规格和饥饿对刺参集群行为的影响
2023-11-21赖明想林承刚杨红生
赖明想, 林承刚, 杨红生
规格和饥饿对刺参集群行为的影响
赖明想1, 2, 3, 4, 5, 6, 林承刚1, 2, 3, 4, 5, 6, 杨红生1, 2, 3, 4, 5, 6
(1. 中国科学院海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071; 2. 青岛海洋科学与技术试点国家实验室海洋生态与环境科学功能实验室, 山东 青岛 266237; 3. 中国科学院海洋大科学研究中心, 山东 青岛 266071; 4. 中国科学院海洋牧场工程实验室, 山东 青岛 266071; 5. 中国科学院大学, 北京 100049; 6. 山东省实验海洋生物学重点实验室, 山东 青岛 266071)
本文通过延时摄影技术探究了不同刺参规格(小规格、中规格和大规格)及饥饿状态(饥饿0 d、5 d、10 d和15 d)对其集群行为的影响。研究发现: 不同规格刺参的聚集率和标准聚集规模之间存在显著差异(<0.05), 且呈现出随刺参规格增大而升高的趋势, 表明大规格刺参更倾向于聚集, 且形成的集群规模更大; 除饥饿10 d和饥饿15 d的组外, 各处理组之间刺参聚集率和标准聚集规模都具有显著差异(<0.05), 随着饥饿时间的延长, 刺参聚集率和标准聚集规模呈先降低, 后升高, 再保持稳定的规律。上述结果查明了规格和饥饿对刺参集群行为的影响, 可为丰富刺参行为学研究及优化刺参增养殖管理策略提供理论依据。
刺参; 集群行为; 规格; 饥饿
刺参()属于棘皮动物门、海参纲、楯手目、刺参科、仿刺参属, 主要分布于西太平洋北部35° N~44° N的近岸海域[1]。作为海洋生态系统的“清道夫”, 刺参具有十分重要的生态价值, 此外, 刺参含有多糖、甾醇和脂肪酸等多种活性物质, 具有很高的营养价值和经济价值, 是中国海水养殖产业的重要增养殖对象[2]。中国刺参养殖业始于20世纪50年代, 目前主要的养殖方式有工厂化养殖、池塘围堰养殖、筏式养殖和浅海底播养殖[3]。近年来, 随着增养殖规模的扩大, 海参产业经济效益不断提高, 根据《2022中国渔业统计年鉴》[4], 2021年全国海参总产量2.227×105t, 比2020年增长13.3%, 养殖面积2.474×105hm2, 比2020年2.427×105hm2增长1.9%。尽管产业发展迅速, 但目前刺参增养殖过程中仍有诸多难题亟待解决, 例如刺参底播养殖面临“去向难测、踪迹难觅”的困境, 难以突破刺参资源评估和成参高效采捕的技术瓶颈。集群行为在动物摄食、抵御敌害和繁殖过程中发挥着非常重要的作用, 对动物的时空分布具有重要影响, 为了解决上述难题, 实现底播刺参的可预测、可追踪和可控制, 必须对刺参的集群行为有充分的认知。
集群行为是指营群体生活的生物个体出于特定原因聚集成群的行为, 集群行为能为动物提供诸多益处, 如提高觅食效率、增强抵御捕食者能力以及有利于繁衍后代[5]。动物的集群行为影响因素众多, 包括光照、食物、种群密度和个体大小等[6], 如绿海胆在高密度(20 ind/m2)时的集群规模大于低密度(4 ind/m2)时的集群规模[7], 饥饿会影响牙鲆()的群体运动速度[8]。
