横断山区六种八居群鼠尾草属植物的核型分析
2023-11-16李文胜金泓燕黄愿如李志敏孙文光
李文胜, 金泓燕, 黄愿如, 李志敏,3, 孙文光,,3*
( 1. 云南师范大学 生命科学学院, 昆明 650500; 2. 中国科学院昆明植物研究所 生物多样性与生物地理学重点实验室, 昆明 650204; 3. 云南师范大学 云南省生物质能与环境生物技术重点实验室, 昆明 650500 )
横断山区位于“世界屋脊”青藏高原东南部,包括云南西北部、四川西部、西藏东部、青海南部以及甘肃西南部地区,面积约36.4万平方千米(李炳元, 1987)。横断山区因南北走向的山脉,海拔落差大,以及受印度洋暖湿气流的影响,形成了显著的沿海拔梯度变化的多种气候类型的生态环境,包括低海拔的干热河谷地区,中海拔的暖温带地区及高海拔的高山冰缘带地区,形成了横断山区丰富的物种多样性。正因如此,横断山区成为全球重要的生物多样性研究中心之一,也因物种多样性丰富而成为研究物种起源演化的天然实验基地(杨勤业等, 1988;曾广权等, 2000; 姚永慧等, 2010)。
唇形科(Lamiaceae)鼠尾草属(Salvia)是世界性广布的大属,广泛分布于中美洲、南美洲、西亚和东亚地区(Walker &Sytsma, 2007),全世界范围共计约980种(Hu et al., 2018)。西亚和地中海地区被认为是鼠尾草的原始分布中心(Masoud et al., 2010)。原产中国的鼠尾草种类有81种24变种,横断山区有37种13变种(中国科学院青藏高原综合科学考察队, 1993; Li &Hedge, 1994)。该属物种在生长形态、花形态等方面表现出显著的多样性。吴征镒和李锡文(1982)认为鼠尾草属的唇形花冠更适应昆虫传粉,因此是更进化和特化的类群,并且认为横断山地区是该属形成和多样化的中心之一。尽管部分研究人员对产于西喜马拉雅地区的鼠尾草进行了细胞学研究,但是对横断山区鼠尾草属植物的细胞学资料仍研究较少(Gill, 1970; Bhattacharya, 1978; Haque, 1980)。
细胞学核型分析是研究植物进化地位及种群间亲缘关系常用的方法之一,通过对中期细胞染色体组的数目、倍性、形态、大小、着丝粒位置、随体有无等特征分析,再通过计算臂比、染色体长度比、着丝粒指数、染色体长度变异系数以及着丝粒指数变异系数等一系列核型参数计算,来对物种进行分类学研究,分析其亲缘关系和进化地位。研究资料表明鼠尾草属的染色体数目具有多样化。Stewart(1939)对美国加利福尼亚地区5个亚属18种鼠尾草属植物的染色体数目进行研究,发现染色体基数为x=8、12、16。Fujita(1970)根据其染色体基数为x=11,认为鼠尾草属是唇形科中最原始的属,并且是唇形科中染色体基数最高的属。