川西北高原不同海拔高度披碱草与垂穗披碱草核型分析
2024-02-26杨财容陈文婧王小琴祁伟亮叶美金刘松青
杨财容,陈文婧,王小琴,祁伟亮,叶美金,刘松青
(1.成都师范学院 化学与生命科学学院,四川 成都 611130;2.特色园艺生物资源开发与利用四川省高校重点实验室,四川 成都 611130;3.陇东学院 农林科技学院 甘肃 庆阳 745000)
披碱草(Elymus dahuricusTurcz.)与垂穗披碱草(Elymus nutansGriseb.)是禾本科小麦族披碱草属的多年生植物。披碱草性耐寒、耐旱、耐碱、耐风沙,是我国广泛栽培的优质高产饲草,分布于我国青海、陕西、新疆、四川、西藏、华北等地[1]。垂穗披碱草作为川西高原主要自然分布的优质牧草,是草原和草甸的主要组成成分,具有极高的饲用价值和水土保护能力,作为麦类作物的野生近缘种,其优异基因资源可用于麦类作物改良。目前关于披碱草属植物的染色体数目和核型前人均做过鉴定与分析,其报道的染色体的数目和倍性一致,即2n =6x = 42,但其报道的核型公式和类型却有一定的差别[2-4]。
居群是物种进化的基本单元[5],目前针对披碱草属居群基于形态标记与分子标记的遗传多样性研究较多。学者用分子标记的方法研究了披碱草居群的遗传多样性,发现披碱草22.64%的遗传变异发生在居群间[6]。前人利用形态学标记和分子标记的方法研究不同海拔高度的垂穗披碱草的遗传多样性,阐释了海拔对垂穗披碱草遗传多样性的影响[7]。海拔梯度差异往往会让光照、温度、湿度和土壤等一系列环境因素发生变化,是影响植物生物进化的重要原因之一,同时也与种内变异和进化有着密切联系[8-9]。严学兵[9]对垂穗披碱草进行的等位酶变异分析得出海拔是遗传变异最重要的因素,进行微卫星变异分析得出海拔是影响遗传变异的第二大因素。通过对不同海拔的3种披碱草属植物的表型测定,聚类分析显示聚为一类的多数种群有着相似的生境[7]。染色体是基因的载体,其形态结构的差异与变化包含着基因重排和基因数量的增减,从而影响着物种的进化[10]。张建波等[11]对川西北高原不同海拔共12个居群的垂穗披碱草进行了核型分析,结果显示12个垂穗披碱草居群的核型公式有5种,核型类型有1A、1B两类型,表明川西北高原不同居群野生垂穗披碱草在核型上发生了变异。