目前, 关于刺参的行为学研究主要集中在个体水平, 如刺参的摄食行为与其个体大小及光周期有关[9], 与湍流和静流相比, 微弱的水流能加快刺参的运动速度[10], 繁殖会影响刺参的运动行为和肌肉生理[11]等。关于刺参的集群行为的研究, 多集中在环境因素及种内竞争对刺参群体的空间分布的影响, 如海州湾前三岛海域中刺参集中在有大型藻类分布的礁石区[12], 密度过高时刺参个体之间互相远离以缓解种内竞争压力[13], 而关于刺参规格和饥饿状态对其集群行为影响的研究尚未见报道。本研究探讨了不同规格和不同饥饿状态下刺参的集群行为特征, 旨在查明规格和饥饿对刺参集群行为的影响, 为底播增养殖刺参的科学管理和采捕提供理论依据。
1 材料与方法
1.1 材料
刺参取自山东省东营市通和海洋科技有限公司养殖池塘。刺参采集后在车间水池(6 m×2 m×1 m)中暂养半月以上, 暂养期间每日下午投喂过量饵料, 次日上午换水并清理水池底部的粪便和残余饵料。暂养所用海水为经120目尼龙滤网过滤的自然海水, 温度为8~10 ℃, pH为7.8~8.2, 盐度为29~31, 暂养期间使用增氧泵充气, 保持水体溶氧高于6 mg/L。待刺参适应环境开始正常活动和摄食后, 挑选外观完好、体质健康的刺参进行后续实验。
1.2 实验设计
1.2.1 规格对刺参集群行为的影响
实验在圆形帆布水池(半径0.6 m, 高0.8 m, 材质为PVC复合材料)中进行。根据湿重将刺参划分为小(4.37±0.63) g、中(22.81±2.62) g、大(102.41±5.74) g三种规格, 分别代表刺参浅海底播增养殖过程中的投苗期、养成期、成熟期。将特定规格刺参以10 ind/m2的密度放入水池圆心半径0.3 m范围内, 水池水深0.5 m, 初始状态下刺参互不接触且刺参个体之间的最小距离为1~2倍刺参体长(图1)。借助延时摄像机(Brinno TLC-200)记录刺参集群行为状况, 拍摄期间不投饵不换水, 环境条件与暂养期间保持一致, 延时摄像机参数设置为每2 h拍摄1帧, 每次记录时长为24 h。每个处理设置5次重复。
1.2.2 饥饿对刺参集群行为的影响
实验在圆形帆布水池(半径0.6 m, 高0.8 m, 材质为PVC复合材料)中进行。将大规格刺参进行为期15 d的饥饿处理, 饥饿处理期间不投饵, 海水温度、盐度和pH与暂养期间保持一致。由于刺参会摄食自身产生的粪便, 需要在每日换水的同时清理刺参粪便。分别在饥饿处理第0 d、第5 d、第10 d和第15 d时, 将刺参以10 ind/m2的密度放入水池圆心半径0.3 m范围内, 水池水深0.5 m, 初始状态下刺参互不接触且刺参个体之间的距离为1~2倍刺参体长(图1), 借助延时摄像机(Brinno TLC-200)记录刺参集群行为状况, 拍摄期间不投饵不换水, 环境条件与饥饿处理期间保持一致, 延时摄像机参数设置为每2 h拍摄1帧, 每次记录时长为24 h。每个处理设置3次重复。

图1 刺参初始位置示意图
1.3 数据计算
拍摄结束后, 分析各处理组的视频, 每2 h统计一次刺参的聚集率和标准聚集规模, 得到12组数据, 将12组数据的平均值作为该处理组的聚集率(aggregation rate)和标准聚集规模(typical aggregation size)[14]。
聚集率指在特定时间特定空间内的参与聚集的个体在全部个体中的占比, 参与聚集的刺参个体身体处于接触状态, 聚集率越高说明参与聚集的个体越多, 其计算公式如下:

其中,为刺参总数量,为参与聚集的个体数量。
由于在聚集率相同的情况下个体仍可产生不同的聚集状态, 因此还需引入另一评价指标, 标准聚集规模。标准聚集规模指个体聚集规模的大小, 用于表征个体的紧凑程度, 其计算公式如下:
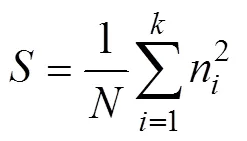
