洋葱黄色条纹突变体叶绿体基因组测序及嵌合体验证
2023-12-02惠林冲陈微张仕林李威亚何林玉杨海峰潘美红
惠林冲 陈微 张仕林 李威亚 何林玉 杨海峰 潘美红


摘要:为鉴定洋葱黄条纹突变体中叶绿体基因组基因突变,利用洋葱黄色条纹突变体进行黄色组织叶绿体基因组测序及差异位点验证分析,对突变体种子果夹表型特征与出苗后表型进行对比分析,并利用ABP1-1、KMOX-1和MYB-2分子标记对4份黄条纹黄化程度在50%以上的植株绿色和黄色组织DNA进行分子标记多态性检测。结果表明,测序获得突变体叶绿体基因组153 585 bp,与洋葱叶绿体参考基因组比对筛选出5个差异位点,PCR扩增及测序分析发现叶绿体基因组碱基的突变、插入并不导致黄条纹突变;突变果夹黄化程度与母株上种子出苗后黄化程度成正比,果夹越黄,种子出苗叶片越黄,3个分子标记对同一株黄色和绿色组织进行PCR验证,发现绿色与黄色组织扩增条带不一致。综上所述,洋葱叶片黄色条纹突变是一种嵌合体突变。
关键词:洋葱;黄条纹;叶绿体;嵌合体;基因组测序
中图分类号:S633.201文献标志码:A
文章编号:1002-1302(2023)20-0043-06
绿色植物叶片容易突变成黄色条纹状,可以在园艺花卉中得到应用。不同植物的突变机制各不相同,有核基因突变[1-2]、核质基因共同作用[3]及母系遗传突变类型[4-5]。洋葱黄条纹突变属于母系遗传,Tatebe研究表明,洋葱黄色条纹正反杂交遗传分析属于母系遗传[6];杨海峰等研究发现,研究此突变材料自交无固定分离比例,且黄色条纹不退化,超微结构观察到类囊体膜结构受损[7],以上均是从生理形态特征进行研究,未从分子角度研究黄色条纹突变机制。祝利霞用2个群体对甘蓝型油菜黄化突变体基因精细定位,结果表明,Bnac.HO1基因在突变体中缺失了C基因组上的同源拷贝[8]。芥菜型油菜黄化突变体L638-y叶片叶绿素合成代谢受阻,受阻位点在CoprogenⅢ到ProtoⅨ的反应步骤,该突变体可能是一种新类型的缺绿体突变体[9]。孙小秋等通过甲基磺酸乙脂(EMS)诱变获得1对隐性核基因控制遗传稳定的水稻黄绿叶突变体ygl98,基因组序列分析发现突变体在编码镁离子螯合酶ChlD 亚基的OsChlD基因编码区第1 522碱基处,碱基G突变为碱基A,造成编码蛋白序列丙氨酸突变成苏氨酸[10]。Li等对银杏叶片黄条纹突变体转录组测序分析及实时定量聚合酶反应(RT-qPCR)分析发现PPO和NYC/NOL基因表达影响叶绿素的生物合成,并促进叶绿素b降解为叶绿素a,而上调的基因Z-ISO、ZDS和LCYE增强了类胡萝卜素的积累。因此,類胡萝卜素与叶绿素比例的变化是形成银杏叶片黄色条纹突变的主要因素,并未定位到基因突变位点[11]。
洋葱黄色条纹突变具有母系遗传特征,而叶绿体基因为母系遗传,相对核基因组保守性较高[12]。洋葱叶绿体基因组已公布,包括正常可育和2种细胞质雄性不育系(CMS-S和CMS-T)[13],为了鉴定洋葱黄条纹突变体是否是叶绿体基因组基因发生突变,拟对叶绿体基因组重测序,对差异位点进行PCR验证,在前期已对洋葱黄条纹突变体形态特征、超显微结构等进行研究的基础上,利用分子标记对同一株洋葱黄色和绿色组织进行DNA扩增,开展叶绿体突变基因研究,以期为研究洋葱叶绿体光合机制及遗传发育奠定基础。
1 材料与方法
1.1 材料
洋葱黄条纹突变体收集于江苏省连云港市农业科学院东辛试验基地,为中日照类型洋葱,根据黄化程度不同选取不同类型洋葱,HT01~HT04、HT06~H19共18个黄条纹突变材料(因HT05苗期黄化程度100%,苗木未能长到取样时期),N20、N21为正常洋葱植株,HT13黄色组织用于叶绿体基因组测序(图1),材料特征见表1。
1.2 试验方法
1.2.1 洋葱叶绿体基因组测序及差异序列验证