目前国产披碱草物种的核型已有报道,但从居群染色体核型层面探讨披碱草种内遗传多样性研究较少,结合环境因子进行相关研究的报道更是鲜见。本试验对分布于川西高原的不同海拔高度的披碱草和垂穗披碱草居群进行核型分析,可有效补充披碱草染色体层面遗传多样性研究,为披碱草的物种形成及演化提供新的细胞学证据。
1 材料与方法
1.1 实验材料
本研究中所有材料的居群编号及来源列于表1,披碱草6个居群于2018年8月采集于川西高原不同海拔地区,垂穗披碱草5个居群由四川农业大学小麦研究所提供,凭证标本保存于成都师范学院教学科研实训基地。
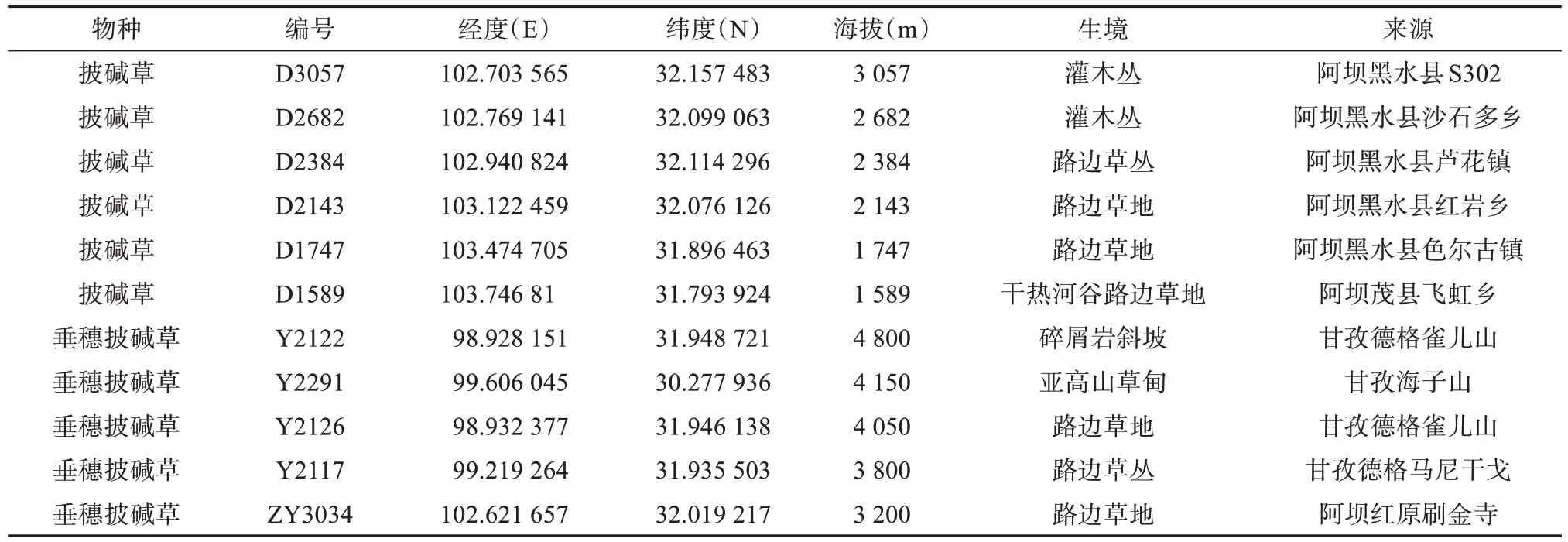
表1 材料来源Tab.1 Origin of materials
1.2 实验方法
1.2.1 根尖的获取
将每个居群3个单株的种子置于垫有湿润滤纸的培养皿上,4℃解除休眠后,置于25℃恒温培养箱中萌发,每隔24 h浇一次水。待根长至1 ~ 1.5 cm时剪下根尖。将剪取的根尖置于空的离心管内,置于N2O 中预处理2 h 左右,取出后用90%冰醋酸固定5 min,于70%酒精中保存。
1.2.2 滴片法制片
将于70%酒精中保存的3 ~ 5个根尖用蒸馏水清洗3 ~ 5次,用刀片切取分生区,置于20 μL混合酶解液(纤维素酶2%,果胶酶1%)中,在37℃水浴条件下酶解2 h。而后用70%酒精洗去酶液,重复操作2 ~ 3次,再加入少量70%酒精,捣碎根尖。6 000 r/min离心后自然晾干,滴加冰醋酸(30 μL每两个根尖),轻轻混匀后滴片。待载玻片自然干燥后滴加改良苯酚品红染液染色,盖上盖玻片在Olympus BX-51(Olympus)下观察并拍照。
1.2.3 核型分析