其中,为样本数量, 样本分为单独个体和聚集个体两种类型, 聚集个体处于身体接触状态,为所有样本中刺参数量总和,n为第个样本中刺参的数量[14]。
1.4 统计分析
实验数据用平均值±标准误差(mean±SE)表示, 对数据进行Shapiro-Wilk正态性检验和Levene’s同质性检验。使用单因素方差分析方法分析不同规格及不同饥饿状态下刺参的聚集率和标准聚集规模。上述统计分析均使用SPSS Statistics 26.0软件进行, 差异显著性水平设为0.05。
2 结果与分析
2.1 三种规格刺参的聚集率和标准聚集规模
不同规格刺参的聚集率和标准聚集规模如图2所示。小规格、中规格和大规格刺参的聚集率分别为(18.67±1.62)%、(48.83±0.73)%和(55.33±0.77)%, 三种规格刺参的聚集率之间存在显著差异(<0.05), 刺参的聚集率随规格的增大而升高。刺参的标准聚集规模也受刺参规格的影响, 小规格、中规格和大规格刺参的分别为(1.23±0.05)头、(1.71±0.06)头和(2.17±0.08)头, 标准聚集规模随刺参规格的增大而增加, 三种规格刺参的标准聚集规模之间存在显著差异(<0.05)。
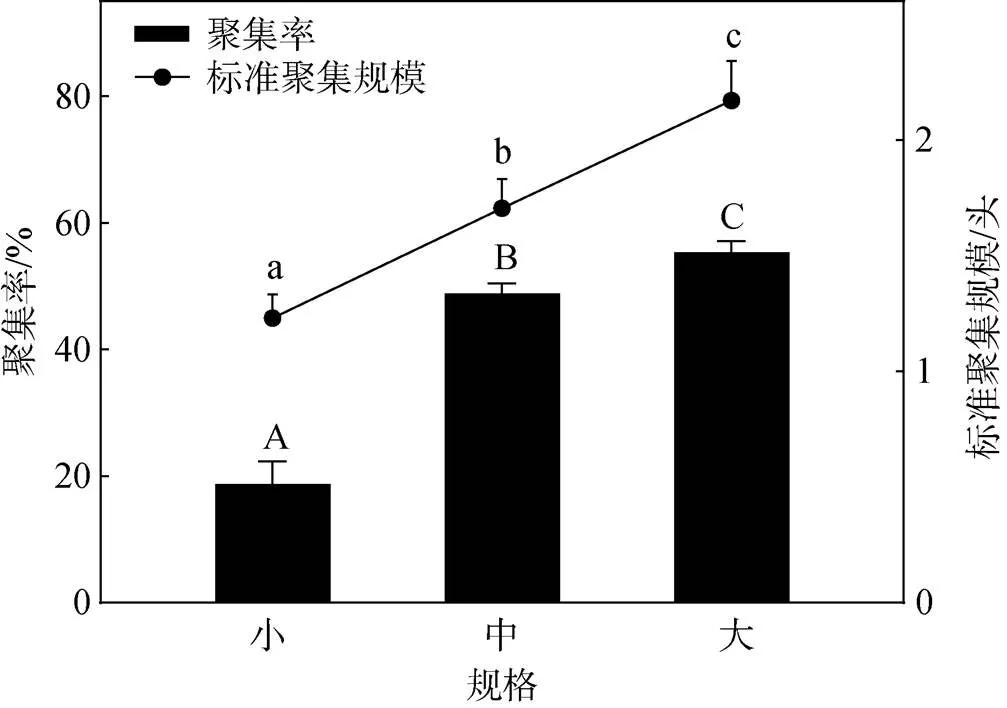
图2 不同规格刺参的聚集率和标准聚集规模
2.2 不同饥饿状态下刺参的聚集率和标准聚集规模
不同饥饿状态下刺参的聚集率和标准聚集规模如图3所示。随着饥饿时间的延长, 刺参的聚集率先降低, 后升高, 再保持稳定。饥饿5 d的刺参聚集率显著低于饥饿0 d、10 d和15 d的刺参(<0.05), 饥饿10 d和15 d的刺参的聚集率显著高于饥饿0 d的刺参(<0.05), 但饥饿10 d和15 d的刺参聚集率之间不存在显著差异(>0.05)。不同饥饿状态下刺参的标准聚集规模呈现出与聚集率相同的变化规律, 除饥饿10 d和15 d组之间外, 其余各组之间刺参标准聚集规模都存在显著差异(<0.05)。

图3 不同饥饿状态下刺参的聚集率和标准聚集规模
3 讨论
3.1 规格对刺参集群行为的影响
规格可以代表动物的生长阶段, 处在不同生长阶段的同种生物的行为可能存在显著差异。例如, 规格对雄性金乌贼的求偶和交配行为有显著影响, 大规格雄性金乌贼的交配成功率随着群体中小规格雄性金乌贼的增加而降低[15]; 不同规格双齿围沙蚕的营养盐溶出效率、轴向爬行速度、泵水量和泵水速率差异显著[16]。研究表明, 当刺参密度超过10 ind/m2时, 刺参聚集率会因种内竞争加剧而显著下降, 为避免种内竞争的影响, 本研究将各组的刺参密度设置为10 ind/m2[13]。本研究中, 在密度相同的条件下, 不同规格刺参的聚集率之间和标准聚集规模之间都存在显著差异, 刺参的聚集率和标准聚集规模随刺参规格的增大而升高, 即大规格刺参更倾向于聚集, 且形成的集群规模更大。关于棘皮动物海胆的集群行为研究也发现了相似的结果, 研究表明, 与小规格海胆相比, 大规格海胆更容易聚集成群且群体更为紧凑[17]。已有研究发现, 不同规格刺参的运动能力存在差异, 中规格和大规格刺参的运动速度与运动时间均高于小规格刺参, 规格大的刺参表现出更强的运动能力[18], 因而在一定空间内更活跃的大规格刺参相互接触并聚集成群的概率更大。此外, Sun等[9]的研究表明在摄食过程中大规格刺参触手的伸缩速率显著高于中规格刺参和小规格刺参的触手伸缩速率, 大规格刺参的摄食速度更快; 而Holtz等[19]的研究发现海参的摄食量与触手的伸缩速率之间存在正相关关系。规格越大的刺参能量需求越大, 所要摄取的食物量更多[9]。研究发现集群能够提高动物的搜寻食物的能力, 提升觅食效率[20], 本研究中规格越大的刺参聚集率越高, 标准聚集规模也越大, 大规格刺参更倾向于聚集成群, 可能是由于大规格刺参需要通过集群提高觅食效率, 获取更多食物, 以满足维持自身生命活动的能量需求。
