不同培育模式下日本落叶松林灌草和土壤养分特征研究
2023-11-28杨圆圆于世河卜鹏图陆爱君王占伟郑颖冯健
杨圆圆 于世河 卜鹏图 陆爱君 王占伟 郑颖 冯健


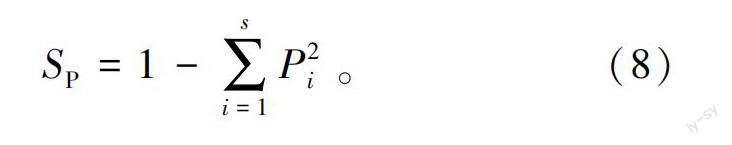
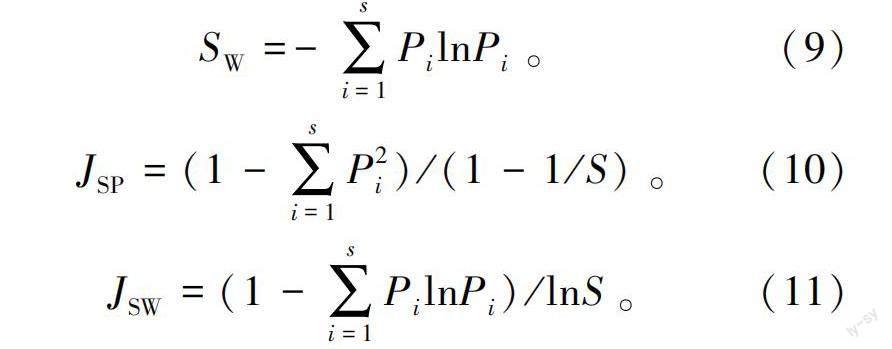
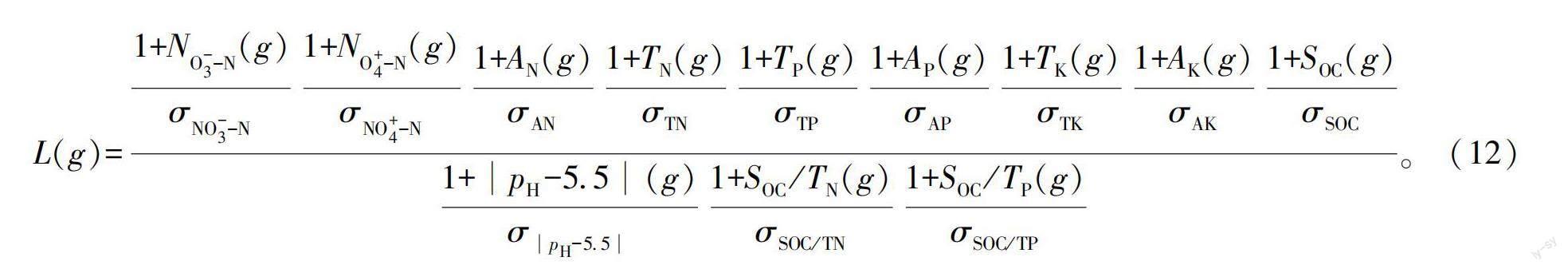









摘 要:探討不同培育模式日本落叶松林灌草、土壤养分特征,以期为人工林的营建和持续健康经营提供参考。以辽东山区现有典型培育模式下的日本落叶松(Larix kaempferi)林,即日本落叶松-白桦(Betula platyphylla)带状混交林(文中记为DZ)、日本落叶松-白桦-日本桤木(Alnus japonica)块状混交林(文中记为KZ)及日本落叶松纯林(CK)为研究对象,调查并分析林下灌草组成、多样性和土壤养分特征及其影响林下灌草分布和多样性的主要环境因子。结果表明,不同培育模式下日本落叶松林灌草组成丰富,灌草总数表现为DZ>KZ>CK;草本层Simpson多样性指数(S)、以Simpson多样性指数为基础的Pielou均匀度指数(J)、Shannon-Wiener多样性指数(S)和以Shannon-Wiener多样性指数为基础的Pielou均匀度指数(J)表现为CK>KZ>DZ,灌木层S、J、J表现为DZ>CK>KZ,但总体上差异不显著。日本落叶松林土壤养分含量和C∶N、C∶P和N∶P有所差异,利用乘除法并结合土壤元素含量判定土壤养分表现为DZ>CK>KZ。影响不同培育模式下落叶松林灌草分布和多样性的主要环境因子不同,土壤硝态氮、铵态氮和速效磷是影响草本分布的主要环境因子,硝态氮和铵态氮是影响灌木分布的主要环境因子,单株材积是影响草本多样性的主要环境因子,而土壤C∶N是影响灌木多样性的主要环境因子。日本落叶松-白桦带状混交培育模式林下灌草数量、灌木多样性,土壤养分条件和木材蓄积优于其他2种培育模式;可结合影响不同培育模式日本落叶松林灌草组成和多样性的主要因子分类进行森林经营,协调灌草多样性、林分因子和土壤养分之间的关系,促进辽东山区日本落叶松林健康可持续经营。
关键词:培育模式;灌草多样性;土壤养分;典范对应分析;冗余分析
中图分类号:S725 文献标识码:A 文章编号:1006-8023(2023)06-0012-14
Study on Shrub-herb and Soil Nutrient Characteristics of Larix kaempferi
Forest under Different Cultivation Modes
YANG Yuanyuan, YU Shihe, BU Pengtu, LU Aijun, WANG Zhanwei, ZHENG Ying, FENG Jian
(1.Liaoning Academy of Forestry Science, Shenyang 110032, China; 2.Liaoning Baishilazi Forest Ecosystem National Research Station, Dandong 118201, China)
Abstract:The characteristics of shrub-herb and soil nutrient in different cultivation modes of Larix kaempferi forest were discussed, so as to provide reference for the establishment and sustainable healthy management of plantation forests. Taking the L. kaempferi forest under the existing typical cultivation modes in the eastern mountainous area of Liaoning Province, such as L. kaempferi-Betula platyphylla strip mixed forest (DZ), L. kaempferi-B. platyphylla-Alnus japonica block mixed forest (KZ) and L. kaempferi pure forest (CK) as the research object, the forest understory shrub-herb composition, diversity, soil nutrient characteristics and the dominant environmental factors affecting the distribution and diversity of understory shrub-herb were investigated and analyzed. The result showed that composition of shrub-herb in L. kaempferi forest under different cultivation modes was rich, and the total number was DZ>KZ>CK. The herb layer Simpson diversity index (S), Pielou community evenness index based on Simpson diversity index (J), Shannon-Wiener diversity index (S) and Pielou community evenness index based on Shannon-Wiener diversity index (J) showed CK>KZ>DZ, and shrub layer SP, J and J showed DZ>CK>KZ, but the overall difference was not significant. The soil nutrient content and C∶N, C∶P and N∶P of L. kaempferi forest were different. The soil nutrient was determined by multiplication division method combined with soil element content as DZ>CK>KZ. The main environmental factors that affected the distribution and diversity of shrub-herb in different cultivation modes of L. kaempferi forest were different. Soil nitrate nitrogen, ammonium nitrogen and available phosphorus were the main environmental factors affecting the distribution of herb, nitrate nitrogen and ammonium nitrogen were the main environmental factors affecting the distribution of shrub, volume was the main environmental factor affecting herb diversity, and soil C:N was the main environmental factor affecting shrub diversity. The number of shrub-herb, shrub diversity, soil nutrient conditions and wood accumulation of L. kaempferi-B. platyphylla strip mixed cultivation mode were better than those of the other two cultivation modes. The forest management can be combined with the classification of dominant factors affecting the composition and diversity of L. kaempferi forest in different cultivation modes to coordinate the relationship between shrub-herb diversity, stand factors and soil nutrients, and to promote the healthy and sustainable management of L. kaempferi forest in the eastern mountainous area of Liaoning Province.