采用Levan等[12]的两点四区系统法确定染色体着丝点类型,按照Stebbins[13]的分类标准确定核型类型,同时参照李懋学和陈瑞阳[14]植物核型分析标准,核型不对称系数按照Arano[15]的方法计算。每个物种选取5 个分裂良好的细胞,利用Karyo-Type2.0进行染色体长度测量,利用该软件可直接输出染色体参数以及核型模式图。每个海拔梯度选取分裂相最好的一张照片作为模式照片,利用Adobe Photoshop 7.0.1软件对模式照片中的染色体进行处理,将模式图片中的染色体配对并进行排列,得到染色体核型图。
2 结果与分析
2.1 核型分析结果
披碱草6个居群、垂穗披碱草5个居群的染色体形态、核型图和核型模式图见图1-2,核型分析结果见表2,居群内未观察到核型差异。
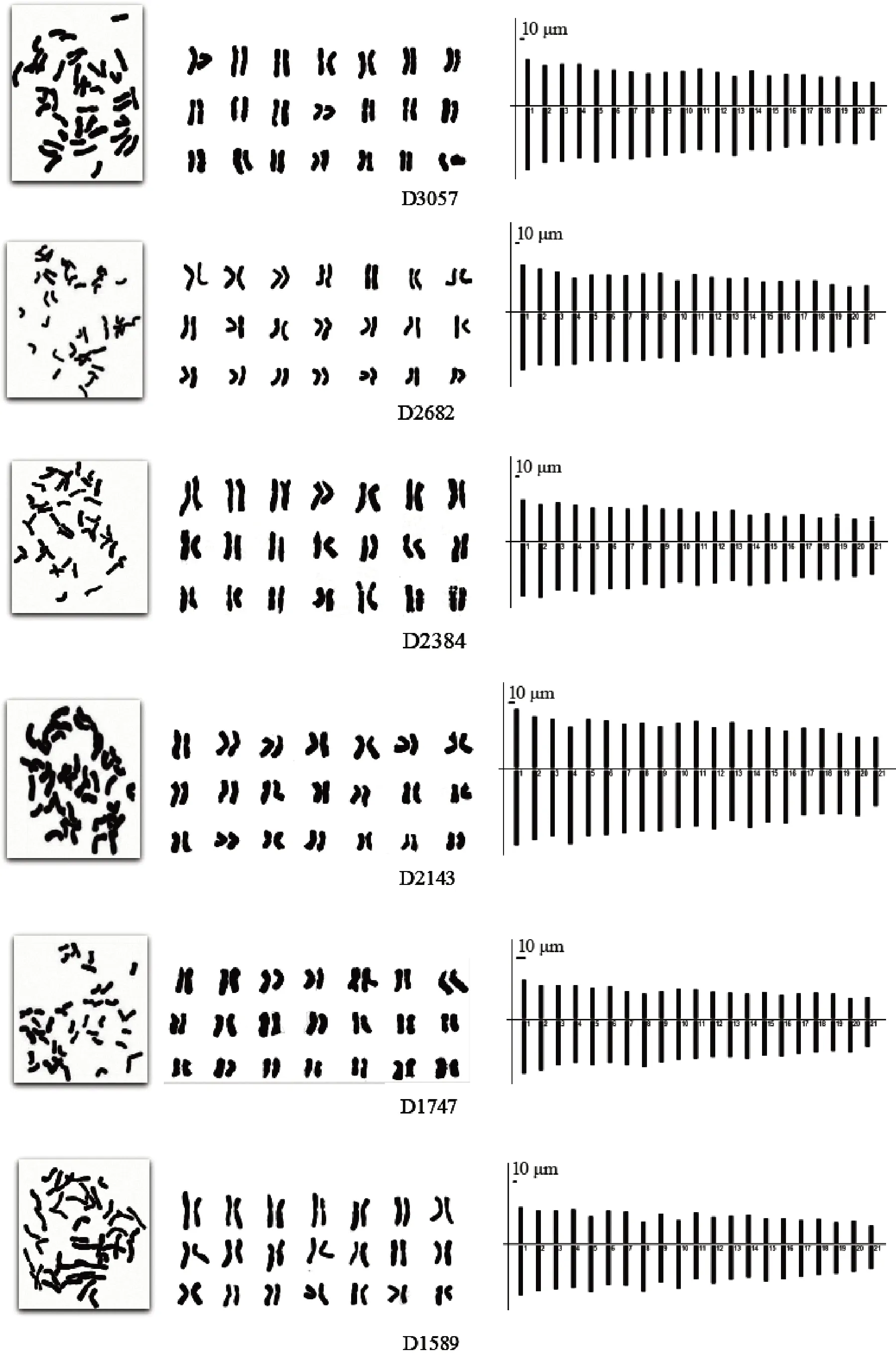
图1 各海拔梯度披碱草染色体形态、核型图、核型模式图Fig.1 Mitotic metaphase chromosomes, karyograms and ideograms of Elymus dahuricus from different altitudes
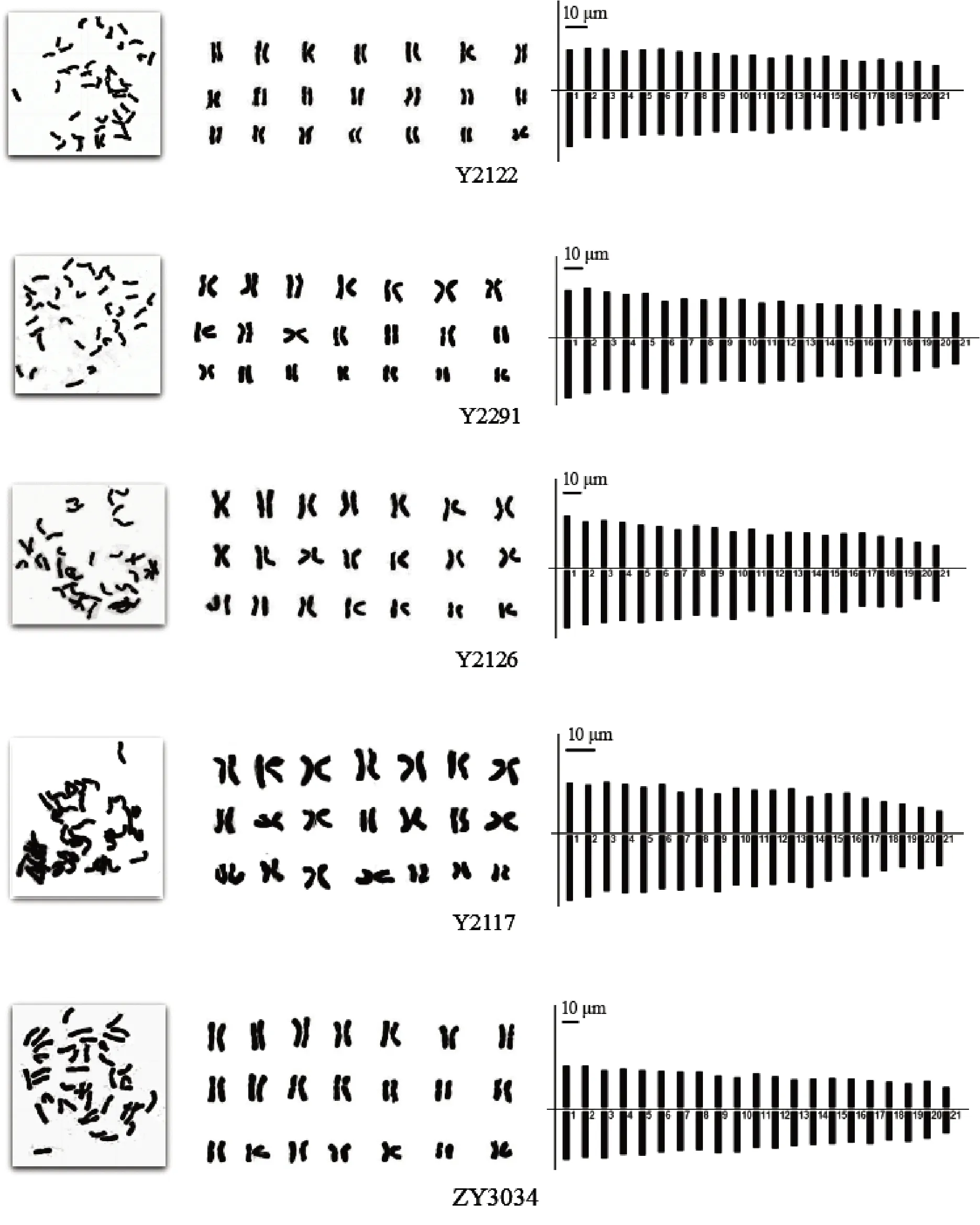
图2 各海拔梯度垂穗披碱草染色体形态、核型图、核型模式图Fig.2 Mitotic metaphase chromosomes, karyograms and ideograms of Elymus nutans from different altitudes
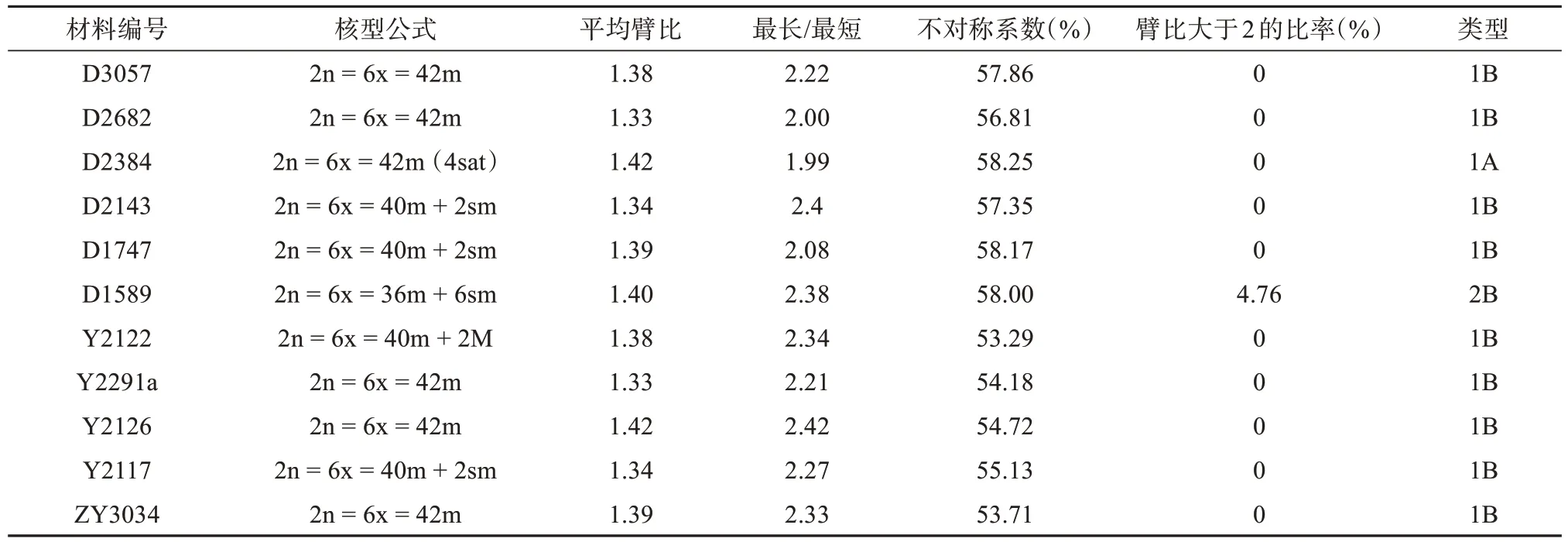
表2 各海拔梯度披碱草与垂穗披碱草居群染色体参数Tab.2 Parameters of karyotype of populations from different altitudes
披碱草D3057居群的核型公式为2n = 6x =42 = 42m,42条染色体均为中部着丝粒染色体,不具有随体染色体。染色体相对长度介于3.33% ~6.48%之间,染色体长度比为2.22。染色体的臂比范围是1.14 ~ 1.66,平均臂比为1.38,臂比值大于2的比例是0。染色体不对称系数为57.86%,核型类型为1B。
