光伏组件遮阴对喀斯特地区植物群落结构的影响
2024-01-11罗久富罗忠新吴赛男
罗久富,梁 松,袁 涛,罗忠新,刘 君,吴赛男,隋 欣
(1.中国水利水电科学研究院, 北京 100038;2.国家水电可持续发展研究中心, 北京 100038;3.国家电投云南国际电力投资有限公司, 云南 昆明 650228)
太阳能作为可再生和环境友好型能源,在改变能源消耗结构、替代化石能源的过程中具有极其重要的作用[1]。“协同推进可再生能源发展和生态保护修复”,探索可再生能源发展与西南石漠化区生态修复与保护发展体系在我国“十四五”特殊类型地区振兴发展中意义重大。石漠化区严苛的非生物环境在植物群落构建过程中发挥环境筛的作用[2],生境差异对植物群落结构演变具有明显的驱动效应[3]。光伏电站建设后,场区微生境差异对植物群落特征的影响机制成为“新能源 + 生态修复”工程的重要生态评价因子[4]。
光伏电站建设切割原生生境,改变场区局地气候、土壤等环境资源特征[5-7]。光伏组件遮挡可降低25%~95%的太阳辐射[8],而且,遮阴时长呈板下 >后檐 > 前檐[9]、光照强度呈前檐 > 后檐 > 板下[10]的规律。光伏组件对降水的再分配作用导致前檐下方土壤储水量高于组件下方和对照区[11],阵列间的土壤水分消耗速度明显高于组件下方[12],土壤水分可利用性降低。变化后的局地气候、土壤等环境资源导致原生植物群落结构发生适应性变化。在英国威斯特米尔太阳能公园(Westmill Solar Park)以及我国毛乌素沙地南缘、内蒙古羊草草原等地区,光伏组件遮阴导致组件正下方物种多样性低于阵列间或者前后檐[11,13-14]。此外,由于植物应对生境变化的生态策略的差异,不同植物功能群或种群响应组件遮阴的变化规律也存在差异。研究发现,太阳能公园组件正下方豆科和杂类草功能群优势度降低[14],青藏高原地区苜蓿(Medicago sativa)仅分布在光伏阵列间而香薷(Elsholtzia ciliate)则分布在组件下方[10],内蒙古羊草草原光伏组件中间缝隙的弱透光和汇水作用驱动正下方羊草(Leymus chinensis)的高度、密度、生物量显著增加[11]。在我国西南喀斯特区,太阳能资源开发是增强岩溶生态系统功能的重要环节,但是原本较为脆弱的喀斯特生态系统结构和功能易受光伏电站建设的影响,植物群落在维持生态系统功能中发挥着重要作用,因此,揭示喀斯特区光伏场区植物群落结构分异格局成为光伏生态修复亟待解决的科学问题。
本研究以云南东川光伏场区植物群落为研究对象,在前檐、后檐、板下3 类典型的组件遮阴区以及未遮阴的对照区开展群落学调查,探讨光伏组件遮阴后植物群落结构空间分异格局,为光伏场区植被恢复物种选择,构建高效、稳定的植物群落,创新“双碳”目标背景下西南喀斯特区植被恢复策略提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于云南省昆明市东川区“扶贫光伏”示范基地(103°08′31″ E,26°09′56″ N),海拔约1 280 m,基地所在东川区属于亚热带季风气候,年均降水量约1 000.5 mm,集中在5 月—10 月,月最大降水量208.3 mm[15]。