采用洋葱HT13突变材料全黄组织,提取DNA后送上海元莘生物医药科技有限公司进行测序,超声波将DNA打断进行纯化构建测序文库,质检合格后采用Illumina HiSeqTM平台进行测序,以NCBI数据库中洋葱叶绿体KM088013.1为参考基因组,进行突变体叶绿体基因组de novo拼接和遗传变异检测。将基因组比对后,对差异位点设计6对引物(表2),送苏州金唯智生物科技有限公司合成。提取18份黄条纹突变材料黄色组织和2份正常洋葱DNA进行PCR扩增,PCR扩增试剂购于宝日医生物技术(北京)有限公司,反应体系为50 μL,包括4 μL DNA,5 μL 10×PCR 缓冲液,引物1和引物2分别 3 μL,4 μL dNTPs(10 mmol/L),0.5 μL TaqM和30.5 μL ddH2O,PCR扩增条件:94 ℃预变性 4 min;94 ℃变性30 s,56 ℃退火30 s,72 ℃延伸 1 min,30个循环;72 ℃延伸10 min[14],PCR扩增产物用1%琼脂糖凝胶电泳检测,胶回收后PCR产物送苏州金唯智生物科技有限公司测序,拼接结果使用DNAman软件进行序列比对分析。
1.2.2 洋葱黄条纹嵌合体突变验证
将洋葱黄条纹突变体开花期单株自交,种子受精灌浆后(未开裂),对种子外壳出现的黄色突变进行标记,收获的种子进行分类(全黄株、黄绿相间株、全绿株),秋季9月中旬播种和第2年4月对植株进行表型统计,并取4株(HT01、HT11、HT13和HT19)黄条纹黄化程度在50%以上正常生长的洋葱球叶片,分别提取黄色组织和绿色组织的DNA,利用笔者所在实验室前期筛选出的3个CDDP多态性丰富的分子标记ABP1-1、KMOX-1和MYB-2[15](相关数据未发表)。引物见表2,PCR扩增条件同上述,PCR产物采用8%聚丙烯酰胺凝胶电泳检测,电压为100 V,电泳2.5 h,硝酸银染色,采用相机在胶片灯上拍照记录。
2 结果与分析
2.1 洋葱黄色条纹突变体叶绿体基因组测序分析
以洋葱(Allium cepa L.)的叶绿体基因组序列(https://www.ncbi.nlm.nih.gov/nuccore/744671124/)作为参考,该参考基因组大小为153 529 bp,测序获得洋葱黄条纹突变体黄色组织叶绿体基因组大小153 585 bp,比已公布的正常可育洋葱多56 bp(图2)。数据基于GATK校正比对结果,利用GATK软件进行SNP、small indel检测,并过滤掉测序深度和比对质量值较低的位点,得到高可信度的20个SNP和5个small indel数据集,采用Annovar程序结合洋葱叶绿体基因组的gff3基因注释信息对得到的SNP和small indel进行注释(表3)。
利用DNAman软件对已公布的洋葱可育株N(GenBank数据中登录号:KM088013,下同)、CMS-S(KM088014)、CMS-T(KM088015)、可育株N(KF728080.1)和CMS-S(KF728079.1)进行比对,排除洋葱不育系CMS-S和CMS-T特有的突变、插入和缺失序列外,筛选出5个差异位点,包括碱基突变、插入和缺失(表4)。
2.2 洋葱黄条纹突变体叶绿体基因组差异位点PCR验证
对18份突变材料黄色组织和2份正常洋葱为对照的DNA进行PCR验证,其中包括测序用的材料HT13,因扩增条带差别较小 在琼脂糖凝胶电泳图上不能完全区分条带的差异。尽管引物3扩增中有54 bp碱基的插入,但不能被清楚地区分开(图3)。对所有的PCR产物进行胶回收测序,DNAman软件进行比对,HT13材料叶绿体DNA全长测序中差异位点与PCR验证完全一致,说明黄条纹突变体基因组序列可靠。引物2中GAA突变TTC,检测发现只有HT13、N20和N21这3份材料为TTC,其他17份突变材料均为GAA;引物3扩增中除了HT04、HT06、HT08、HT14外,其他均有插入TTCTATTCTATATGTACATCTACTATATACATATTGTAAT TTTATCTAAATTTT片段,且2份绿色正常组织N20和N21均有插入片段(图4);同样引物1、4、5中的插入或突变在黄条纹突变体和正常植株中均有出现,与黄条纹突变和黄化程度均无关系,洋葱黄条纹突变体与叶绿体基因组序列碱基突变无关系。