披碱草D2682居群的核型公式为2n = 6x =42 = 42m,42条染色体均为中部着丝粒染色体,不具有随体染色体。染色体相对长度介于3.14% ~6.33%之间,染色体长度比为2.00。染色体的臂比范围是1.12 ~ 1.59,平均臂比为1.33,臂比值大于2的比例是0。染色体不对称系数为56.81%,核型类型为1B。
披碱草D2384居群的核型公式为2n = 6x =42 = 42m( 4sat),42条染色体均为中部着丝粒染色体,有两对染色体有随体,位于第19、21对染色体上,且随体均位于短臂一端。相对长度介于3.40% ~ 6.23%之间,染色体长度比为1.99。染色体的臂比范围是1.17 ~ 1.65,平均臂比为1.42,臂比值大于2的比例是0。染色体不对称系数为58.25%,核型类型为1A。
披碱草D2143居群的核型公式为2n = 6x =42 = 40m + 2sm,42条染色体中有40条中部着丝粒染色体,2条为近中部着丝粒染色体,不具有随体染色体。染色体相对长度介于3.17% ~ 6.41%之间,染色体长度比为1.34。染色体的臂比范围是1.13 ~1.81,平均臂比为1.38,臂比值大于2的比例是0。染色体不对称系数为57.35%,核型类型为1B。
披碱草D1747居群的核型公式为2n = 6x =42 = 40m + 2sm,42条染色体中有40条中部着丝粒染色体,2条为近中部着丝粒染色体,不具有随体染色体。染色体相对长度介于3.34% ~ 6.52%之间,染色体长度比为2.08。染色体的臂比范围是1.19 ~1.91,平均臂比为1.39,臂比值大于2的比例是0。染色体不对称系数为58.17%,核型类型为1B。
披碱草D1589居群的核型公式为2n = 6x =42 = 36m + 6sm,42条染色体中有36条中部着丝粒染色体,6条为近中部着丝粒染色体,不具有随体染色体。染色体相对长度介于3.01% ~ 6.36%之间,染色体长度比为2.38。染色体的臂比范围是1.11 ~2.16,平均臂比为1.4,臂比值大于2的比例是4.76%。染色体不对称系数为58%,核型类型为2B。