Alberto等(2003)研究了阿根廷地区12种鼠尾草属植物有丝分裂时期染色体形态和减数分裂时期的染色体,发现S.coccinea、S.farinacea、S.involucrate和S.microphylla为二倍体物种,染色体数目为2n=2x=22、20、22、22,S.cardiophylla、S.procurrens、S.splendens和S.uliginosa为四倍体物种,染色体数目分别为2n=4x=44、52、44、52,S.stachydifolia、S.pallida为六倍体物种,染色体数目均为2n=6x=66,S.guaranitica和S.rypara为八倍体物种,染色体数目均为2n=8x=88。Gill(1970)对喜马拉雅山系的20种鼠尾草属植物的染色体进行研究,发现鼠尾草属植物的染色体基数具有多样性,其中x=6、7、8是常见的染色体基数。Özdemir和enel(1999)报道了S.sclerea的染色体数目2n=22。赵红霞等(2006)报道了我国鼠尾草属植物丹参(S.miltiorrhiza)、黄花鼠尾(S.flava)和雪山鼠草(S.evansiana)的染色体数目2n=16和32。Yang等(2009)对我国横断山脉地区的6种鼠尾草属植物的染色体数目进行了研究,发现S.przewalskii为四倍体,染色体数目为2n=4x=32,S.castanea、S.flava、S.trijuga和S.yunnanensis为二倍体,染色体数目均为2n=2x=16,染色体基数为x=8。中国的鼠尾草物种染色体基数主要为x=8。经统计,全世界范围内鼠尾草属植物染色体数据已报道了223种(Ranjbar et al., 2015; Rice et al., 2015),中国地区鼠尾草属染色体数据已报道26种(含4变种1变型),其中横断山区15种(中国科学院青藏高原综合科学考察队, 1993; 赵红霞等, 2006; Yang et al., 2009; 洪培培等, 2011; Yang et al., 2014)。鼠尾草属细胞学研究依旧十分薄弱,而鼠尾草属的分子系统学分类与形态学分类的差异较大,细胞学研究可以为该属物种演化形成及物种多样性提供重要的参考资料(Adams, 1968; Weiss &Schneeweiss, 2013)。
鼠尾草属的属下分类系统自建立以来就一致存在争议,Bentham(1832)最早构建了鼠尾草属的属下分类系统,此后联合其他新发表的类群整合为4个亚属。此外Briquet在Bentham基础上进行修正,将鼠尾草属划分为8亚属17个组(Engler &Prantl, 1899),以上两个系统仍然是目前世界鼠尾草属分类相对完整的属下分类系统。国产鼠尾草属的分类系统由吴征镒和李锡文(1982)在Briquet系统基础上建立,他们依据鼠尾草属植物雄蕊特征将国产鼠尾草分为3个亚属,即弧隔鼠尾草亚属、荔枝草亚属和鼠尾草亚属,以及组和系的分类等级。随着分子生物学的发展,众多学者利用核基因组和叶绿体基因组进行分子系统树的构建(Walker &Sytsma, 2007; Hu et al., 2018),以此来对鼠尾草属进行修订。