2.3 洋葱黄条纹突变体嵌合体验证
对洋葱黄条纹突变体的种子果夹进行观察分类,分为全黄、半黄和全绿色3种类型,因种子外表皮是黑色,在未成熟果夹上做好标记,分类收集种子(图5)。播种后观察苗期的表型:无固定分离比,外壳为全黄色,对应种子出苗后也是全黄株;外壳绿色对应种子苗期和生长期均为全绿色植株;黄绿相间的在苗期及生长期同样表现出黄绿色相間条纹状,黄化程度与母株上种子出苗后黄化程度成正比,具有嵌合体母系遗传特征。
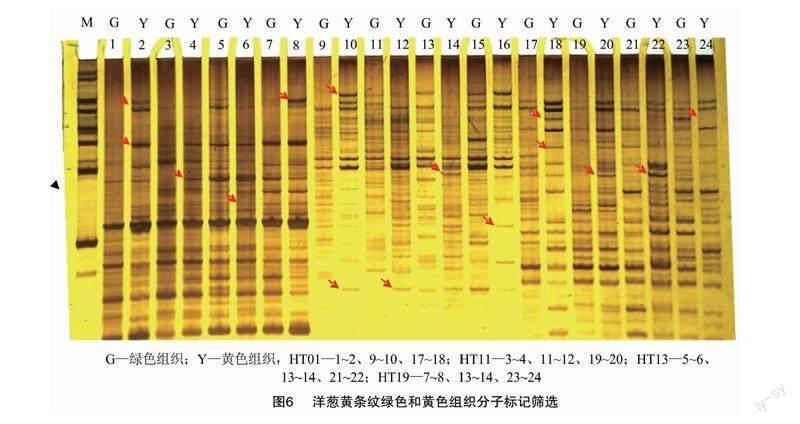
对4株(HT01、HT11、HT13和HT19)黄条纹黄化程度在50%以上的植株绿色和黄色组织DNA进行分子标记多态性检测,3个标记(ABP1-1、KMOX-1和MYB-2)结果显示同一株绿色部分和黄色部分多态性存在差异,3个标记均显示4株黄绿2个部分DNA存在差异(图6),表明洋葱黄条纹突变体是一种嵌合体突变类型。
3 讨论与结论
洋葱黄色条纹突变属于母系遗传,研究发现黄色组织中仍然具有叶绿素,但含量减少,叶绿素合成受阻导致黄色条纹,叶色变异主要由核基因控制,也可由叶绿体基因或核质互作控制[16],叶绿体中含有大量的蛋白酶,以促进蛋白稳态和发育,拟南芥中EVR3编码叶绿体金属蛋白酶调控叶片斑纹的形成[17]。白茶新叶发现黄叶突变体,对黄绿2个部分转录组测序分析发现,黄色叶片着色主要受色素代谢的影响,包括叶绿素、类胡萝卜素和类黄酮的色素代谢[18],百合黄色幼苗致死突变体lrysl1转录组分析发现叶绿体发育异常且相关的基因下调表达[19]。在叶片黄化突变基因定位研究中,黄瓜幼叶黄色突变体C777精细定位到yyl-1基因上SNP中G变成A,形成终止密码子导致黄叶的出现[20]。而水稻白化转绿突变体albg是Os03g0597200基因从ATG开始第1 387位发生了1个碱基G的插入,导致叶色在苗期出现突变[21]。本研究中研究思路考虑转录组测序和突变基因的定位,可能是洋葱叶绿体基因发生突变引起洋葱黄条纹突变体。对洋葱黄条纹黄色组织进行叶绿体基因组测序,获得153 585 bp,与洋葱已公布的参考基因组KM088013进行比对,尽管分析发现有20个SNP和5个插入或缺失,但与洋葱其他几个不育系类型进行DNAman软件序列分析,在不育系中有同样在黄条纹中出现的碱基,并不是黄条纹叶绿体基因所特有的,筛选出5个差异位点。设计引物PCR验证,差异位点在黄条纹中和正常洋葱中均存在突变和插入或缺失,并不是叶绿体基因组碱基的变化导致洋葱出现黄条纹叶片。
田间发现具有典型黄条纹突变体的洋葱单株人工自交后,果夹也具有黄化表型特征,分为全黄、黄绿相间和全绿色3种类型果夹,但种子无差别均为黑色,并做标记分类播种后,洋葱苗黄条纹的表型特征与母株上的果夹黄色区域表型特征对应。与植物嵌合体有一定的联系,基因突变也能产生嵌合体,典型特征是植物的茎尖分生组织由2种或2种以上遗传型不同的细胞组成,共同发育成完整植株[22]。SSR、SNP/InDel作为检测嵌合体已在植物中应用,研究发现橙子品种Zhiwenzhou是一种嵌合体[23],葡萄开发新品种比较困难,Preiner等利用葡萄分生组织嵌合体2个新品种(Cabernet sauvignon和Babic),并利用SSR标记进行鉴定[24]。Zhou等对2种嵌合体进行随机扩增多态性DNA标记(RAPD)分析发现嵌合体不仅具有2个亲本的特异性条带,还分别产生了各自特有的新条带,说明嵌合体在DNA可能存在着互作[25]。而王彩霞等利用69条ISSR引物对文心兰黄化突变体和正常植株进行遗传变化分析发现,其中67条引物在黄化突变体和正常植株之间未呈现多态性,引物UBC827和UBC897对黄化突变体和正常植株的扩增产物具有差异,分别多扩增出1个约为600 bp和200 bp的条带[26]。本研究利用前期在洋葱中筛选CDDP分子标记多态性丰富的ABP1-1、KMOX-1和MYB-2等3个标记,对洋葱黄条纹突变体的黄色组织和绿色组织进行嵌合体验证,每个标记在4株不同洋葱突变株中均有特异条带,同一株黄绿2个部分DNA有差异,说明洋葱黄条纹突变体是嵌合体突变。