垂穗披碱草Y2122居群的核型公式为2n =6x = 40m + 2sm,40条染色体为中部着丝粒染色体,其余2条染色体为近中部着丝粒染色体。染色体相对长度区间为2.79% ~ 6.17%,染色体长度比为2.27。染色体臂比区间为1.07 ~ 1.48,平均臂比为1.24,臂比值大于2的比例为0。染色体不对称系数为55.13%,核型类型为1B。
垂穗披碱草Y2291居群的核型公式为2n =6x = 42m,42条染色体均为均为中部着丝粒染色体。染色体相对长度区间为2.92% ~ 6.20%,染色体长度比为2.33。染色体臂比区间为1.03 ~ 1.34,平均臂比为1.16,臂比值大于2的比例为0。染色体不对称系数为53.71%,核型类型为1B。
垂穗披碱草Y2126居群的核型公式为2n =6x = 42m,42条染色体均为中部着丝粒染色体。染色体相对长度区间为3.02% ~ 6.49%,染色体长度比为2.21。染色体臂比区间为1.04 ~ 1.34,平均臂比为1.18,臂比值大于2的比例为0。染色体不对称系数为54.18%,核型类型为1B。
垂穗披碱草Y2117居群的核型公式为2n =6x = 42m,42条染色体均为中部着丝粒染色体。染色体相对长度区间为3.09% ~ 6.42%,染色体长度比为2.42。染色体臂比区间为1.06 ~ 1.39,平均臂比为1.22,臂比值大于2的比例为0。染色体不对称系数为54.72%,核型类型为1B。
垂穗披碱草ZY3034居群的核型公式为2n =6x = 40m + 2M,42条染色体均为中部着丝粒染色体。染色体相对长度区间为3.32% ~ 6.10%,染色体长度比为2.34。染色体臂比区间为1.02 ~ 1.36,平均臂比为1.15,臂比值大于2的比例为0。染色体不对称系数为53.29%,核型类型为1B。
2.2 核型与海拔高度的相关分析