为补充鼠尾草属植物细胞学染色体数据,在细胞学水平与基因组水平共同探讨鼠尾草属内分类学地位,本研究通过收集鼠尾草属已报道染色体数据资料,并对横断山区6种8居群鼠尾草属植物进行核型分析,还通过叶绿体基因组片段构建的鼠尾草属物种系统发育树,结合细胞学资料,讨论形态分类学与分子系统学之间的分类关系,探究横断山区物种在细胞学水平的进化方式。
1 材料与方法
1.1 实验材料
本研究中使用的鼠尾草属植物种子均采自横断山区,种子详细信息见表1,凭证标本均存放在中国科学院昆明植物研究所标本馆(KUN),细胞学凭证装片存放在云南师范大学(YNNU)植物细胞学研究室。

表1 本实验材料信息Table 1 Experimental material information in this study
1.2 实验方法
1.2.1 染色体制片步骤 选取发育良好、籽粒饱满的种子,用蒸馏水清洗种子后放到潮湿滤纸培养皿中,置于24 ℃恒温箱中萌发,待其根长至1 cm左右将根取下,装入有0.03%的8-羟基喹啉的离心管中,常温避光预处理1.5~2.5 h。放入装有现配卡诺氏固定液(冰乙酸∶无水乙醇=1∶3)的离心管中,置于4 ℃冰箱中冷藏20 h后转移至70%酒精中3 h。用1 mol·L-1盐酸于60 ℃下解离11 min。以上步骤之间需用蒸馏水反复冲洗3次将试剂洗净,最后放入装有卡宝品红的离心管中染色2 h。通过压片把制作好的片子放到光学显微镜(Olympus BX-53)下镜检,并挑选分裂中期细胞染色体形态清楚的细胞观察拍照。将染色体形态完好并分散均匀的片子使用中性树胶封装,制作永久封片。
1.2.2 核型分析方法 根据核型分析标准统计30个以上的中期细胞染色观察记录超过85%细胞恒定的染色体数目为该种的最终染色体数目 (李懋学和陈瑞阳, 1985)。选取6张染色体较为分散、形态良好且位于同一细胞内的图片,利用Adobe Photoshop CC添加5 μm标尺、裁剪及调整后利用染色体测量软件Karyotype V.2量取6个细胞的染色体长短臂长度、臂比等数据(Altinordu et al., 2016),进行同源染色体核型配对分析,然后根据6个细胞的核型数据利用Excel软件进行综合配对,并依据综合配对结果绘制核型模式图,并根据李懋学和陈瑞阳(1985)、Paszko(2006)、Peruzzi和Eroglu(2013)、Peruzzi和Altnordu(2014)等人的标准确定各个种的基本核型参数。并根据Levan等(1964)的染色体分类系统和Stebbins(1971)的标准确定间期核类型、核型公式和染色体的核型不对称类型等(孙文光等, 2019)。
1.2.3 鼠尾草属基数与倍性统计 本研究通过整理染色体数据库CCDB(Rice et al., 2015)结合Ranjbar等(2015)的统计研究,分别对鼠尾草属的染色体基数与倍性在世界、中国和横断山区范围内进行了统计,并进行统计图的绘制。