一般嵌合体存在长期保存的问题,尤其不适合低温存储,容易导致芽变,Kulus等利用包囊脱水技术冷冻保存周缘嵌合体菊花防止芽变[27]。而洋葱黄条纹是通过种子进行稳定的母系遗传,能够很好的保存,是研究光合作用机制和形态学标记的重要材料。
参考文献:
[1]Qin D D,Dong J,Xu F C,et al. Characterization and fine mapping of a novel barley stage green-revertible albino gene (HvSGRA) by bulked segregant analysis based on SSR assay and specific length amplified fragment sequencing[J]. BMC Genomics,2015,16(1):1-14.
[2]Ye W J,Hu S K,Wu L W,et al. White stripe leaf 12 (WSL12),encoding a nucleoside diphosphate kinase 2 (OsNDPK2),regulates chloroplast development and abiotic stress response in rice (Oryza sativa L.)[J]. Molecular Breeding,2016,36(5):57-72.
[3]Zhao H B,Guo H J,Zhao L S,et al. Agronomic traits and photosynthetic characteristics of chlorophyll-deficient wheat mutant induced by spaceflight environment[J]. Acta Agronomica Sinica,2011,37(1):119-126.
[4]Gu J,Miles D,Newton K J. Analysis of leaf sectors in the NCS6 mitochondrial mutant of maize[J]. The Plant Cell,1993,5(8):963-971.
[5]Sujatha M,Subrahmanyam N C. Characterisation of nuclear gene controlled yellow stripe mutant of Pennisetum glaucum (L.) R.Br.[J]. Plant Science,1991,73(1):55-64.
[6]Tatebe T. Genetic studies on the leaf variegation of Allium cepa L.[J]. Engei Gakkai Zasshi,1968,37(4):345-348.
[7]楊海峰,惠林冲,潘美红,等. 洋葱黄色条纹突变体的特征特性[J]. 江苏农业学报,2019,35(1):157-165.
[8]祝利霞. 甘蓝型油菜黄化和光叶突变体的基因定位及克隆[D]. 武汉:华中农业大学,2014.
[9]吕 明. 芥菜型油菜黄化突变体L638-y叶片缺绿的生化机制探讨[D]. 杨凌:西北农林科技大学,2010.