根据核型分析结果,6个来自于不同海拔的披碱草居群中,居群D3057和D2682核型公式相同,居群D2143和D1747也具有相同的核型公式,但6个居群的平均臂比、最长/最短、不对称系数等参数存在差异,六个居群进化程度由高到低依次为:D1589(1 589 m) (2B) > D3057 (3 057 m)、D2682 (2 682 m)、D2173 (2 173 m)、D1747 (1 747 m) (1B) > D2384(2 384 m) (1A)。来自于海拔4 150 m的垂穗披碱草居群Y2291、4 050 m的Y2126和3 200 m的居群ZY3034具有相同的核型公式,但5个居群的平均臂比、最长/最短、不对称系数等染色体参数存在差异。从核型进化程度来看,5个不同海拔梯度的垂穗披碱草居群核型进化程度一致,其核型类型均为1B型。
3 讨论
3.1 不同海拔高度披碱草和垂穗披碱草居群的核型变异
前人针对披碱草属物种的核型研究报道较多[2-4,16],但从居群角度分析披碱草物种核型变异研究较少。核型分析的结果显示来自于不同海拔高度的披碱草和垂穗披碱草居群有着明显的核型变异,证明披碱草具有丰富的种内遗传多样性。本研究中不同海拔披碱草与垂穗披碱草居群染色体主要以中部着丝粒染色体为主,有少量亚中部着丝粒染色体。两个物种不同海拔居群间的核型参数(染色体相对长度,染色体长度比,染色体臂比,平均臂比,臂比大于2的比例)等均存在差异,核型公式也存在差异。张建波等[11]报道川西北高原12个不同海拔的垂穗披碱草居群的核型与本文报道结果存在差异,其报道核型类型有1A和1B两类,该研究中垂穗披碱草来源于四川甘孜与阿坝地区,海拔为3 300 m到4 200 m之间,其研究发现了更为丰富的核型变异,可能与其取样数量较多有关。任建东等[17]发现三江源地区的7份糙毛以礼草种质在不同类型的着丝点染色体数目、染色体有无随体上表现出丰富的多态性,核型类型也存在一定差异,与本文结果一致。Stebbins[13]认为,由于生境的不同,物种的染色体形态可以产生一定程度的变异,变异也是基因型的反应。物种遗传多样性的高低体现了物种对环境适应能力的强弱[18],川西北高原位于青藏高原东南部,平均海拔在3 000 ~ 4 000 m以上,低温、辐射大,气候条件恶劣,来自于不同海拔的披碱草和垂穗披碱草居群的丰富核型变异体现了其丰富的遗传多样性,增强了川西北高原披碱草环境适应能力。
