单氰胺打破葡萄冬芽休眠的生理响应*
2024-01-09李小琴张永福
刘 朝,李小琴,陈 姣,王 凯,张永福
(昆明学院 农学与生命科学学院,云南 昆明 650214)
我国作为葡萄属(VitisL.)植物的起源地之一,境内葡萄属植物种类占全世界的60%左右,是世界上葡萄属植物种类最丰富的国家之一[1].葡萄(VitisviniferaL.)作为世界五大果树之一,更是我国重要的经济果树植物.自2012年,中国葡萄总产量就位居世界首位,在2018年约占全世界葡萄总产量的15.04%[2].目前,除澳门外,葡萄在我国各地均有种植,其中最具代表的是新疆、河北、山东、陕西、云南等地[2].葡萄作为一种冬季落叶果树,冬季休眠是其生长过程中的自然现象,在休眠期间低温积累量是影响葡萄冬芽萌发,从休眠状态进行到生长阶段的重要因素.随着葡萄栽培面积的扩大,南方地区已成为重要产区,但由于南方地区葡萄易在冬季低温积累量不足,使其冬芽不能完成自然休眠,从而导致休眠期延长、萌芽不整齐也不完全、葡萄营养生长过盛、葡萄晚熟、产量下降等问题[3,4].利用化学试剂处理的方法打破葡萄冬芽休眠,提早萌芽,并保持萌芽一致性,进而提高葡萄的产量及品质,已经成为各地葡萄栽培过程中的重要环节.
目前,硝酸铵(NH4NO3)、二氯乙醇(C2H5ClO)、单氰胺(H2CN2)、石灰氮(CaCN2)等破眠剂被广泛应用于葡萄冬芽破除休眠,其中以单氰胺效果最好且无残留[5-8],并且单氰胺处理葡萄冬芽后对果实的还原性糖、可滴定酸、可溶性固形物及维生素C含量[8-11]影响不显著.单氰胺可引起植物的过氧化物酶(POD)活性上升,进而可一定程度上提高POD和氧自由基等引起的氧化过程,使植物呼吸作用向戊糖磷酸途径转化,提高组织新陈代新的活力,打破植物休眠[8].并且,在葡萄冬芽打破休眠的过程中,淀粉、可溶性糖和蛋白等含量、POD和超氧化物歧化酶(SOD)活性均会发生增加或降低的变化[3],从而打破葡萄冬芽休眠,提前冬芽萌芽,提高萌芽率,并最终使葡萄果实提前1~3周成熟[12],提高经济效益[5].
‘水晶’葡萄(Vitisvinifera×V.labrusca‘Shuijing’)为欧美杂交种,目前在云南、山东、贵州等地被广泛种植[13,14],但目前并未见关于单氰胺打破‘水晶’葡萄冬芽休眠的生理机制相关研究报道.因此本试验以‘水晶’葡萄为材料,探讨单氰胺打破葡萄冬芽休眠的生理响应,旨在为葡萄产业化栽培过程中打破葡萄冬芽休眠技术提供理论和技术依据.
1 材料与方法
1.1 试验材料
本研究的‘水晶’葡萄枝条采集于云南省红河州弥勒市东风韵葡萄种植基地,为2021年12月中旬冬剪时,收集修剪下来的一年生木质化枝条,单氰胺(纯度为95%)购自安徽优雅化工有限公司.
1.2 试验方法
1.2.1 试验设计
挑选生长状况一致的葡萄枝条,顶端剪成平口,而下端剪成斜口,每个枝条留3个冬芽.将剪好的葡萄枝条分为10组,每组10枝(共约30个冬芽),其中5组用质量分数2.5%单氰胺溶液均匀涂抹芽和枝条,另外5组涂抹蒸馏水作为对照,然后将枝条扦插到蒸馏水中,放入恒温箱在 25 ℃ 环境下光照 16 h,22 ℃ 环境下黑暗 8 h 进行培养.扦插后每隔 7 d 取样一次,共取6次(第0,7,14,21,28,35 d),进行生理生化指标的测定,同时统计葡萄枝条的发芽率和平均芽长度.
1.2.2 指标测定
取葡萄枝条冬芽采用氮蓝四唑光氧化法、愈创木酚-过氧化氢显色法分别进行SOD[15]和POD活性[16]的测定;采用考马斯亮蓝G-250染色、茚三酮显色法、蒽酮比色法和苯酚-硫酸法分别进行蛋白质含量[17]、游离氨基酸[18]、可溶性糖[19]和淀粉[20]含量的测定;采用纳氏定氮法、钼蓝显色法和四苯硼钠法分别进行氮、磷、钾[21]含量的测定.
总碳含量和碳氮比计算方法如下:
总碳含量=可溶性糖含量×0.42+淀粉含量×0.44;碳氮比(C·N-1)=碳含量/氮含量.
1.3 数据统计与分析
试验数据使用Excel 2007与SPSS 20软件进行统计分析.
2 结果与分析
2.1 葡萄枝条发芽情况观察和统计
据观察在第 0 d 全部葡萄枝条冬芽均处于休眠状态,无萌芽;第 35 d,单氰胺溶液处理(以下简称:试验组)的葡萄枝条与对照组相比,冬芽发芽情况和芽长势更好,且差异显著(图1).试验组葡萄的冬芽在第 7 d 开始萌芽,而对照组在第 21 d 才开始萌芽,明显晚于试验组 14 d.试验组葡萄第 35 d 的萌芽率达到93.6%,为对照组(萌芽率:43.6%)的2.1倍(图2).此外,试验组葡萄第 35 d 的冬芽平均芽长度为 6.8 cm,为对照组(冬芽平均芽长度:3.5 cm)的1.9倍(图3).上述结果说明单氰胺确实可以促进葡萄冬芽萌芽、提高萌芽率并促进冬芽生长.