Keywords:Cultivation mode; shrub-herb diversity; soil nutrient; CCA; RDA
收稿日期:2023-03-05
基金项目:国家重点研发计划(2022YFD220030204);辽宁省农业科学院学科建设(2022DD217036)。
第一作者简介:杨圆圆,硕士,助理工程师。研究方向为林木育种及森林培育。E-mail: 1805932544@qq.com
*通信作者:冯健,博士,正高级工程师。研究方向为林木育种。E-mail: lnlkyfj@163.com
引文格式:杨圆圆,于世河,卜鹏图,等.不同培育模式下日本落叶松林灌草和土壤养分特征研究[J].森林工程, 2023, 39(6):12-25.
YANG Y Y, YU S H, BU P T, et al. Study on shrub-grass and soil nutrient characteristics of Larix kaempferi forest under different cultivation modes[J]. Forest Engineering, 2023, 39(6):12-25.
0 引言
落叶松是中国东北地区主要造林树种,现有林分占人工林的70%以上,长期以纯林为主的培育模式导致落叶松林生产力下降、生态系统稳定性和服务功能被削弱,为此,该地区在长期造林实践过程中通过引入白桦(Betula platyphylla)、日本桤木(Alnus japonica)、水曲柳(Fraxinus mandshurica)和核桃楸(Juglans mandshurica)等阔叶树种,形成了以不同树种搭配和不同混交方式相结合的落叶松培育模式;在东北地区重要生态区辽东山区,存在3种典型的日本落叶松培育模式,即日本落叶松(Larix kaempferi)-白桦带状混交、日本落叶松-白桦-日本桤木块状混交和纯林培育模式。
不同培育模式的林分往往具有独特的林分结构、林下物种多样性和土壤养分特征,一方面,这些特征因混交方式、混交树种、混交比例不同而产生各异格局,如油松(Pinus tabuliformis)-元宝枫(Acer truncatum)块状混交培育模式土壤养分及涵养水源功能优于带状混交与油松纯林;华北落叶松(Larix gmelinii var. principis-rupprechtii)-白桦混交林土壤肥力质量好于华北落叶松纯林和华北落叶松-樟子松(Pinus sylvestris var. mongolica)混交林;落叶松(Larix gmelinii)-水曲柳混交林在林分生长、植物多样性和生物量分配比例上优于落叶松纯林,但土壤硝态氮、全磷和有效磷含量却低于纯林。另一方面,林分因子、物种多样性和土壤养分之间互相影响,在人工林物种多样性逐渐受到重视的大背景下,探究林下植被生长、多样性格局和主导因素成为研究熱点,如木荷(Schimasuperba)次生林土壤全钾、有机质含量和含水率是影响物种多样性的主要环境因子,而土壤容重、全磷和含水量则显著影响阔叶红松林物种多样性。目前相关研究关注了东北地区落叶松林的林分状况、植被特征和土壤条件等,但对于特定培育模式下落叶松林灌草和土壤特征的相关研究较少,特别是块状混交林的相关研究鲜见报道。因此,本研究从辽东山区现有3种典型的日本落叶松培育模式入手,分析其林下灌草组成、多样性和土壤养分条件差异及其关联,旨在评价不同培育模式下灌草和土壤养分优劣,探究影响林下灌草多样性的主导因子,为研究区域内日本落叶松林的营建和持续健康经营提供参考。
1 材料与方法
1.1 研究区概况
研究区位于辽宁省清原满族自治县(124°20′~125°29′E,41°48′~42°29′N),地处辽宁东部山区。气候类型为中温带大陆性季风气候,年均气温为5 ℃,年均降水量为700~800 mm,主要集中在4—9月,无霜期120 d。林地土壤多为山地棕色森林土。该地区植被覆盖率高,森林资源丰富,森林覆盖率达77.1%,森林总蓄积达2 056万m。该区植被属于长白植物区系,以人工林为主,主要包括日本落叶松、红松(Pinus koraiensis)和长白落叶松(Larix olgensis)。
1.2 样地设置与样品采集
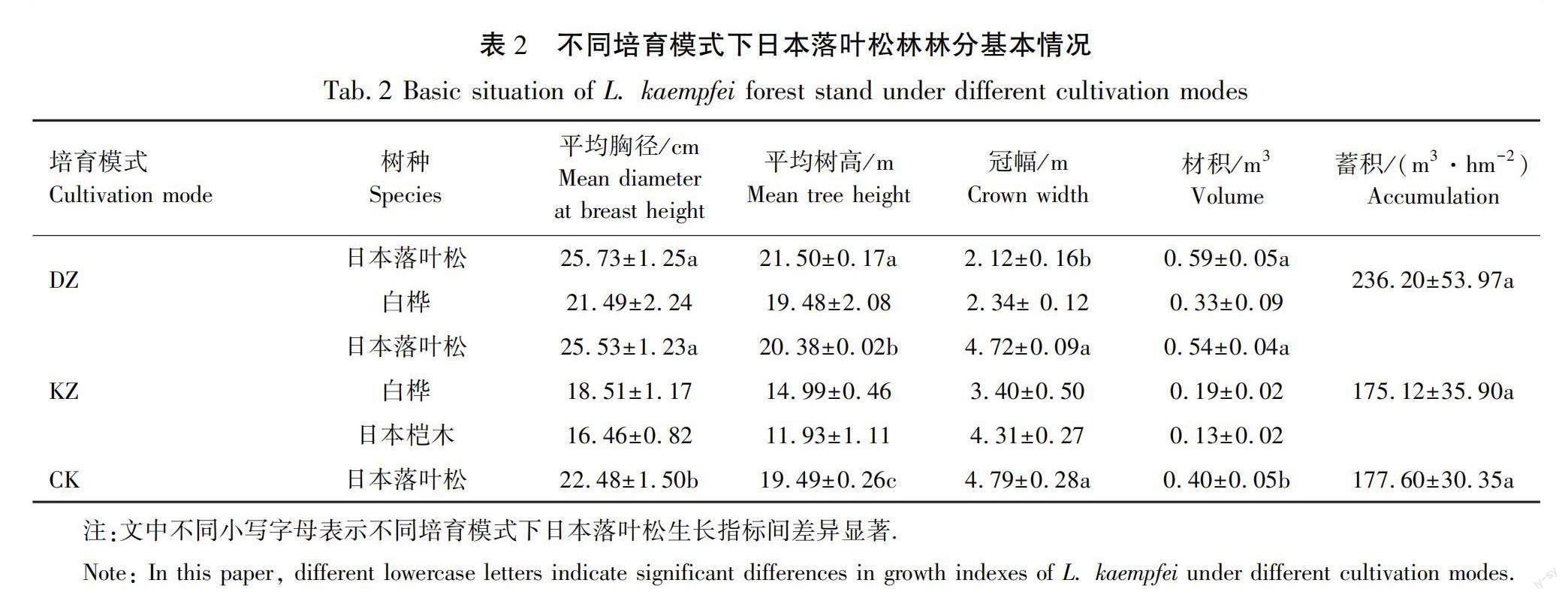
以研究区域内现有的3种典型培育模式下的日本落叶松人工林作为研究对象,其中日本落叶松-白桦带状混交培育模式为5行日本落叶松与3行白桦呈带状排列,研究中记为DZ,日本落叶松-白桦-日本桤木块状混交培育模式为日本落叶松、白桦和日本桤木分别以30 m×30 m呈品字形配置,且相同树种呈对角线排列,文中记为KZ,纯林培育模式为日本落叶松带状造林,文中记为CK。3种培育模式的林龄均为40 a,造林时采用水平阶整地,株行距为2 m×2 m,后经间伐,株行距调整为4 m×4 m。样地设置采用典型样地法,各培育模式的样地各布设3块,位于下坡位的样地为2块,中坡位则为1块,样地大