3.2 饥饿对刺参集群行为的影响
在自然条件下, 受季节变动等因素的影响, 动物的食物资源常常处于波动状态; 在人工养殖条件下, 由于养殖密度较高而单位水体的食物资源有限, 动物面临的种内竞争加剧[12, 21]。无论是在自然条件还是人工养殖条件, 动物都可能因缺乏食物而处于营养不稳定的状态。营养状态与动物生理密切相关, 是影响动物行为的重要因素, 动物的诸多行为包括集群行为会随营养状态的改变而变化[22]。研究发现, 处于饥饿状态的三刺鱼和真鱥的集群性减弱[23]。本研究中, 饥饿5 d后刺参的聚集率和标准聚集规模均显著降低, 可能是刺参为提高觅食成功率而扩大搜索食物的范围, 使得刺参个体更为分散, 该结果与Hansen等[24]对杜氏虹银汉鱼()的研究结果一致, 饥饿状态下鱼群通过增大个体间距离来扩大获取食物的范围, 但也会因此提高个体被捕食的风险。而饥饿10 d后, 刺参聚集率和标准聚集规模升高, 刺参分布更为集中, 原因可能是饥饿导致刺参个体代谢减弱, 在食物极度匮乏条件下刺参为降低能量消耗而减少活动, 同时, 增加集群性可以提高抵御捕食者能力, 降低被捕食的风险。该结果与王蕾等报道的饥饿处理两周后中华倒刺鲃幼鱼群体的凝聚力和协调性提高的现象一致[25]。因此, 刺参的聚集率和标准聚集规模随着饥饿时间的延长先降低, 后升高, 再保持稳定的现象可能是刺参应对饥饿胁迫的一种集群反应, 是刺参在提高反捕食效益和降低觅食成本之间权衡的结果。刺参应对饥饿胁迫的集群行为策略可能与其他动物存在一定差异, 一方面是由于动物不同的运动特性, 与鱼类等水生生物相比, 刺参运动较为缓慢[10]; 另一方面, 不同动物所需的食物资源在时空分布上存在很大差异, 且不同动物对饥饿的耐受能力也不同[26]。本研究中, 饥饿10 d和饥饿15 d的刺参聚集率和标准聚集规模的差异不显著, 而高鹏程等发现刺参对饥饿的承受范围可能在10 d以内, 饥饿时间超出这一范围后, 即使再投喂饵料, 刺参也会出现持续性生长阻滞现象[27]。因此, 推测可能存在一个饥饿时间上限, 超过这一上限后刺参集群行为不再因饥饿水平的变化而发生明显改变, 而这一时间上限仍需进一步实验加以明确。
4 结论
综上所述, 规格和饥饿是影响刺参集群行为的重要因素。规格大的刺参聚集率更高, 且形成的集群规模更大。刺参饥饿状态的变化会引起集群行为的改变, 随着饥饿时间的延长, 刺参的聚集率和标准聚集规模先降低, 后升高, 再保持稳定, 这种应对饥饿胁迫的集群行为策略可能有利于提高群体的生存能力。
[1] 廖玉麟. 中国动物志: 棘皮动物门海参纲[M]. 北京: 科学出版社, 1997: 148-149. LIAO Yulin. Fauna Sinica, Phylum echinodermata, class holothuroidea[M]. Beijing: Science Press, 1997: 148-149.
[2] 姜健, 杨宝灵, 邰阳. 海参资源及其生物活性物质的研究[J]. 生物技术通讯, 2004(5): 537-540. JIANG Jian, YANG Baoling, TAI Yang. Studies on resources and bioactive substances of sea cucumber[J]. Letters in Biotechnology, 2004(5): 537-540.