蒸馏水 单氰胺 蒸馏水 单氰胺 (a)第0 d (b)第35 d图1 蒸馏水和2.5%单氰胺溶液处理后葡萄的发芽情况对比

图2 蒸馏水和2.5%单氰胺溶液处理 葡萄的发芽率对比注:同一处理,不同时段数据上不同小写字母表示差异 显著(P<0.05);不同处理,同一时段数据上不同 大写字母表示差异显著(P<0.05),下同.

图3 蒸馏水和2.5%单氰胺溶液处理葡萄的 冬芽平均芽长度对比
2.2 单氰胺处理对葡萄冬芽SOD和POD活性影响
对照组和试验组葡萄冬芽的SOD活性均随时间呈上升趋势且变化显著.与第 0 d 相比,第 35 d 试验组‘水晶’葡萄冬芽SOD活性增强了51.96%,而对照组SOD活性增强了33.42%.而在整个时间段,试验组的‘水晶’葡萄冬芽SOD活性始终高于对照组,第 35 d 试验组葡萄冬芽SOD活性比对照组高出13.70%(图4).两组葡萄的POD活性也均随时间总体呈上升趋势且变化显著,与第 0 d 相比,在第 35 d 试验组葡萄冬芽的POD活性增强了 1 904.76%,而对照组葡萄的POD活性增强了733.33%.同样的,在整个时间段中,试验组葡萄冬芽的POD活性始终高于对照组,第 35 d 试验组葡萄冬芽的POD活性比对照组高出140.57%(图5).上述结果说明单氰胺溶液处理可以提高葡萄冬芽机体内的SOD和POD活性,增加葡萄冬芽的抗氧化能力,有利于冬芽的生长萌发.