小为20 m×30 m,其中日本落叶松-白桦带状混交培育模式样地宽为1个林带宽度,日本落叶松-白桦-日本桤木块状培育模式样地设置在树种交错区,样地包含全部3个树种,样地和林分生长主要概况见表1和表2。在每个样地内根据对角线法选取5个1 m×1 m 的小样方调查草本植物,同时选择3个5 m×5 m的样方调查灌木,草本样方共75个,灌木样方45个,记录每种灌草种名、株数/丛数、高度和盖度,学名和生活型均利用“植物智”平台(http: //www. iplant. cn)进行确认。依据样地土层平均厚度,在5个草本调查处分0~20 cm和20~40 cm土层采集土壤,同一土层土壤均匀混合,装入土壤袋,风干过筛后进行土壤化学性质分析。
1.3 土壤化学性质测定方法
硝态氮和氨态氮含量采用扩散法测定;全氮含量的测定采用凯氏定氮法;全磷、有效磷采用钼锑抗比色法测定;土壤有机碳含量采用重铬酸钾氧化外加热法测定;全钾含量的测定采用HSO消解-火焰光度法测定;速效钾含量的测定采用NHOA浸提-火焰光度法测定;土壤干物质采用重量法测定;土壤pH测定采用电位法测定。
1.4 数据处理
1.4.1 材积计算
各树种材积计算见式(1)—式(3)。
1)日本落叶松
V=0.000 051 269 179DH。 (1)
2)白桦
V=0.000 051 935 16DH。 (2)
3)日本桤木
V=0.000 041 960 698DH。 (3)
式中:V为材积;D为树种胸径,cm;H为树高,m。
1.4.2 物种多样性指标计算
物种重要值、多样性指数和均匀度指数计算方法可参考方精云等的方法。
1)物种重要值。物种重要值计算方法见式(4)—式(8)。
2)多样性指数。物种丰富度指数(S)即为样地内出现物种的数量。Simpson多样性指数(S)、Shannon-Wiener多样性指数(S)的计算方法见式(9)—式(10)。
3)物种均匀度指数。以Simpson多样性指数为基础的Pielou群落均匀度指数(J)计算方法见式(11),以Shannon-Wiener多样性指数为基础的Pielou群落均匀度指数(J)计算方法见式(12)。
灌木层和草本层的重要值=(相对频度+相对多度+相对盖度)/3。 (4)
相对频度=(某一种的频度/所有种的频度之和)×100%。 (5)
相对多度=(某一种的个体数/全部种的个体数之和)×100% 。 (6)
相对盖度=(某一种的基面积之和/所有种的基面积之和)×100% 。 (7)
式中:P=n/N,P为物种i的相对多度,N为样方内所有物种的个体总数;n指样方中物种i的个体数,并且∑n=N。
1.4.3 土壤肥力评价指数计算
本研究采用乘除法的基本原理进行土壤肥力评价指数计算,见式(12)。由于各指标的参数意义、数量级和量纲不完全一致,故采用归一化处理式(13)将各参数值进行归一化变换。
X=(x-x)/(x-x)。 (13)
式中:L(g)为3种林分的土壤肥力评价指数;N(g)、N(g)、A(g)、T(g)、T(g)、A(g)、T(g)、A(g)、S(g)、S/T(g)、S/T(g)分别为土壤硝态氮、铵态氮、速效氮、全氮、全磷、速效磷、全钾、速效钾、有机碳含量及C∶N、C∶P;σ为上述各指标的标准差;X和x分别表示归一化前后各指标的数值;x和x分别为样本数据中的最大值和最小值;p代表pH的值。
1.5 数据分析
用SPSS 21.0对不同培育模式下日本落叶松林灌草多样性指数和土壤养分含量等进行单因素方差分析,用Duncan法进行多重比较,显著性水平均为0.05,以探寻不同培育模式间相关指标的差异。典型相关分析(Canonical Correlation Analysis,CCA)和冗余分析(Redundancy Analysis,RDA)分析用Canoco 5進行,灌草组成包括不同培育模式中重要值位于前五的物种,环境因子包括土壤养分(铵态氮、硝态氮、速效氮、速效氮、pH、全氮、全钾、全磷、速效钾、速效磷、有机碳、C∶N、C∶P和N∶P)和林分因子(平均胸径、平均树高、平均冠幅和单株材积);首先对灌草分布和多样性指数进行除趋势对应分析法(Detrended Correspondence Analysis,DCA)分析,发现灌草分布DCA第一轴大于4,分析其和环境因子的关系应采用CCA分析,而灌草多样性指数DCA第一轴小于3,应采用RDA分析;数据经对数转化后,利用Canoco 5的前向选择法确定影响灌草分布和多样性的环境因子,并用蒙特卡洛随机置换检验确定主要环境因子,置换检验次数设为499次。
2 结果与分析
2.1 不同培育模式下日本落叶松林灌草特征
2.1.1 日本落叶松林灌草重要值
基于各样地调查数据可知,3种培育模式下日本落叶松林灌草总数表现为DZ>KZ>CK;草本共有66种,属30科50属,其中日本落叶松-白桦带状混交培育模式林下草本有31种,属22科27属,日本落叶松-白桦-日本桤木块状混交培育模式林下草本有30种,属20科26属,纯林草本有27种,属20科26属;灌木共有15种,属11科13属,其中日本落叶松-白桦带状混交培育模式林下灌木有10种,属8科10属,日本落叶松-白桦-日本桤木块状混交培育模式林下灌木有9种,属8科8属,纯林灌木有9种,属7科8属。