最大风速40 m·s-1,多西南风,相对湿度76%。光伏电站所在区域年均太阳总辐射超过5 000 MJ·m-2,属于太阳能资源二类区。光伏电站建于2019 年,光伏组件依地势而建,前檐高约0.5 m,板下宽4.0 m,相邻阵列间距3.7 m 以上。场区无放牧活动干扰,为避免火灾每年进行地上植物刈割。场区内原生植被优势种为黄茅(Heteropogon contortus),伴生种为假杜鹃(Barleria cristata)、戟叶酸模(Rumex hastatus)、牛角瓜(Calotropis gigantea)等。
1.2 试验设计
于2021 年生长季选择平坦地势区域的光伏组件,在光伏组件低檐正下方(前檐,front eaves of the panel, FP)、中间正下方(板下,underside of the panel,UP)、高檐正下方(后檐,rear eaves of the panel, RP)以及无光伏组件遮阴的开阔区域(对照,Control,CK)设置东西走向条带样地,每个样地内布设5 个1 m × 1 m 样方,记录样方内出现的植物种及其盖度、高度、株丛数,所得单个样方内所有植物总株丛数为群落植株密度(plant·m-2)。所有样地重复3 次,随机分布在光伏场内避开边缘的区域。共调查12块样地,60 个样方(图1A)。
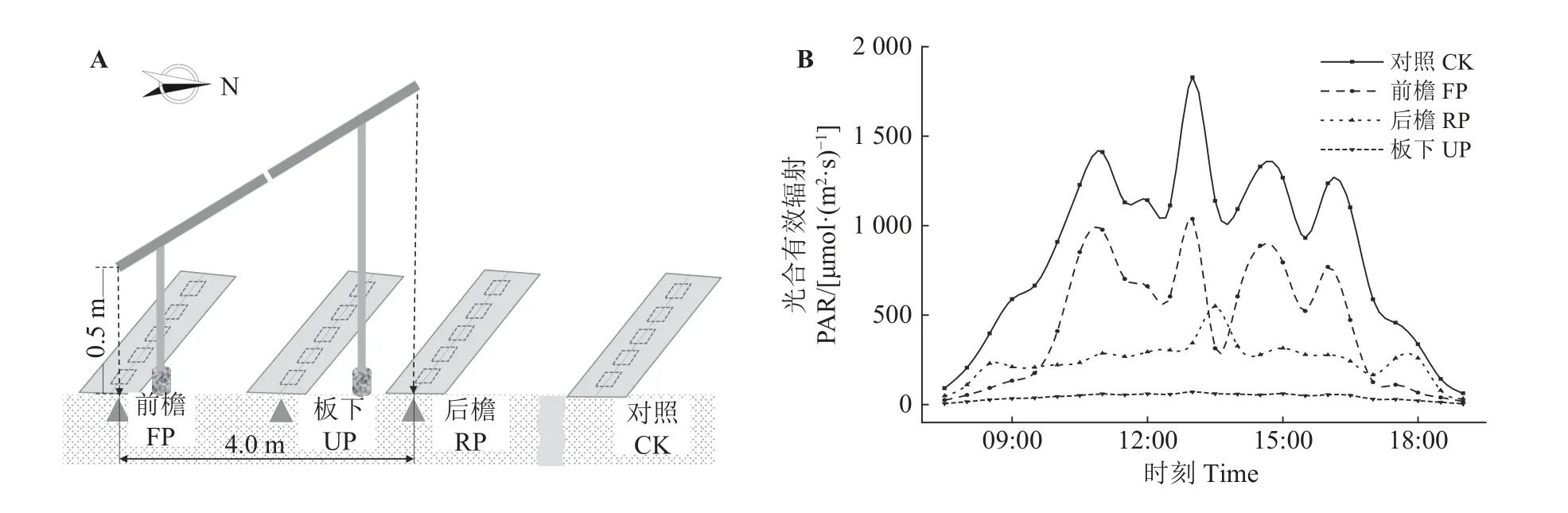
图1 光伏场区调查样地及样方布设示意图(A)及光合有效辐射(B)Figure 1 Spatial distribution of sample plots and quadrats in the photovoltaic arrays (A) and photosynthetically active radiation (PAR) values (B)
选择场区内地势平坦区域利用FM-G2A 光合有效辐射传感器监测前檐、板下、后檐、对照区域的植被顶层光合有效辐射,监测时间为8 月21 日—30 日,记录07:30—19:00 逐30 min 光合有效辐射值[photosynthetically active radiation,PAR,μmol·(m2·s)-1]。10 d 内PAR 日变化峰值分别为:对照1 828.43 μmol·(m2·s)-1,前檐1 037.57 μmol·(m2·s)-1,后檐550.14 μmol·(m2·s)-1,板下71.00 μmol·(m2·s)-1。PAR 日总量均值为,对照36.71 mol·m-2,前檐18.80 mol·m-2,后檐10.48 mol·m-2,板下1.82 mol·m-2。因此,光伏场区组件遮阴程度为板下 > 后檐 > 前檐 > 对照(图1B)。
1.3 数据计算与分析方法
1.3.1 物种重要值
1.3.2 功能群划分