3.2 各海拔梯度披碱草和垂穗披碱草的核型进化程度
核型进化是进化过程中最重要的方面之一,被认为是物种形成过程中的一种孤立机制,具有自身的进化趋势,与遗传进化无关[19]。核型不对称可以反映核型的进化,核型进化的基本趋势是从对称到不对称[13,19]。基于Stebbins[13]总结的核型类型的分类标准,划分出1A、1B、1C、2A、2B、2C、3A、3B、3C、4A、4B、4C共12种核型类型,其进化程度从1A ~ 4C依次增加,其中1A最对称最原始,4C最不对称进化程度最高。本研究中6个不同海拔的披碱草居群的核型进化程度由高到低依次为:D1589 > D3057、D2682、D2173、D1747 > D2384。川西高原5个不同海拔梯度的垂穗披碱草核型进化程度一致,其核型类型均为1B型,进化程度不高。基于徐克学[20]、钟扬等[21]的数量分类学和分支系统学理论,可计算出不同海拔居群的进化指数。不同海拔的披碱草居群的进化指数由高到低依次为:D1589 > D2384、D2143、D1747 > D3057 > D2682。不同海拔的垂穗披碱草居群的核型进化程度由高到低依次为Y2117 > Y2122 > ZY3034、Y2126、Y2291a。
进化指数的分析结果和基于Stebbins核型类型的分析结果存在差异,导致这种差异的原因可能是核型类型仅以臂比大于2的比例和染色体长度比来判断,而进化指数则还要参考不对称系数和平均臂比,究竟采用何种方法进行分析更加准确还需进一步研究。尽管分析方法不同导致结果有所差异,两种方法均显示来自于不同海拔高度的披碱草居群其进化程度存在差异,但川西高原披碱草居群染色体核型进化程度与海拔高度变化未呈现出一定规律性,原因可能在于染色体核型变异为海拔因素和其他多种生态因子长期综合作用的结果,未来可用其他分子生物学方法进一步研究披碱草遗传多样性与海拔高度或其他生态因子之间的关系[22-23]。
综上,来自于川西北高原不同海拔高度的披碱草与垂穗披碱草不同居群的染色体核型存在变异,可能与其环境适应能力有关,但是染色体结构层面究竟发生了何种变异还需借助其他方法进行进一步研究。