不同培育模式日本落叶松林重要值位于前五的灌草科属和具体重要值如表3和图1所示。从优势物种来看,灌木层中东北山梅花(Philadelphus schrenkii)和狗枣猕猴桃(Actinidia kolomikta)重要值均大于0.16,草本层中猴腿蹄盖蕨(Athyrium multidentatum)和荨麻叶龙头草(Meehania urticifolia)的重要值均大于0.1,是日本落叶松-白桦带状混交培育模式的优势灌草,卫矛(Euonymus alatus)、毛榛子(Corylus mandshurica)和繁缕(Stellaria media)是日本落叶松-白桦-日本桤木块状混交培育的优势灌草,忍冬(Lonicera japonica)、卫矛(Euonymus alatus)、茶条槭(Acer tataricum subsp. ginnala)和和尚菜(Adenocaulon himalaicum)则是纯林的优势灌草。
从物种的共有性来看,不同培育模式下日本落叶松林灌木较为相似,卫矛、牛叠肚(Rubus crataegifolius)和山葡萄(Vitis amurensis)在3种培育模式中均出现,且重要值较高;草本植物共有性则较低,猴腿蹄盖蕨、龙常草(Diarrhena mandshurica)、鸡腿堇菜(Viola acuminata)、老鹳草(Geranium wilfordii)和野大豆(Glycine soja)在3种培育模式中均出现,仅猴腿蹄盖蕨重要值均位于前十。从物种特有性看,日本落叶松-白桦带状混交培育模式林下灌木出现辽东楤木(Aralia elata var. glabrescens)、东北茶藨子(Ribes mandshuricum)和五味子(Schisandra chinensis),纯林中出现葡萄(Vitis vinifera)和茶条槭(Acer tataricum subsp. ginnala),这些灌木均未在日本落叶松-白桦-日本桤木块状混交培育模式中出现;草本植物中,日本落叶松-白桦带状混交培育模式中独有蕨(Pteridium aquilinum var. latiusculum)、狼牙委陵菜(Potentilla cryptotaeniae)、日本散血丹(Physaliastrum echinatum)、山茄子(Brachybotrys paridiformis)和山莴苣(Lactuca sibirica)等10种草本,日本落叶松-白桦-日本桤木块状混交培育模式中独有大三叶升麻(Cimicifuga heracleifolia)、大叶堇菜(Viola diamantiaca)、大叶芹(Ostericum sieboldii)、广布鳞毛蕨(Dryopteris expansa)和宽叶山蒿(Artemisia stolonifera)等11种草本,日本落叶松纯林则出现宝珠草(Disporum viridescens)、北马兜铃(Aristolochia contorta)、蝙蝠葛(Menispermum dauricum)、和尚菜和辣蓼铁线莲(Clematis terniflora var. mandshurica)等11种草本。
2.1.2 日本落叶松林灌草多样性
不同培育模式下日本落叶松林灌草多样性指数见表4。总体来看,3种培育模式下日本落叶松林灌草多样性指数差异不明显。在草本层,除S表现为DZ>KZ>CK外,S、J、S和J均表現为CK>KZ>DZ,其中,仅有S、J和J在DZ和CK之间差异显著。在灌木层,S表现为CK>DZ>KZ,J表现为DZ>KZ>CK,其余S、J、S均表现为DZ>CK>KZ,除S在KZ和CK之间存在显著性差异外,其余均不存在显著性差异。
2.2 不同培育模式日本落叶松林土壤养分特征
2.2.1 日本落叶松林土壤养分含量
不同培育模式日本落叶松林土壤养分含量如图2所示。从不同土层厚度来看,硝态氮、铵态氮、速效氮、全氮、速效磷、速效钾和有机碳总体上呈现出随土层厚度加深而显著减少的趋势,而干物质则表现出相反的趋势,但差异不显著,全钾和pH在2个土层厚度间没有明显差异。从不同培育模式来看,铵态氮、速效氮、全氮、速效磷、全钾、速效钾和有机碳含量在2个土层厚度间均表现为DZ>CK>KZ,其中,DZ中0~20 cm土壤铵态氮、速效氮、全氮含量是分别是KZ的2.67、11.50、3.47倍,且2种培育模式间具有显著性差异;全磷含量在2个土层厚度间均表现为KZ最低,但0~20 cm土层表现为DZ最高,在20~40 cm土层则表现为CK最高,且存在显著性差异;pH在2个土层厚度间均表现为KZ最低,表明土壤酸化较为严重。干物质在2个土层厚度间均表现为KZ最高,CK次之,DZ最低。
2.2.2 日本落叶松林土壤化学计量特征