[10]孙小秋,王 兵,肖云华,等. 水稻ygl98黄绿叶突变基因的精细定位与遗传分析[J]. 作物学报,2011,37(6):991-997.
[11]Li W X,Yang S B,Lu Z G,et al. Cytological,physiological,and transcriptomic analyses of golden leaf coloration in Ginkgo biloba L.[J]. Horticulture Research,2018,5:12.
[12]李志远,于海龙,方智远,等. 一个芥蓝变种特异的叶绿体InDel标记CP-ID120026[C]//中国园艺学会2017年论文摘要集.昆明:中国园艺学会,2017:109.
[13]Kim S,Park J Y,Yang T.Comparative analysis of the complete chloroplast genome sequences of a normal male-fertile cytoplasm and two different cytoplasms conferring cytoplasmic male sterility in onion (Allium cepa L.)[J]. The Journal of Horticultural Science and Biotechnology,2015,90(4):459-468.
[14]潘美红,杨海峰,惠林冲,等. 洋葱CMS-T型育性分子标记的筛选与鉴定[J]. 西北农业学报,2018,27(4):576-585.
[15]Collard B C Y,MacKill D J.Conserved DNA-derived polymorphism (CDDP):a simple and novel method for generating DNA markers in plants[J]. Plant Molecular Biology Reporter,2009,27(4):558-562.
[16]魏彦林,施勇烽,吴建利. 水稻核基因控制的叶色变异[J]. 核农学报,2011,25(6):1169-1178.
[17]Qi Y F,Wang X M,Lei P,et al. The chloroplast metalloproteases VAR2 and EGY1 act synergistically to regulate chloroplast development in Arabidopsis[J]. Journal of Biological Chemistry,2020,295(4):1036-1046.
[18]Wang P J,Zheng Y C,Guo Y C,et al. Widely targeted metabolomic and transcriptomic analyses of a novel albino tea mutant of “Rougui”[J]. Forests,2020,11(2):229.
[19]Du W K,Hu F R,Yuan S X,et al. The identification of key candidate genes mediating yellow seedling lethality in a Lilium regale mutant[J]. Molecular Biology Reports,2020,47(4):2487-2499.
[20]Hu L L,Zhang H Q,Xie C,et al. A mutation in CsHD encoding a histidine and aspartic acid domain-containing protein leads to yellow young leaf-1 (yyl-1) in cucumber (Cucumis sativus L.)[J]. Plant Science,2020,293:110407.
[21]简 磊,王仲康,曾冬冬,等. 水稻白化转绿突变体albg的鉴定和基因精细定位[J]. 核农学报,2017,31(12):2289-2297.
[22]朱雪云,陈利萍. 植物嵌合体的研究与应用[J]. 核农学报,2010,24(6):1185-1191.
[23]Zhu S P,Wang F S,Shen W X,et al. Genetic diversity of Poncirus and phylogenetic relationships with its relatives revealed by SSR and SNP/InDel markers[J]. Acta Physiologiae Plantarum,2015,37(7):141.
[24]Preiner D,Markovi[KG-*5]c[DD(-1*2/3]' Z,ikuten I,et al. In vitro synthesis of grapevine (Vitis vinifera L.) intraspecific chimeras using meristematic bulk tissue grafting[J]. Scientia Horticulturae,2019,246:965-970.
[25]Zhou J M,Hirata Y,Nou I S,et al. Interactions between different genotypic tissues in citrus graft chimeras[J]. Euphytica,2002,126(3):355-364.
[26]王彩霞,田韋韦,田 敏,等. 文心兰黄化突变体的初步研究[J]. 核农学报,2013,27(12):1845-1852.
[27]Kulus D,Abratowska A,Mikua A. Morphogenetic response of shoot tips to cryopreservation by encapsulation-dehydration in a solid mutant and periclinal chimeras of Chrysanthemum×grandiflorum/Ramat./Kitam.[J]. Acta Physiologiae Plantarum,2018,40(1):18.
收稿日期:2022-11-30
基金项目:江苏省连云港市财政专项(编号:QNJJ2206);连云港市第六期“521工程”科研项目(编号:LYG06521202134) 。
作者简介:惠林冲(1988—),男,江苏灌南人,硕士,助理研究员,从事蔬菜育种、栽培及分子生物学研究。E-mail:huilinchong@yeah.net。
通信作者:潘美红,硕士,副研究员,从事蔬菜育种、栽培及利用研究。E-mail:7991454@163.com。