根据物种生活型特征,将研究区植物分为以下5 类植物功能群(functional groups, FG):1)一年生丛生禾草(annuals bunch grasses, ABG);2)多年生丛生禾 草(perennial bunch grasses; PBG);3) 一 年 生 杂 类草(annuals forbs, AF);4) 多年生杂类草(perennial forbs, PF);5) 小灌木及半灌木(shrub and subshrub,SS)。
1.3.3 物种多样性指数
通过物种丰富度(richness,R)、Margalef 指数(M)、Simpson 指数(D)、Equitability 指数(E) 分析前檐、板下、后檐、对照区域草地植物群落物种alpha多样性,即:
以Jaccard 相似性指数(jaccard comparability index,J)分析前檐、板下、后檐、对照区域之间植物群落物种beta 多样性,即:
式中:S为调查样方中物种数;j为两个群落中共有物种数;a和b分别为两个群落中物种数;Pi为第i个物种的分盖度占样方物种盖度和的比例。
利用Excel 2016 对原始数据进行预处理,数据分析在SPSS 25.0 中实现,利用One-Way ANOVA 分析数据差异显著性,利用Origin 2016 制图。
2 结果
2.1 植物群落物种组成及植株密度特征
60 个样方中共调查到15 种被子植物,分属7 科15 属。以菊科和禾本科为主,分别包含5 种和4 种。随遮阴程度增加,黄茅重要值显著降低(P<0.05) (表1),后檐和板下白苞猩猩草重要值显著增加,白苞猩猩草替代黄茅成为板下群落优势种;赛葵和狗肝菜在后檐和板下群落中出现并成为优势种;后檐群落中假杜鹃重要值显著高于其他区域(P<0.05);而假苜蓿仅在对照和前檐群落中调查到,羽芒菊和鬼针草仅在前檐调查到,狗尾草仅在后檐调查到,艾纳香、马唐、酢浆草和破坏草则仅在板下调查到。组件遮阴后,植物群落发生明显的优势种替代和物种更替现象。

表1 光伏组件对植物重要值的影响Table 1 Effects of the photovoltaic panels on the species important values%
光伏组件遮阴改变群落植株密度(图2)。组件遮阴后,群落植株密度整体上呈先增高后降低趋势,对照和前檐群落植株密度显著高于后檐和板下(P< 0.05)。其中前檐群落中植株密度最大,比对照区群落高8.07 plant·m-2(P> 0.05);板下群落植株密度最小,仅68.00 plant·m-2,分别比对照、前檐、后檐低34.87%、39.54%和6.34%。
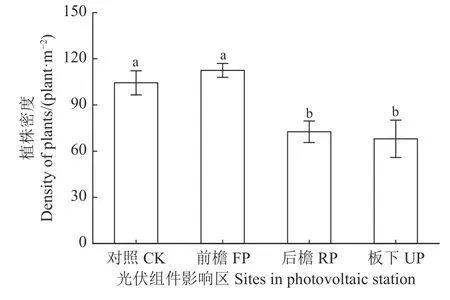
图2 光伏组件对植物群落植株密度的影响Figure 2 Effects of the photovoltaic panels on the density of plants
2.2 植物群落物种多样性特征