不同培育模式下日本落叶松林土壤化学计量特征见表5。从不同土层厚度来看,不同培育模式土壤0~20 cm的C∶N、C∶P和N∶P均高于20~40 cm土层,但C∶N差异不显著(P>0.05)。从不同培育模式来看,0~20 cm土层C∶N、C∶P和N∶P在3种培育模式中没有明显差异,20~40 cm土层的C∶N表现为CK>KZ>DZ,且CK和DZ具有显著性差异,C∶P和N∶P则表现为DZ>KZ>CK,在三者间均存在显著性差异。3种培育模式日本落叶松林灌草生长可能受到氮元素的胁迫,且土壤氮磷养分释放效率和有效性均低于我国森林土壤。
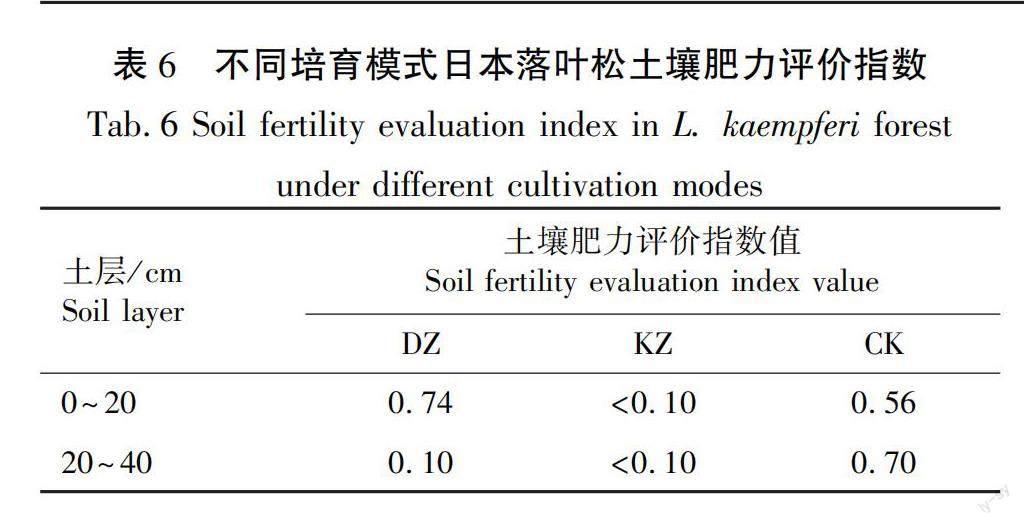
2.2.3 日本落叶松土壤肥力评价
本研究利用乘除法计算不同培育模式下日本落叶松林土壤肥力评价指数,结果见表6。日本落叶松-白桦-日本桤木块状混交培育模式土壤肥力最低,其中在0~20 cm土层,具体表现为DZ>CK>KZ,这与土壤多个化学元素含量的结果相一致,但在20~40 cm土层则表现为CK>DZ>KZ。综合林下土壤各元素含量和灌草植物根系深度分析,可判断日本落叶松-白桦带状混交培育模式土壤肥力最优。
2.3 环境因子对灌草分布、多样性的关系
2.3.1 环境因子对灌草分布的影响
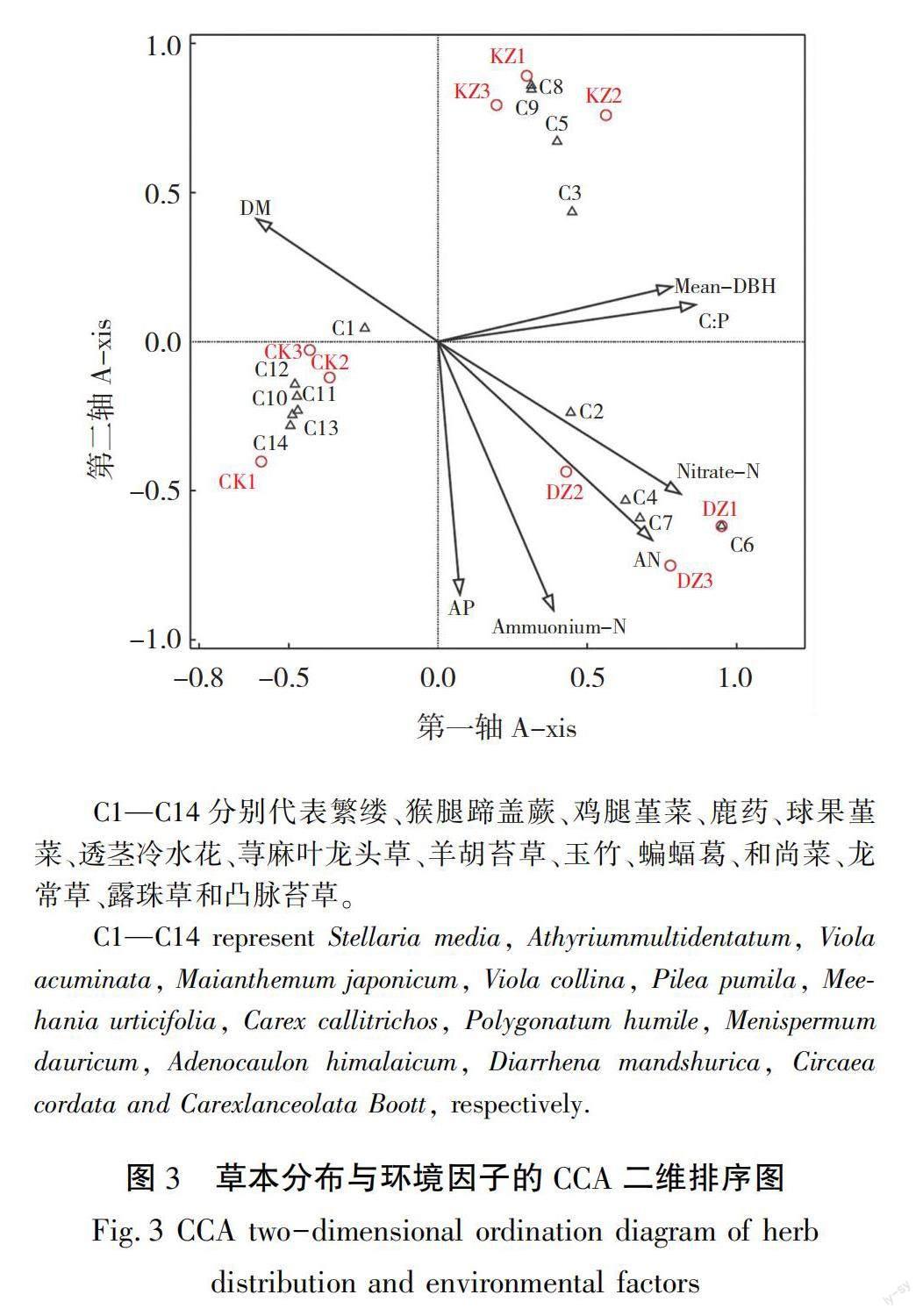
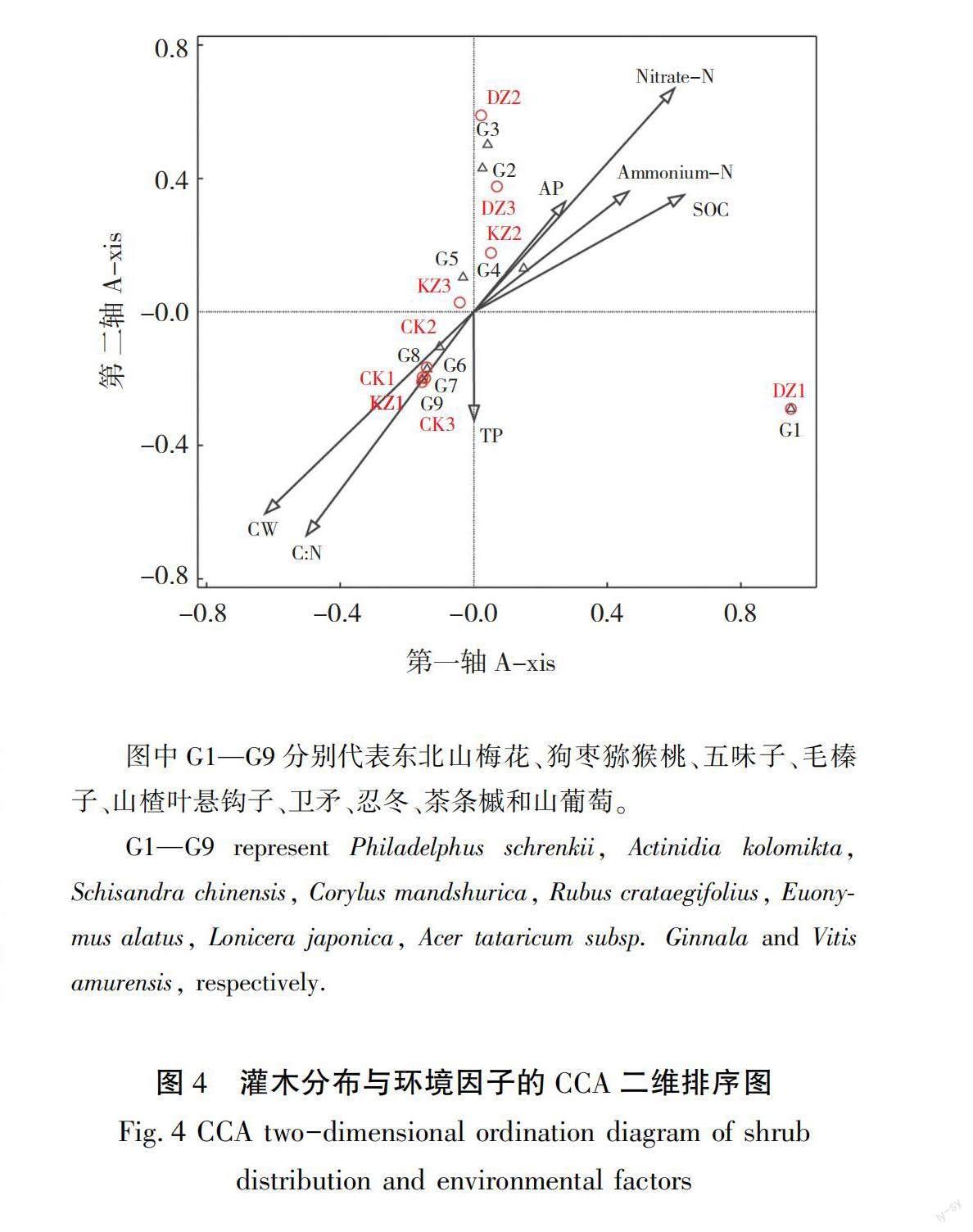
分别对辽东山区日本落叶松林灌木、草本分布和环境因子进行CCA排序,结果见表7和表8,如图3和图4所示,CCA前四轴总体解释率分别达98.0%(F为6.8,P为0.002)和96.3%(F=3.7,P=0.032),F为误差。排序结果可以解释环境因子和灌草分布间的关系。其中排序轴前两轴的解释率为60.8%和67.0%,与物种-环境因子的相关性均达到0.97以上,具有较好的生态意义,故可选择前两轴绘制二维排序图。3种培育模式样地在排序图中有规律地分布,反映了日本落叶松林灌草所处的环境梯度,总体来看,其与不同培育模式的土壤养分高低和林分条件一致;DZ草本主要分布在土壤氮磷养分含量高、林分平均胸径大,干物质低的环境梯度中,灌木主要分布在土壤有机碳、氮磷速效养分含量高的环境梯度中,KZ草本主要分布在养分含量低、林分平均胸径高、干物质大的环境梯度,灌木则对生境的倾向性不明显。CK草本则主要分布在养分含量低,林分平均胸径小的地方,灌木分布在土壤全磷含量高,C∶N高,冠幅小的环境梯度中。各优势灌草分布趋势与其培育模式所处环境梯度基本吻合,如鸡腿堇菜、鹿药和荨麻叶龙头草等草本主要分布在土壤氮磷养分含量高、林分平均胸径大,干物质低的环境梯度中,山楂叶悬钩子在3种培育模式中均出现,对环境没有明显的趋向性。蒙特卡洛随机置换检验结果表明,土壤硝态氮、铵态氮和速效磷是影响辽东山区日本落叶松林草本分布的主要环境因子,而灌木分布也主要受土壤硝态氮和铵态氮影响。
2.3.2 环境因子对灌草多样性的影响
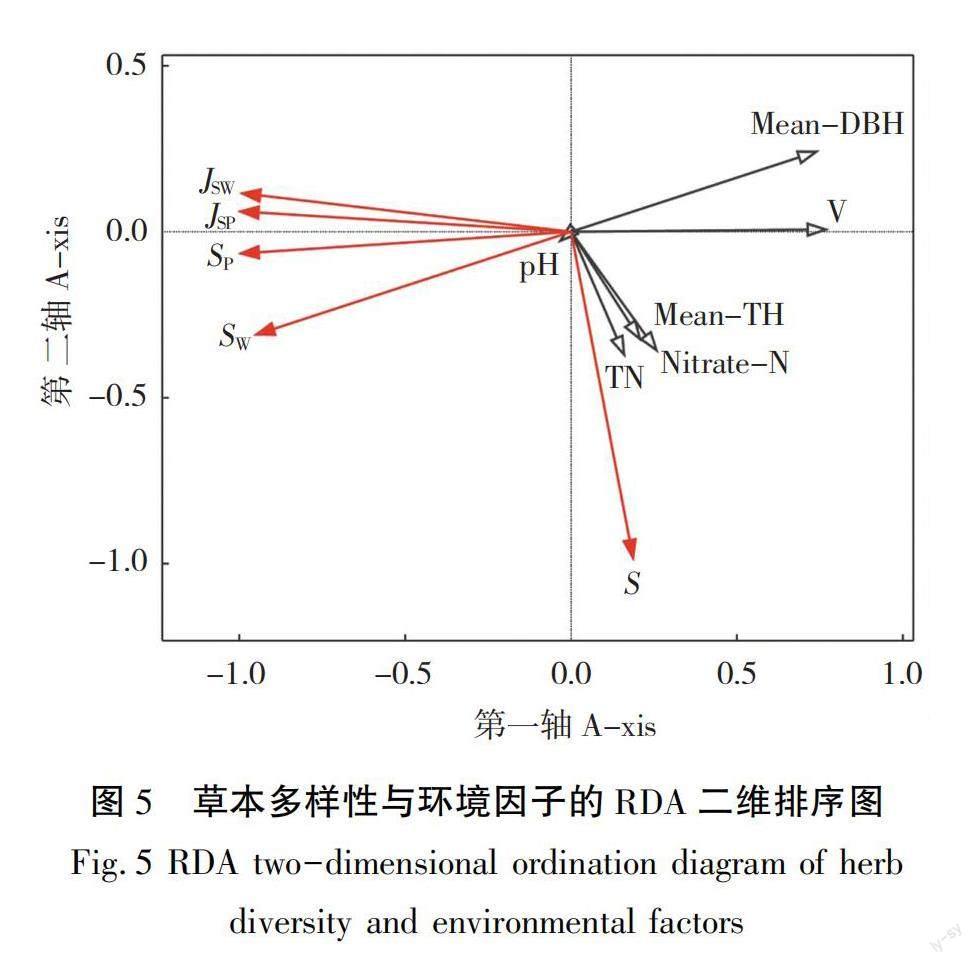
日本落叶松林灌木、草本多样性和环境因子RDA分析结果见表9和表10,如图5和图6所示,RDA前四轴总体解释率分别达96.8%(F=10.1,P=0.016)和97.6%(F=13.7,P=0.012),前两轴的解释率为96.6%和97.5%,与物种-环境因子的相关性均达到0.93以上,可选择前两轴绘制二维排序图。草本S、S、J、J随林分平均胸径、平均树高、单株材积、土壤铵态氮、全氮减小,pH增加而增加,S则总体上呈现与其他4个指数相反的规律。灌木S、S、J、J随土壤pH、速效磷、全氮增加,林分平均胸径、冠幅、土壤C∶N减小而增加。蒙特卡洛随机置换检验结果表明,单株材积是影响辽东山区日本落叶松林草本多样性的主要环境因子,而土壤C∶N是影响灌木多样性的主要环境因子。
3 讨论与结论
3.1 讨论
3.1.1 不同培育模式下日本落叶松林灌草多样性特征
林下植被是人工林生态系统的重要组成部分,其物种组成及多样性特征对生态系统健康发展和正向演替有着重要推动作用。本研究中,2种混交培育模式下日本落叶松林灌草丰富度大于纯林培育模式,这是因为相对于单一针叶纯林培育模式,针阔混交培育模式增加了环境异质性,为林下灌草的进入和生长创造了条件。本研究中草本S、S、J和J均表現为CK>KZ>DZ,RDA结果表明单株材积是影响草本多样性的主导环境因子,且其与多样性指数间呈现负相关,因此草本多样性呈现同单株材积相反的格局,但这与边丽宁的研究结果不同,原因在于不同林分条件和土壤养分下,主导物种多样性的环境因子具有明显差异。总体来看,3种培育模式下日本落叶松林灌草多样性差异不显著,尤其是灌木层植被,这是由于在相同气候条件下,各灌草经过长时间演替已形成稳定种群,特别是灌木逐渐趋于稳定,各培育模式的共有种和重要值均处于较高水平。
3.1.2 不同培育模式下日本落叶松林土壤养分特征