图4 蒸馏水和2.5%单氰胺溶液处理葡 萄冬芽的SOD活性对比

图5 蒸馏水和2.5%单氰胺溶液处理葡 萄冬芽的POD活性对比
2.3 单氰胺处理后葡萄冬芽氮、磷、钾含量变化
试验组葡萄冬芽的氮含量先下降后上升,且差异显著.在第 21 d,其氮含量达到最低值,随后不断上升.而对照组的氮含量则呈现逐渐下降趋势.在第21,28,35 d,试验组葡萄冬芽的氮含量较对照组分别增加了17.47%,85.52%和212.41%,说明单氰胺溶液处理可促使葡萄枝条中氮向冬芽转移,促进冬芽萌发生长(图6).试验组葡萄冬芽的磷含量随时间均呈下降趋势.在整个时间段中,试验组葡萄冬芽的磷含量均明显高于对照组,且差异显著.在第7,14,21,28,35 d,其较对照组分别高了37.70%,26.37%,29.43%,24.95%和21.58%,说明单氰胺处理可减缓葡萄冬芽中磷含量的降低(图7).试验组和对照组葡萄冬芽的钾含量随时间的变化也均呈下降趋势.在整个时间段中,试验组葡萄冬芽的钾含量均明显高于对照组,且差异显著,在第 35 d,其高于对照组41.11%(图8).综上所述,单氰胺溶液处理‘水晶’葡萄冬芽的氮、磷、钾含量均高于对照组,说明单氰胺溶液处理可以提高葡萄冬芽中氮、磷、钾含量,有利于葡萄冬芽的生长.
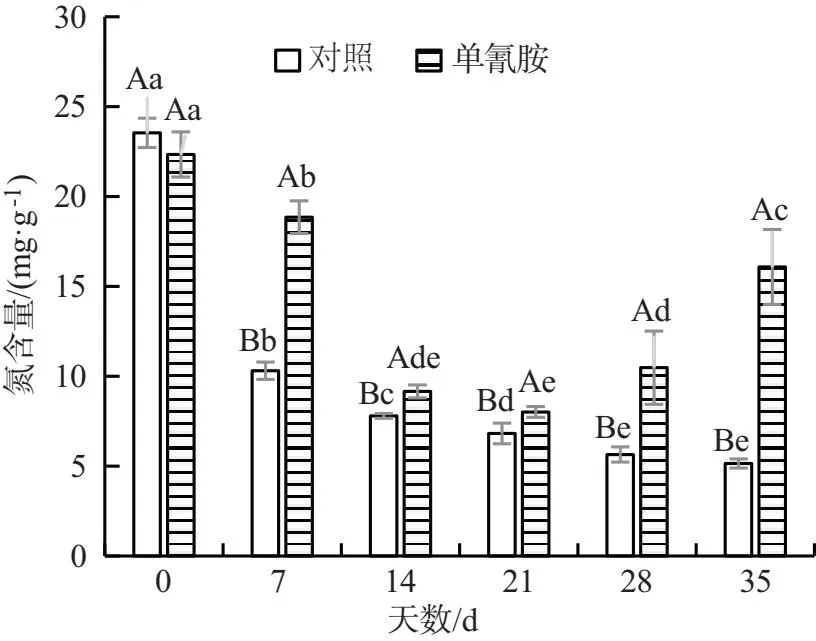
图6 蒸馏水和2.5%单氰胺溶液处理 葡萄冬芽的氮含量对比