1.2.4 系统发育分析 通过美国国家生物技术信息中心(National Center for Biotechnology Information, NCBI)官方网站(https://www.ncbi.nlm.nih.gov)下载原产中国的并记录在《中国植物志》和FloraofChina中的鼠尾草属植物叶绿体标记基因数据(psbA-trnH、TrnL-TrnF、rbcL、ycf1-rps15),将其导入Geneious 9.0.2软件连接,并在此软件中对下载自NCBI的叶绿体数据进行处理,分别对不同基因序列开展多重MAFFT比对(Kazutaka et al., 2002)及多基因联合处理(Concatenate Sequences or Alignments)。使用IQ-TREE V.2.1.3版本构建进化树,经过检测最优模型为“TVM+F+R2”模型(Nguyen et al., 2015),通过文献查阅设置龙口花(Horminumpyrenaicum)作为外类群,基于最大似然法(maximum likelihood,ML)构建进化树。通过查询文献,将所有已经发表的染色体数据与本研究中鼠尾草属植物的染色体数据共同在分子进化树上进行体现。
2 结果与分析
2.1 六种八居群鼠尾草属植物核型特征
由染色体形态图、核型模式图及核型分析结果(图1,表2)可知,除甘西鼠尾草(S.przewalskii)为同源四倍体外,其他5种鼠尾草属植物皆为基数x=8的二倍体。着丝粒位置以近中部着丝粒(sm)为主,也有中部着丝粒(m)和近端部着丝粒(st)。核型不对称性除采自四川宝兴的甘西鼠尾草为3B型和采自云南丽江的黄花鼠尾草(S.flava)为2B型外,其余居群核型不对称性均为3A型。平均臂比范围为1.62~2.48,核型不对称系数为61.75%~70.95%,最长/最短染色体比值为1.66~2.17,臂比>2∶1染色体的百分比为25.00%~87.50%。其中黄花鼠尾草、粘毛鼠尾草(S.roborowskii)、雪山鼠尾草(S.evansiana)、毛地黄鼠尾草(S.digitaloides)皆在第一对染色体短臂上具随体,而采自四川宝兴甘西鼠尾草具4~6条数目不定的B染色体,这是首次在甘西鼠尾草内发现B染色体。6种8居群鼠尾草属植物的间期核型均为简单染色中心型。

a. 甘西鼠尾草(四川宝兴)(白色箭头指示为B染色体); b. 甘西鼠尾草(云南香格里拉); c. 黄花鼠尾草(四川宝兴); d. 黄花鼠尾草(云南丽江); e. 毛地黄鼠尾草(云南丽江); f. 西藏鼠尾草(西藏昌都); g. 雪山鼠尾草(云南德钦); h. 粘毛鼠尾草(四川乡城)。*表示该染色体包含随体。