不同培育模式的林分具有独特的树种组成、冠层分布、根系分布、凋落物种类、凋落物分解性质和土壤质地等特征,从而使林地土壤养分也具有明显差异;本研究中日本落叶松-白桦带状混交培育模式的土壤肥力评价指数高于日本落叶松纯林培育模式,速效氮、全氮、速效磷、全钾、速效钾和有机碳等多个元素含量亦是如此,表明该培育模式有较好的养分固持作用,这也与目前大多数研究指出混交培育模式土壤的养分含量要优于针叶树种纯林培育的结论一致。本研究中日本落叶松-白桦-日本桤木块状混交培育模式土壤肥力最低,从CCA和RDA的结果来看,土壤养分与灌草分布、多样性相互影响,其中灌木多样性指数与土壤氮磷元素呈现负相关关系,表明林下植被生长需要损耗土壤养分,使得该模式下养分含量降低;也可能是因为引入白桦和日本桤木2种阔叶树种后对土壤养分吸收增多,导致土壤养分含量较低,但是林地土壤养分维持具有复杂的机制,与多种因素密切相关,目前对于块状混交林的相关研究鲜见报道,因此,需要持续开展植物养分吸收、凋落物归还、土壤酶和微生物活性等与林地养分转化循化能力密切相关的研究,揭示相关机理。
生态化学计量学理论引入到陆地生态系统后,土壤化学计量比广泛用于衡量土壤养分质量,C∶N∶P反映土壤氮、磷矿化为有效养分的能力及养分限制状况,低C∶N和C∶P表明氮磷养分释放快,有效性较高,本研究中林地土壤C∶N和C∶P均大于我国森林土壤(C∶N=13.7和C∶P=44.5),表明研究区域內不同培育模式下日本落叶松林土壤氮磷养分释放效率和有效性均低于我国森林土壤,但不同培育模式之间差异不明显。本研究中2个土层的N∶P平均值小于我国森林土壤(N∶P=7.9),且研究表明通常当N∶P<10时,植被生长受到氮元素的限制,因此研究区域内3种培育模式下的日本落叶松林可能受到土壤氮素的胁迫,需要加强土壤氮素的管理,这与中国北方森林更易受到氮元素限制的观点一致,也与本研究中灌草分布与环境因子的排序结果相互佐证。
3.1.3 不同培育模式下日本落叶松林灌草分布、多样性与环境因子的关系
林下植被、土壤养分和林分因子是相互影响的统一整体,三者在一定程度上具有紧密的相关性。本研究中草本的S、J、S和J等指数表现为CK>KZ>DZ,但土壤肥力指数在0~20 cm表现为DZ>CK>KZ,二者之间并不一致,原因在于灌草多样性受到诸土壤条件、林分状况及人为干扰等多种因素影响,且主要环境因子不同。CCA结果表明,土壤硝态氮、铵态氮和速效磷是影响辽东山区日本落叶松林灌草分布的主要因子,这是由于辽东山区日本落叶松林土壤氮磷养分释放效率和有效性均较低,该生境相当于环境筛,通过生境过滤筛选出具有较强氮磷利用能力的物种,使得植被的生长和分布格局不同。本研究发现单株材积是影响草本多样性的主要因子而非土壤养分,这可能是因为草本作为乔-灌-草分层的最下层,更易受到林分因子的影响;而土壤C∶N是影响灌木多样性的主要环境因子,这是因为相对于草本而言,林下灌木多样性虽然一定程度上受林分平均胸径和冠幅等林分因子限制,但其光环境条件优于草本植物,在氮素有效性较低的生境条件下,生长受到氮素影响,形成了C∶N限制格局。因此,可通过修枝等营林方式改善林内环境,并在森林经营时注意保存林下凋落物,以促进林地养分回归,保育林下物种多样性,发挥林下植被的生态效益。
3.2 结论
日本落叶松-白桦带状混交培育模式是适宜研究区域内日本落叶松培育的主要模式,此模式下灌草数量、灌木多样性、土壤养分及木材蓄积均处于较高水平。影响不同培育模式下日本落叶松林灌草分布和多样性的主要环境因子不同,可结合不同培育模式的主导因子进行林地管理,通过修枝等措施调整林分结构,保存林下植被和凋落物以促进养分回归,促使灌草多样性、林分因子和土壤养分协调发展,促进辽东山区日本落叶松林可持续经营。
【参 考 文 献】
[1]屈红军,牟长城,李婉姝.落叶松人工林群落结构特征及自然化经营[J].南京林业大学学报(自然科学版),2010,34(4):153-156.
QU H J, MU C C, LI W S. A study on the naturalization and community structure characteristics of Larix gmelinii plantation[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2010, 34(4): 153-156.
[2]刘胜涛,高鹏,刘潘伟,等.泰山森林生态系统服务功能及其价值评估[J].生态学报,2017,37(10):3302-3310.
LIU S T, GAO P, LIU P W, et al. An ecosystem services assessment of Tai Mountain[J]. Acta Ecologica Sinica, 2017, 37(10): 3302-3310.
[3]刘欣,彭道黎,邱新彩.华北落叶松不同林型土壤理化性质差异[J].应用与环境生物学报,2018,24(4):735-743.
