J]. 生态学报, 2006, 26(3): 697–705.
[25] LȔTTGE U, HERTEL B. Diurnal and annual rhythms in trees[J]. Trees, 2009, 23(4): 683–700.
[26] ORIBE Y, FUNADA R, SHIBAGAKI M, et al. Cambial Reactivation in Locally Heated Stems of the Evergreen(Schmidt) Masters[J]. Planta, 2001, 212(6): 684–691.
[27] ROSSI S, MORIN H, DESLAURIERS A, et al. Predicting xylem phenology in black spruce under climate warming[J]. Global Change Biology, 2015, 17(1): 614–625.
[28] Gao J, Yang B, He M, et al. Intra-annual stem radial increment patterns of Chinese Pine, Helan Mountains, Northern Central China[J]. Trees, 2019, 33(3): 751–763.
[29] 郭滨德, 王晓春, 张远东. 阈值温度和积温对川西高原林线岷江冷杉径向生长的影响[J]. 生态学报, 2019, 39(3): 895–904.
[30] KÖRNER C, PAULSEN C K. A World-wide study of high altitude treeline temperatures[J]. Journal of Biogeography, 2004, 31(5): 713–732.
[31] 牛豪阁. 祁连山东部三种针叶树径向生长动态对气候的响应[D]. 兰州: 兰州大学, 2018.
[32] TARDIF, FLANNIGAN, BERGERON. An analysis of the daily radial activity of 7 boreal tree species, Northwestern Quebec[J]. Environment Assess, 2001, 67(1/2): 141–160.
[33] D'ARRIGO R D, KAUFMANN R K, DAVI N, et al. Thresholds for warming-induced growth decline at elevational tree linein the Yukon Territory, Canada [J]. Global Biogeochemical Cycles, 2004, 18, doi: 10.1029/ 2004GB002249.
[34] DRISCOLL W W, WILES G C, D'ARRIGO R D, et al. Divergent tree growth response to recent climatic warming, Lake Clark National Park and Preserve, Alaska[J]. Geophysical Research Letters, 2005, 32, doi: 10.1029/2005GL024258.
[35] 吕庚鑫, 徐军亮, 何存存, 等. 雪松年内径向生长节律研究[J]. 西北林学院学报, 2021, 36(2): 186–192.
Intra-annual stem radial growth characteristics ofand its response to climate on Changling Mountain
JIA Feifei1,*, SUN Ru1, LI Xin1, SUN Hongyue1, DONG Yan1, WANG Jie2
1. School of Geography, Liaoning Normal University, Dalian 116029, China 2. Changling Mountain Nature Reserve Station of Qilian Mountain National Nature Reserve Administration, Wuwei 733104, China
Global warming-related tree growth decline and mortality have been paid more attention around the world. To well understand the intra-annual stem radial growth characteristics ofand the respective roles of climatic variables on the tree intra-annual stem radial growth, the automatic dendrometer was used to continuously monitor the annual radial growth ofsin Changling mountain of Gansu province for three consecutive years. The net radial growth data ofwere extracted by method of mean value, and the radial growth curve was fitted by Gompertz function. Then the characteristics of intra-annual radial growth ofwere analyzed. The response of the radial growth ofto temperature and precipitation was analyzed based on meteorological data and monitoring data. Finally, the relationships between the beginning time of radial growth and temperature and the mechanism of climate affecting the radial growth ofwere discussed. The main conclusions are as follows. (1)The annual radial growth rate ofincreased and then decreased, and the maximum radial growth rate occurred in late May and early June, and the main growth time was from May to August. The beginning time of radial growth of the twowas consistent, which appeared in early and middle April, but the end time was different. (2)The radial growth ofwas negatively correlated with average temperature, maximum temperature and minimum temperature, and positively correlated with precipitation, but the strength of correlation was different between years. (3)The mean temperature threshold of the radial growth ofin Changling Mountain area was about 7 ℃. Temperature in the growing season might indirectly influence the radial growth ofby affecting water conditions. Abnormal precipitation might be the main reason for the excessive radial growth of the twotrees in June 2019.
Changling Mountain;; stem radial growth; climatic factors
10.14108/j.cnki.1008-8873.2024.01.020
Q149
A
1008-8873(2024)01-170-07
2021-09-11;
2021-11-25
国家自然科学基金项目(41601190)
贾飞飞(1988—), 女, 山东济宁人, 博士, 副教授, 主要从事树木年轮、古气候变化研究, E-mail: jiafeifei15@163.com
通信作者:贾飞飞
贾飞飞, 孙茹, 李鑫, 等. 昌岭山油松年内径向生长特征及其对气候的响应[J]. 生态科学, 2024, 43(1): 170–176.
JIA Feifei, SUN Ru, LI Xin, et al. Intra-annual stem radial growth characteristics ofand its response to climate on Changling Mountain[J]. Ecological Science, 2024, 43(1): 170–176.