[3] 黄华伟, 王印庚. 海参养殖的现状、存在问题与前景展望[J]. 中国水产, 2007(10): 50-53. HUANG Huawei, WANG Yingeng. Status, problems and prospects of sea cucumber culture[J]. China Fisheries, 2007(10): 50-53.
[4] 农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会. 2022中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2022: 23-50. Ministry of Agriculture and Pural Affairs of thr People′s Republic of China Fisheries Administration, National Fisheries Technology Extension Center, China Society of Fisheries. 2022 China Fishery Statistical Yearbook[M]. Beijing: China Agriculture Press, 2022: 23-50.
[5] PITCHER T J. Functions of shoaling behaviour in teleosts[M]. Boston: Springer, 1986: 294-337.
[6] SCHAERF T M, DILLINGHAM P W, WARD A J W. The effects of external cues on individual and collective behavior of shoaling fish[J]. Science Advances, 2017, 3(6): e1603201.
[7] HAGEN N T, MANN K H. Experimental analysis of factors influencing the aggregating behaviour of the green sea urchin(Müller)[J]. Journal of Experimental Marine Biology and Ecology, 1994, 176(1): 107-126.
[8] GWAK W-S, SEIKAI T, TANAKA M. Evaluation of starvation status of laboratory-reared Japanese flounderlarvae and juveniles based on morphological and histological characteristics[J]. Fisheries Science, 1999, 65(3): 339-346.
[9] SUN J, ZHANG L, PAN Y, et al. Feeding behavior and digestive physiology in sea cucumber[J]. Physiology & Behavior, 2015, 139: 336-343.