光伏组件遮阴增加植物群落物种多样性(图3)。随遮阴程度增加,群落物种丰富度、Margalef 指数显著增加。前檐、后檐和板下群落物种丰富度分别比对照区增加2.07、2.67 和3.27 species·m-2(P< 0.05),板下物种丰富度最高。光伏组件遮阴后,群落均匀度显著增加,后檐均匀度显著高于其他区域(P< 0.05);但遮阴程度增加显著降低群落优势度指数(P< 0.05),后檐和板下群落优势度指数显著低于对照和前檐。
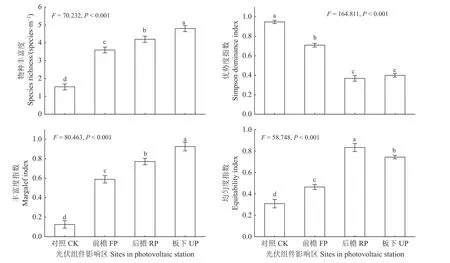
图3 光伏组件对植物群落alpha 多样性指数的影响Figure 3 Effects of the photovoltaic panels on the alpha diversity of species
随遮阴程度增加,前檐、后檐、板下与对照植物群落物种相似性指数降低,分别为0.50、0.38 和0.27(表2)。前檐与后檐、前檐与板下的相似性指数分别为0.36 和0.29,后檐与板下的相似性指数为0.42。遮阴程度差异越大群落之间物种相似性指数越低,共有物种数减少。
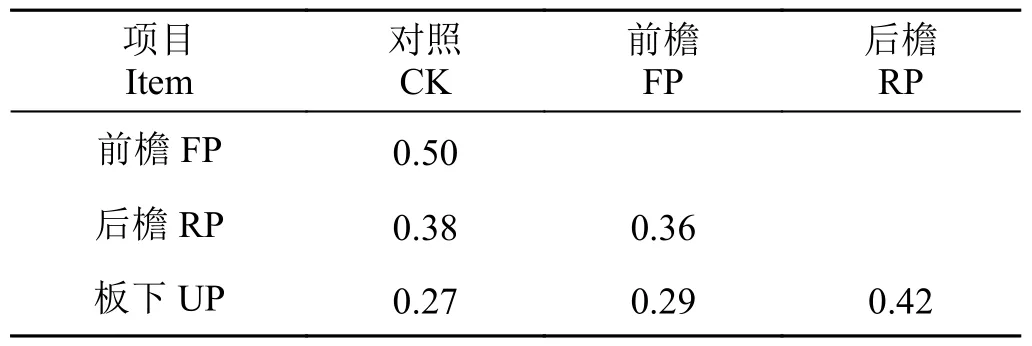
表2 光伏组件对地植物群落物种相似性的影响Table 2 Effects of the photovoltaic panels on similarity index among different sites
2.3 植物群落功能群特征
如图4 所示,在对照区仅存在多年生丛生禾草、多年生杂类草和小灌木3 种功能群。与对照相比,前檐和板下群落出现一年生丛生禾草和一年生杂类草,后檐群落中出现一年生丛生禾草。前檐、后檐和板下群落中多年生丛生禾草重要值比对照区降低19.52%、67.34% 和92.73%,前檐、后檐和板下多年生杂类草重要值分别为对照区的2.11 倍、9.51 倍和18.95 倍,相比对照区显著增加(P< 0.05),小灌木在后檐群落中重要值显著高于其他区域。总体上,组件遮阴后植物功能群种类增加。

图4 光伏组件对植物群落功能群重要值的影响Figure 4 Effects of the photovoltaic panels on the plant functional groups among different sites
3 讨论
3.1 光伏组件遮阴对植物群落种群结构的影响
环境变化影响着植物群落物种组成及周转,植物群落为适应环境变化在空间分布上具有自适应性[16-18]。相比于木本植物群落,草本植物受光照、土壤等局地微生境异质性影响更为显著[19]。本研究区具有“河谷型萨王纳”特征,土壤贫瘠,保水性差,原生植物群落结构简单[20]。黄茅作为原生群落中优势种,其极强的分蘖能力、较高的株高使之具有数量效应和极强的竞争力[21],而且其致密的地上植被层和地下根系层限制了其他植物拓殖[20],这导致了高度同质化禾草群落的形成。不同于原生生境,光伏电站人工—自然复合生态系统对场区资源的再分配可增加生境破碎度[22],组件倾斜布设使得组件下方不同位置植物群落获得的光、热、水等资源差异明显[8-11],进而导致植物群落结构发生改变。