图7 蒸馏水和2.5%单氰胺溶液处理 葡萄冬芽的磷含量对比

图8 蒸馏水和2.5%单氰胺溶液处理葡萄 冬芽的钾含量对比
2.4 单氰胺处理对葡萄冬芽淀粉、可溶性糖、总碳含量和碳氮比的影响
试验组和对照组中葡萄冬芽的淀粉含量均随时间呈下降趋势,在第14,21,28,35 d 分别降低了6.80%,23.75%,46.39%和19.93%(图9);两组中葡萄冬芽的可溶性糖含量均随时间上下波动,在第7,14,21,35 d,试验组葡萄冬芽的可溶性糖含量分别较对照组高11.07%,32.16%,14.20%和97.64%(图10);试验组葡萄冬芽的总碳含量在第21,28 d 较对照组分别低了14.20%和41.07%,在第7,35 d 较对照组分别高了20.89%、16.34%,而在第 14 d 与对照差异不显著(图11);试验组葡萄冬芽的碳氮比则先上升后下降,在第7,14,21,28,35 d 较对照分别降低了34.55%,12.57%,27.83%,65.58%和61.45%,且差异显著(图12).综上所述,单氰胺溶液处理和对照组的葡萄冬芽淀粉和总碳含量均逐渐下降;可溶性糖含量和碳氮比均呈现上下波动趋势.在整个时间段中,单氰胺处理的葡萄冬芽中可溶性糖含量总体高于对照组,而淀粉含量和碳氮比均低于对照组.上述结果说明,单氰胺能促进葡萄冬芽淀粉水解,增加可溶性糖含量并提高其氮含量,降低碳含量,为葡萄冬芽的生长提供能量.
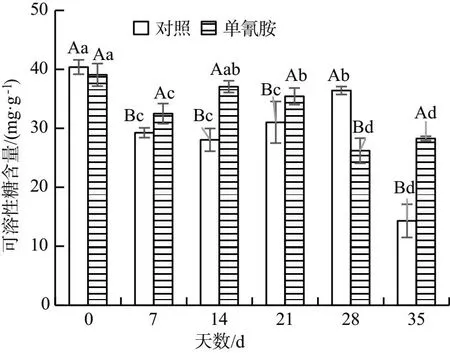
图10 蒸馏水和2.5%单氰胺溶液处理 葡萄冬芽的可溶性糖含量对比
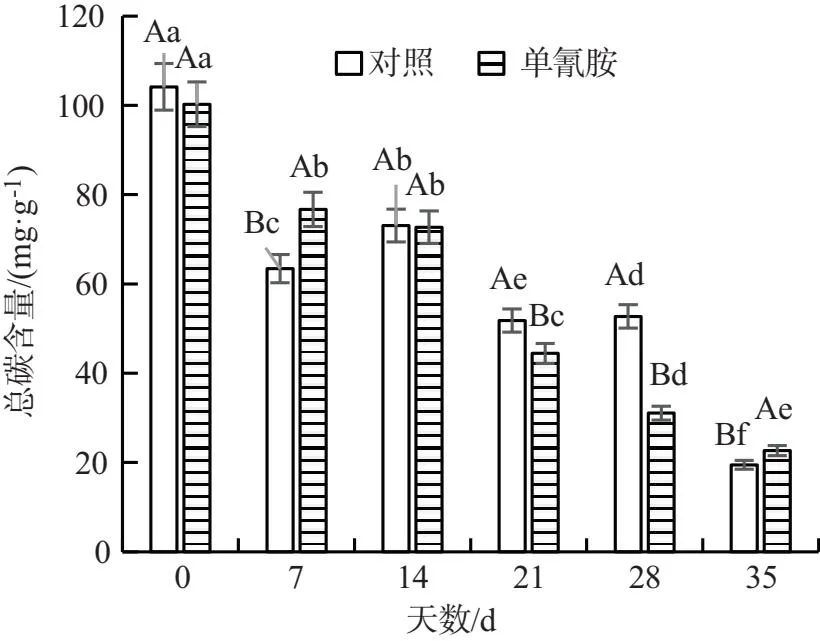
图11 蒸馏水和2.5%单氰胺溶液处理 葡萄冬芽的总碳含量对比

图12 蒸馏水和2.5%单氰胺溶液处理 葡萄冬芽的碳氮比对比
2.5 单氰胺处理对葡萄冬芽蛋白质和游离氨基酸含量的影响
试验组和对照组葡萄冬芽的蛋白质含量随时间总体呈上升趋势,且在整个时间段中,试验组葡萄冬芽的蛋白质含量均明显高于对照组,且差异显著.在第7,14,21,28,35 d,试验组较对照组分别高了41.91%,40.51%,40.28%,90.23%和98.68%(图13).而两组葡萄冬芽中游离氨基酸含量随时间均呈上下波动的趋势,且在整个时间段中,试验组葡萄冬芽的游离氨基酸含量总体比对照组高,在第14,21,28,35 d,分别高了161.92%,154.80%,47.52%和93.89%(图14).综上而言,单氰胺可促进葡萄冬芽蛋白质的合成,增加游离氨基酸含量,促进冬芽生长.