[10] PAN Y, ZHANG L, LIN C, et al. Influence of flow velocity on motor behavior of sea cucumber[J]. Physiology & Behavior, 2015, 144: 52-59.
[11] RU X, ZHANG L, LIU S, et al. Reproduction affects locomotor behaviour and muscle physiology in the sea cucumber,[J]. Animal Behaviour, 2017, 133: 223-228.
[12] 张宏晔, 许强, 刘辉, 等. 海州湾前三岛海域底播刺参群体特征初探[J]. 海洋科学, 2015, 39(6): 1-7. ZHANG Hongye, XU Qiang, LIU Hui, et al. Preliminary study on the property of bottom mariculture sea cucumber () group in Qiansan Islands, Haizhou Bay[J]. Marine Sciences, 2015, 39(6): 1-7.
[13] 辛孝科, 张立斌, 于正林, 等. 栖息地分布与种内竞争对刺参集群特征的影响[J]. 海洋科学, 2018, 42(5): 138-144. XIN Xiaoke, ZHANG Libin, YU Zhenglin, et al. Effects of habitat distribution and intraspecific competition on aggregation features of sea cucumber[J]. Marine Sciences, 2018, 42(5): 138-144.
[14] YU Z, HU Z, SONG H, et al. Aggregation behavior of juvenileand effects on seed production[J]. Journal of Oceanology and Limnology, 2020, 38(5): 1590-1598.
[15] 柏青青, 张东雪, 王林龙, 等. 金乌贼雄性规格和社群数量对求偶与交配行为的影响[J]. 中国水产科学, 2020, 27(2): 147-157. BAI Qingqing, ZHANG Dongxue, WANG Linlong, et al. Effects of male size on the reproductive behavior and competition of[J]. Journal of Fishery Sciences of China, 2020, 27(2): 147-157.
[16] 房景辉, 孟珊, 袁伟, 等. 温度对不同规格双齿围沙蚕运动行为特征的影响[J]. 中国水产科学, 2021, 28(10): 1251-1262. FANG Jinghui, MENG Shan, YUAN Wei, et al. Effects of temperature on the motion behavior ofof different sizes[J]. Journal of Fishery Sciences of China, 2021, 28(10): 1251-1262.
[17] BERNSTEIN B B, WILLIAMS B E, MANN K H. The role of behavioral responses to predators in modifying urchins'() destructive grazing and seasonal foraging patterns[J]. Marine Biology, 1981, 63(1): 39-49.
[18] 潘洋. 刺参运动节律行为的数量化研究[D]. 青岛: 中国科学院海洋研究所, 2015. PAN Yang. Quantitative research on motor behavioral rhythms of sea cucumber(Selenka)[D]. Qingdao: Institute of Oceanology, Chinese Academy of Sciences, 2015.
[19] HOLTZ E H, MACDONALD B A. Feeding behaviour of the sea cucumber(Echinodermata: Holothuroidea) in the laboratory and the field: relationships between tentacle insertion rate, flow speed, and ingestion[J]. Marine Biology, 2009, 156(7): 1389-1398.
[20] CLARK C W, MANGEL M. The evolutionary advantages of group foraging[J]. Theoretical Population Biology, 1986, 30(1): 45-75.
[21] 裴素蕊, 董双林, 王芳, 等. 限定食物资源下密度对刺参个体生长的影响[J]. 中国海洋大学学报(自然科学版), 2013, 43(3): 32-37. PEI Surui, DONG Shuanglin, WANG Fang, et al. Effects of sea cucumber () density on its growth under limited feed supply[J]. Periodical of Ocean University of China, 2013, 43(3): 32-37.
[22] 谢小军, 邓利, 张波. 饥饿对鱼类生理生态学影响的研究进展[J]. 水生生物学报, 1998(2): 181-188. XIE Xiaojun, DENG Li, ZHANG Bo. Advances and studies on ecophysiological effects of starvation on fish[J]. Acta Hydrobiologica Sinica, 1998(2): 181-188.