LIU X, PENG D L, QIU X C. Differences in soil physicochemical properties between different forest types of Larix principis-rupprechtii[J]. Chinese Journal of Applied and Environmental Biology, 2018, 24(4): 735-743.
[4]冯健,王骞春,陆爱君,等.辽东山区落叶松-水曲柳混交林植物多样性和土壤特性研究[J].西北农林科技大学学报(自然科学版),2021,49(6):27-37.
FENG J, WANG Q C, LU A J, et al. Plant diversity and soil characteristics of larch-manchurian ash mixed stand in eastern Liaoning[J]. Journal of Northwest A & F University (Natural Science Edition), 2021, 49(6): 27-37.
[5]冯健,高慧淋,王骞春,等.辽东山区落叶松-水曲柳混交林及其纯林生长与生物量分配特征[J].东北林业大学学报,2021,49(7):22-27,32.
FENG J, GAO H L, WANG Q C, et al. Growth and biomass allocation of mixed larch-ash forest and its pure stand in the eastern mountainous area of Liaoning Province[J]. Journal of Northeast Forestry University, 2021, 49(7): 22-27, 32.
[6]陈笑,李远航,左亚凡,等.林分特征和土壤养分对林下草本物种多样性的影响[J].西北植物学报,2022,42(8):1396-1407.
CHEN X, LI Y H, ZUO Y F, et al. Effects of stand characteristics and soil nutrient characteristics on herbaceous diversity[J]. Acta Botanica Boreali-Occidentalia Sinica, 2022, 42(8): 1396-1407.
[7]刘博,潘存德,李贵华,等.喀纳斯泰加林火成演替群落数量分类与排序[J].生态环境学报,2019,28(10):1961-1973.
LIU B, PAN C D, LI G H, et al. Quantitative classification and sequencing of communities in pyrogenic succession of Kanas Taiga[J]. Ecology and Environmental Sciences, 2019, 28(10): 1961-1973.
[8]张荣,李婷婷,金锁,等.人为干扰对蒙顶山木荷次生林物种多样性及土壤理化性质的影响[J].浙江农林大学学报,2020,37(5):867-875.
ZHANG R, LI T T, JIN S, et al. Effects of human disturbance on species diversity and soil physiochemical properties of Schima superba community in Mengding Mountain[J]. Journal of Zhejiang A & F University, 2020, 37(5): 867-875.
[9]陳瑶,李云红,邵英男,等.阔叶红松林物种多样性与土壤理化特征研究[J].生态环境学报,2022,31(4): 679-687.
CHEN Y, LI Y H, SHAO Y N, et al. Study on species diversity and soil physical and chemical characteristics of broad-leaved Pinus koraiensis forest[J]. Ecology and Environmental Sciences, 2022, 31(4): 679-687.
[10]YANG K, ZHU J J, ZHANG M, et al. Soil microbial biomass carbon and nitrogen in forest ecosystems of Northeast China: a comparison between natural secondary forest and larch plantation[J]. Journal of Plant Ecology, 2010, 3(3): 175-182.
[11]党乾顺,隋心,陈福元,等.兴安落叶松林土壤养分与酶活性的动态分析[J].森林工程,2022,38(5):18-28.