图13 蒸馏水和2.5%单氰胺溶液处理 葡萄冬芽的蛋白质含量对比

图14 蒸馏水和2.5%单氰胺溶液处理 葡萄冬芽的游离氨基酸含量对比
3 讨论
单氰胺作为一种植物生长调节剂,可以促进果树萌芽、冬芽生长,目前已在蓝莓、桃、葡萄等多种果树上成功应用[22].李鹏飞等[23]利用单氰胺处理‘霞多丽’葡萄枝条,结果显示单氰胺可打破葡萄休眠,提高葡萄发芽率和萌芽整齐度,与本实验结果较为一致.黄晓婧等[3]利用单氰胺处理‘夏黑’葡萄枝条,发现‘夏黑’葡萄的POD和SOD均显著上升,并且处理组均明显高于对照组,也与本试验结果较为一致.张昂等[24]利用单氰胺对‘霞多丽’和‘赤霞珠’葡萄进行处理,但其SOD活性变化为先下降后上升,与本试验结果不一致.这主要是因为葡萄萌芽时间存在差异.而当‘霞多丽’和‘赤霞珠’葡萄萌芽后,其SOD活性也为上升趋势,并且其单氰胺处理组明显高于对照组,与本试验结果也较为一致.这也佐证了SOD活性是葡萄打破休眠,进入萌芽的重要指标.然而,‘霞多丽’和‘赤霞珠’葡萄萌芽后的POD活性呈下降趋势,与本试验结果不一致.因此在打破葡萄休眠,进入萌芽这一过程中,引起POD活性变化的机制还需要进行进一步的探讨.
黄晓婧等[3]利用单氰胺处理‘夏黑’葡萄后,其可溶性糖先升高后降低,并且在萌芽前,单氰胺处理后的可溶性糖含量高于对照组,而在萌芽后却低于对照组.单氰胺处理‘夏黑’葡萄后,其淀粉含量下降,且对照组高于单氰胺处理组.这些结果与本试验结果较为一致.谭一婷等[25]研究表明单氰胺处理后的不同葡萄品种中的可溶性糖和淀粉含量均高于对照,与本研究结果不一致.上述研究结果表明,在葡萄打破休眠进入萌芽过程中,可溶性糖和淀粉含量的变化在不同葡萄品种中并没有一致的变化规律,其原因还有待进一步的探讨.
氮、磷、钾对植物的生长发育至关重要,并且氮、磷、钾可从植株的老叶或枝条转移至新叶或新芽,促进新叶或新芽的生长.本试验发现,单氰胺处理‘水晶’葡萄冬芽的氮、磷、钾、蛋白质和游离氨基酸含量明显高于对照组,说明氮、磷、钾、蛋白质和游离氨基酸含量也在葡萄打破休眠后萌芽过程中具有重要作用.总体而言,单氰胺处理可增加‘水晶’葡萄淀粉水解,提高SOD和POD活性,促进葡萄枝条中氮、磷、钾转移至冬芽,提高葡萄冬芽中蛋白质合成,使葡萄冬芽提前萌发并生长旺盛.
4 结论
本研究通过对单氰胺处理后‘水晶’葡萄冬芽的萌发情况观察及其生理指标测定,发现:在处理后第 35 d,与对照组相比,试验组葡萄的萌芽时间提前了 14 d,冬芽萌芽率和平均芽长度分别是对照组的2.1,1.9倍;试验组葡萄冬芽的SOD、POD活性较对照组分别增加了13.70%和140.57%,其氮、磷、钾、可溶性糖、蛋白质、游离氨基酸含量较对照组分别增加了212.41%,21.58%,41.11%,97.64%,98.68%,93.89%;而其淀粉、碳氮比较对照组分别降低了19.93%和61.15%.由此可见,POD活性、氮、可溶性糖、蛋白质、游离氨基酸含量是单氰胺解除葡萄冬芽休眠的主要生理因子.综上所述,2.5%单氰胺处理‘水晶’葡萄后,可引起葡萄冬芽内生理变化,缩短葡萄冬芽萌芽周期,提高萌芽率和萌芽整齐度,促进冬芽的生长.