研究发现,光伏组件遮阴后,群落均匀度、物种多样性显著增加,植株密度显著降低,黄茅优势种地位被替代。随微生境之间遮阴程度差异增大,物种组成相似性呈递减格局,这反映了光伏组件遮阴程度会改变物种周转[23-24],在一定程度上支持“空间自相关性”定律[25]。对内蒙古半干旱区羊草草原的研究[11]发现,板下羊草高度、盖度、密度等显著提升,群落物种多样性降低。这种差异可能来源于半干旱区光伏组件遮阴为羊草提供更好的水热条件的同时,减弱了羊草的“午休”现象,增加了羊草有性繁殖投资,并增加了植物产量[26]。但是本研究区生长季降水丰富,草本植物较短生活史周期使其在时间维度上具有较高的机动性[24],喜热、喜阳的黄茅不适应光伏组件持续遮阴的环境[27],机会主义生活史策略有利于分枝能力强、种子繁殖率高的白苞猩猩草[28]、耐阴耐瘠薄的狗肝菜等植物占领遮阴生境斑块。不同物种株高、冠幅以及光资源生态位等差异导致植物群落立体结构逐渐复杂,有利于更多植物种稳定共存[16,20],从而提高光伏板下物种多样性。
3.2 光伏组件遮阴对植物群落功能群的影响
植物功能群作为响应环境变化的基本单元,与植物的环境适应性、群落构建机制、群落演替方向等密切相关[29]。当生境条件发生改变,具有不同觅养行为和可塑性的植物功能群可在异质生境下形成与之相适应的植物功能群组合[30-31]。分析发现,从对照区、前檐、后檐到板下,组件遮阴程度增加,植物功能群组成在整体上呈现杂类草替代禾草的变化趋势。板间和前檐的较丰富的光资源条件下,多年生丛生禾草依赖其较强的形态可塑性、克隆整合以及觅养行为等占据接近全部的生态空间[32-33],抑制其他功能群植物,成为群落中的优势类群[34]。而在光伏组件的阻风、遮阴、增湿的作用下,多年生丛生禾草从群落中被淘汰。具有较强的耐阴性的杂类草得以生长,在荫蔽环境中占领优势地位。
功能群结构的改变暗示原生群落的生态功能改变[35]。黄茅作为我国西南干热河谷优势禾草类,是光伏板间多年生丛生禾草的关键组分,幼嫩时为优质牧草,但是生长末期的黄茅适口性差,而且该时期其坚硬的茎叶易刺伤牲畜皮肤,对牲畜构成潜在的威胁[36]。光伏组件遮阴后,多年生丛生禾草的优势地位消失,一年生杂类草、多年生杂类草占据群落主要地位,而其组分主要包括白苞猩猩草、假杜鹃、狗肝菜等具有药用价值的多年生植物[37-39]。因此,光伏组件的建设改变了研究区原生草地的牧草生产功能,增加了草地对具有突出药用价值植物类群的产出,但是白苞猩猩草是否对场区植物群落结构和功能产生威胁应进一步深入研究。
4 结论
本研究阐明了云南喀斯特区光伏电站建设后草地植物群落结构空间分异格局,发现光伏组件遮阴可降低群落植株密度、优势度指数,提高多样性指数,黄茅的优势种地位逐渐被白苞猩猩草取代;遮阴程度差异增大,群落物种相似性降低。遮阴后植物功能群种类增加,多年生杂类草替代多年生丛生禾草成为群落中优势功能群;群落由黄茅牧草主导的群落演变为牧草、药用植物共存的群落。总之,光伏电站建设运行后,场区物种多样性增加,群落结构复杂化。