DANG Q S, SUI X, CHEN F Y, et al. Dynamic analysis of soil nutrients and enzyme activities in Larix gmelinii forest[J]. Forest Engineering, 2022, 38(5): 18-28.
[12]夏成财,刘忠玲,王庆成,等.16年生落叶松白桦纯林与混交林林分生长量及生物量对比[J].东北林业大学学报,2012,40(10):1-3.
XIA C C, LIU Z L, WANG Q C, et al. Comparison of the growth and biomass production of monoculture and mixed stands of Larix gmelinii and Betula platyphylla[J]. Journal of Northeast Forestry University, 2012, 40(10): 1-3.
[13]方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548.
FANG J Y, WANG X P, SHEN Z H, et al. Methods and protocols for plant community inventory[J]. Biodiversity Science, 2009, 17(6): 533-548.
[14]方精云,沈泽昊,唐志尧,等.“中国山地植物物种多样性调查计划”及若干技术规范[J].生物多样性,2004,12(1):5-9.
FANG J Y, SHEN Z H, TANG Z Y, et al. The protocol for the survey plan for plant species diversity of China's Mountains[J]. Chinese Biodiversity, 2004, 12(1): 5-9.
[15]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000.
LU R K. Methods of soil agrochemical analysis[M]. Beijing: China Agriculture Science and Technology Press, 2000.
[16]郑颖,冯健,于世河,等.辽东山区不同密度落叶松人工幼龄林林木生长和土壤养分特性[J].中南林业科技大学学报,2022,42(1):94-103.
ZHENG Y, FENG J, YU S H, et al. Study on the forest growth and soil nutrient characteristics of Larix spp. plantations with different densities in mountainous area of Liaodong[J]. Journal of Central South University of Forestry & Technology, 2022, 42(1): 94-103.
[17]宁坤,刘笑平,林永红,等.白桦子代遗传变异与纸浆材优良种质选择[J].植物研究,2015,35(1):39-46.
NING K, LIU X P, LIN Y H, et al. Germplasm selection of the progeny genetic variation and superior pulpwood of Betula platyphylla[J]. Bulletin of Botanical Research, 2015, 35(1): 39-46.
[18]国家林业局.立木材积表(LY/T 1353-1999)[S].北京:中国标准出版社,1999:48-54.
STATE FORESTRY ADMINSTRATION. Tree volume tables(LY/T1353-1999)[S].Beijing:China Standard Press,1999:48-54.
[19]曹小玉,李际平,闫文德.杉木林改造前后的土壤肥力對比分析及综合评价[J].土壤通报,2016,47(5):1231-1237.
CAO X Y, LI J P, YAN W D. Comprehensive evaluation and variation of soil fertility before and after reconstruction with broadleaved trees in Cunninghamia lanceolata plantations[J]. Chinese Journal of Soil Science, 2016, 47(5): 1231-1237.
[20]张贾宇,佘婷,鄂晓伟,等.杨树人工林幼林阶段林下植被管理对土壤微生物生物量碳、氮酶活性的影响[J].生态学报,2021,41(24):9898-9909.
ZHANG J Y, SHE T, E X W, et al. Effects of understory vegetation management on soil microbial biomass carbon and nitrogen and extracellular enzyme activities in the early stages of poplar plantation[J]. Acta Ecologica Sinica, 2021, 41(24): 9898-9909.
[21]吴昊,肖楠楠,林婷婷.秦岭松栎林功能多样性与物种多样性和环境异质性的耦合关系[J].生态环境学报,2020,29(6):1090-1100.
WU H, XIAO N N, LIN T T. Relationships between functional diversity and species diversity of pine-oak mixed forest in Qinling Mountains and their environmental explanations[J]. Ecology and Environmental Sciences, 2020, 29(6): 1090-1100.
[22]边丽宁.华北落叶松纯林与混交林生长、林下植被和枯落物差异分析[D].保定:河北农业大学,2018.
BIAN L N. Difference of growth, understory vegetation and litter in pure and mixed forests of Larix principis rupprechti[D]. Baoding: Hebei Agricultural University, 2018.
[23]刘凯,陈乾,王希贤,等.不同林龄福建柏混交林与纯林土壤养分的动态变化[J].福建农林大学学报(自然科学版),2022,51(2):185-194.
LIU K, CHEN Q, WANG X X, et al. Dynamics of soil nutrients in Fokienia hodginsii-Pinus massoniana mixed forest and Fokienia hodginsii pure forest at different ages[J]. Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 2022, 51(2): 185-194.
[24]杨斌,王家学,郭小玮,等.景谷县思茅松人工林土壤养分特征与肥力评价[J].西部林业科学,2023,52(4):7-12.
YANG B, WANG J X, GUO X W, et al. Soil nutrient characteristics and fertility evaluation of Pinus kesiya var. langbianensis plantations in Jinggu County[J]. Journal of West China Forestry Science, 2023, 52(4): 7-12.
[25]曹小玉,赵文菲,李际平,等.中亚热带几种典型森林土壤养分含量分析及综合评价[J].生态学报,2022,42(9):3525-3535.
CAO X Y, ZHAO W F, LI J P, et al. A comprehensive evaluation of soil nutrients in main typical forests in central-subtropical China[J]. Acta Ecologica Sinica, 2022, 42(9): 3525-3535.
[26]趙航,贾彦龙,王秋凤.中国地带性森林和农田生态系统C-N-P化学计量统计特征[J].第四纪研究,2014,34(4):803-814.
ZHAO H, JIA Y L, WANG Q F. Statistical characteristics of C-N-P stoichiometry in Chinese zonal forest and farmland ecosystems[J]. Quaternary Sciences, 2014, 34(4): 803-814.
[27]朱秋莲,邢肖毅,张宏,等.黄土丘陵沟壑区不同植被区土壤生态化学计量特征[J].生态学报,2013,33(15):4674-4682.
ZHU Q L, XING X Y, ZHANG H, et al. Soil ecological stoichiometry under different vegetation area on loess hilly gully region[J]. Acta Ecologica Sinica, 2013, 33(15): 4674-4682.
[28]HUANG W J, LIU J X, WANG Y P, et al. Increasing phosphorus limitation along three successional forests in Southern China[J]. Plant and Soil, 2013, 364(1): 181-191.
[29]庞宏东,李玲,崔鸿侠,等.不同立地因子对泡桐人工林生长量及土壤理化性质的影响[J].东北林业大学学报,2021,49(7):28-32.
PANG H D, LI L, CUI H X, et al. Effects of different site factors on the growth and soil physical and chemical properties of Paulownia plantation[J]. Journal of Northeast Forestry University, 2021, 49(7): 28-32.
[30]刘道锟,孙海龙,甘秋妹,等.大兴安岭干旱阳坡不同植被退化阶段土壤理化性质与物种多样性研究[J].森林工程,2016,32(2):1-6.
LIU D K, SUN H L, GAN Q M, et al. Soil physicochemical properties and plant species diversity of different vegetation degradation stages in arid sunny-slope of Great Xing'an Mountains[J]. Forest Engineering, 2016, 32(2): 1-6.
[31]谭一波,申文辉,付孜,等.环境因子对桂西南蚬木林下植被物种多样性变异的解释[J].生物多样性,2019,27(9):970-983.
TAN Y B, SHEN W H, FU Z, et al. Effect of environmental factors on understory species diversity in Southwest Guangxi Excentrodendron tonkinense forests[J]. Biodiversity Science, 2019, 27(9): 970-983.