[23] ARBER B, HUNTINGFORD F A, CROMPTON D W T. The effect of hunger and cestode parasitism on the shoaling decisions of small freshwater fish[J]. Journal of Fish Biology, 1995, 47(3): 524-536.
[24] HANSEN M J, SCHAERF T M, WARD A J W. The influence of nutritional state on individual and group movement behaviour in shoals of crimson-spotted rainbowfish ()[J]. Behavioral Ecology and Sociobiology, 2015, 69(10): 1713-1722.
[25] 王蕾, 唐金玉, 覃英莲, 等. 饥饿对中华倒刺鲃幼鱼代谢、个性和集群的影响[J]. 生态学报, 2019, 39(3): 1095-1104. WANG Lei, TANG Jinyu, QIN Yinglian, et al. Effect of starvation on energy metabolism, fish behavior, and schooling behavior of[J]. Acta Hydrobiologica Sinica, 2019, 39(3): 1095-1104.
[26] 吴立新, 董双林. 水产动物继饥饿或营养不足后的补偿生长研究进展[J]. 应用生态学报, 2000(6): 943-946. WU Lixin, DONG Shuanglin. Advance in studies on compensatory growth of aquatic animals after starvation or undernutrition[J]. Chinese Journal of Applied Ecology, 2000(6): 943-946.
[27] 高鹏程, 周朝生, 吴洪喜, 等. 饥饿及恢复投喂对刺参生长、耗氧率和排氨率的影响[J]. 渔业科学进展, 2014, 35(1): 125-132. GAO Pengcheng, ZHOU Chaosheng, WU Hongxi, et al. Effects of starvation on growth, oxygen consumption and ammonia excretion rate of sea cucumber[J]. Progress in Fishery Sciences, 2014, 35(1): 125-132.
Effects of size and starvation on the aggregating behavior of sea cucumber
LAI Ming-xiang1, 2, 3, 4, 5, 6, LIN Cheng-gang1, 2, 3, 4, 5, 6, YANG Hong-sheng1, 2, 3, 4, 5, 6
(1. CAS Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. Laboratory for Marine Ecology and Environmental Science, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266237, China; 3. Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China; 4. Laboratory of Marine Ranching Engineering, Chinese Academy of Sciences, Qingdao 266071, China; 5. University of Chinese Academy of Sciences, Beijing 100049, China; 6. Shandong Province Key Laboratory of Experimental Marine Biology, Qingdao 266071, China)
This study investigated the effects of size (small, medium, and large) and starvation state (0, 5, 10, and 15 days of starvation) on the aggregating behavior ofusing time-lapse photography. Significant differences were found in the aggregation rate and typical aggregation size ofof different sizes (<0.05). The aggregation rate increased with the size of, indicating that large-sizedtended to aggregate, and their aggregating size was large. The aggregation rate and typical aggregation size ofin all treatment groups were significantly different (<0.05), except for those starved for 10 and 15 days. Further, the aggregation rate and typical aggregation size ofinitially decreased, then increased, and then attained a plateau as the starvation time was prolonged. This study revealed the influence of size and starvation state on the aggregating behavior of. These results provide a theoretical basis for improving the behavioral research ofand optimizing the management strategy for their breeding.
; aggregating behavior; size; starvation
Sep. 30, 2022
[National Natural Science Foundation of China, No. 42030408]
P735
A
1000-3096(2023)8-0001-06
10.11759/hykx20220930002
2022-09-30;
2022-11-25
国家自然科学基金重点项目(42030408)
赖明想(1998—), 男, 瑶族, 广西贺州人, 硕士研究生, 主要从事生物资源生态学研究, E-mail: laimingxiang@qdio.ac.cn; 杨红生(1964—), 通信作者, 男, 安徽霍邱人, 博士, 研究员, 长期从事养殖生态学、海参遗传育种与养殖、海洋牧场建设等研究, E-mail: hshyang@qdio.ac.cn
(本文编辑: 杨 悦)