a. S. przewalskii(Baoxing, Sichuan) (The black arrow indicates the B chromosome); b. S. przewalskii(Shangri-La, Yunnan); c. S. flava(Baoxing, Sichuan;); d. S. flava(Lijiang, Yunnan); e. S. digitaloides(Lijiang, Yunnan); f. S. wardii(Changdu, Xizang); g. S. evansiana(Deqen, Yunnan); h. S. roborowskii(Xiangcheng, Sichuan). * indicates that the chromosome contains a satellite.图1 6种8居群鼠尾草属植物中期细胞图、配对染色体形态图及模式图Fig. 1 Metaphase cell, paired chromosome morphology and karyogram of six species of Salvia in eight populations
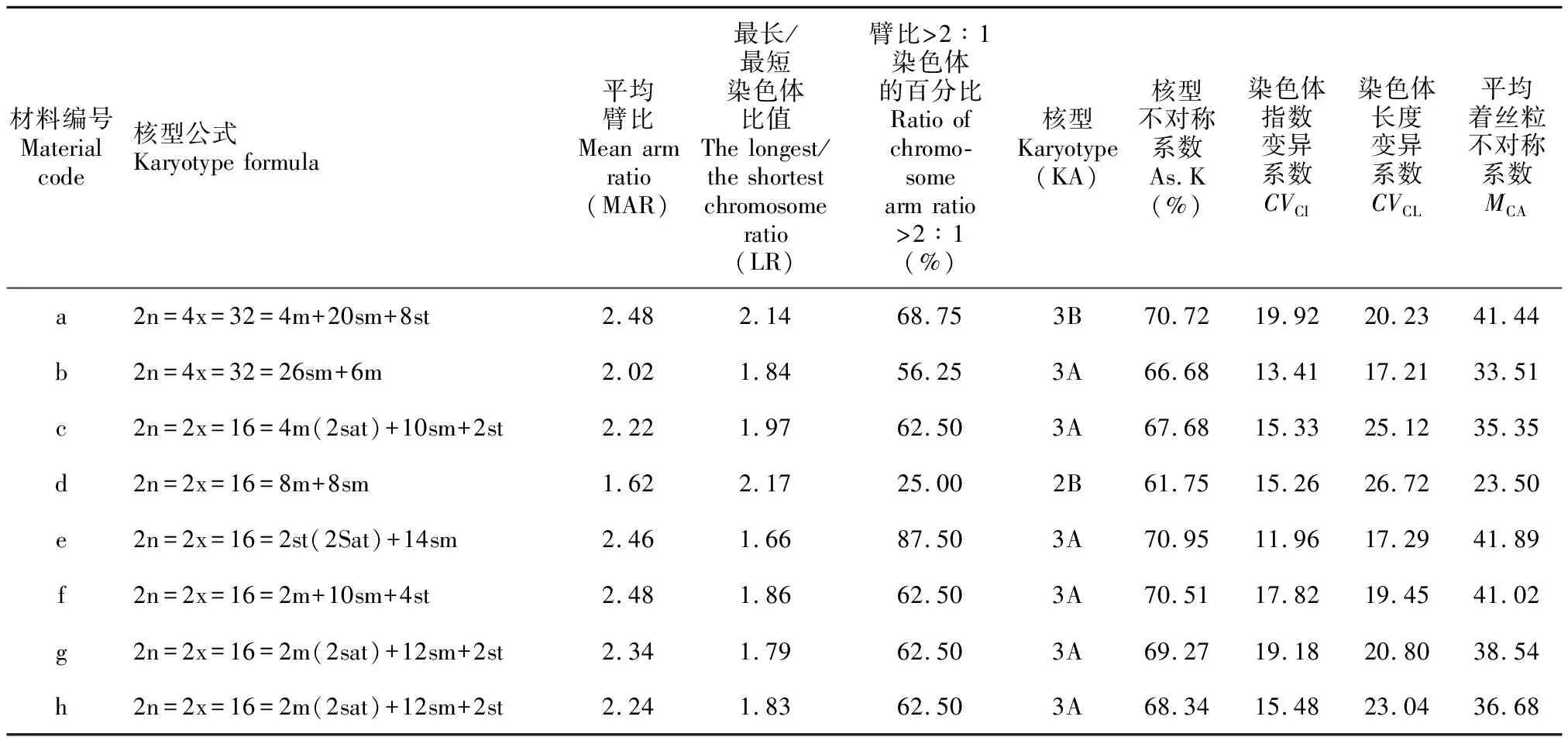
表2 鼠尾草属植物的染色体核型特征Table 2 Karyotype characteristics of Salvia chromosome in this study
2.2 鼠尾草属基数与倍性统计结果
染色体基数与染色体倍性统计结果如图2所示。从全世界范围来看,染色体基数以x =7、8、9、10、11为主,x=8和x=11合计约占总数的一半,染色体倍性主要以二倍体物种为主;产自中国地区的鼠尾草属物种染色体基数有x=7、8、10、11,主要以x=8为主,染色体倍性主要以二倍体为主。相比世界范围内的鼠尾草属,中国地区特别是横断山区的染色体基数更加集中且均以x=8为主要基数类型,染色体倍性统计中,无论在哪个范围内统计,二倍体物种都具有绝对的比例优势。
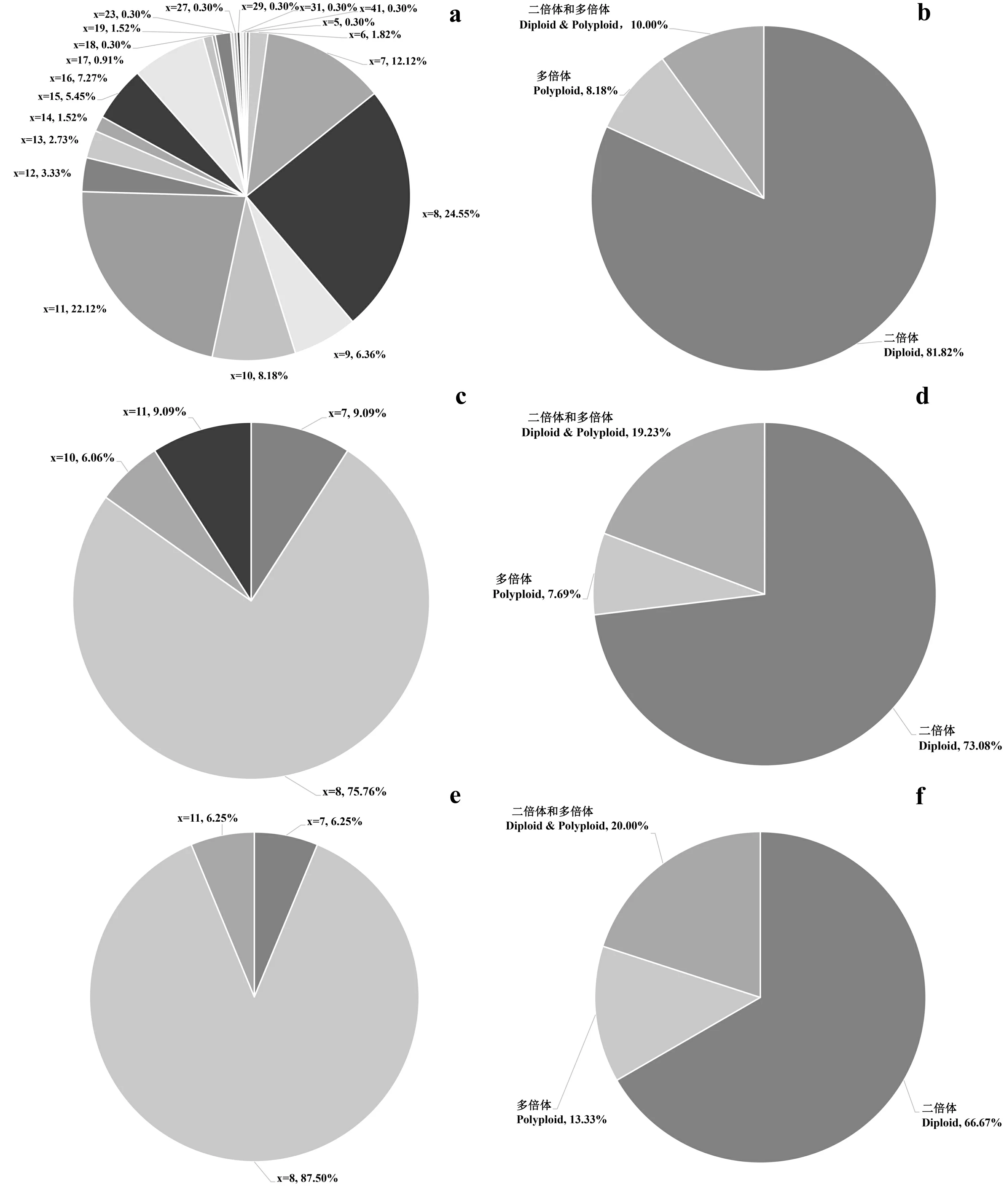
a. 世界鼠尾草属染色体基数统计; b. 世界鼠尾草属染色体倍性统计; c. 中国鼠尾草属染色体基数统计; d. 中国鼠尾草属染色体倍性统计; e. 横断山区鼠尾草属染色体基数统计; f. 横断山区鼠尾草属染色体倍性统计。a. Chromosome cardinal number statistic of Salvia in the world; b. Chromosome ploidy statistic of Salvia in the world; c. Chromosome cardinal number statistic of Salvia in China; d. Chromosome ploidy statistic of Salvia in China; e. Chromosome cardinal number statistic of Salvia in Hengduan Mountains; f. Chromosome ploidy statistic of Salvia in Hengduan Mountains.图2 鼠尾草属染色体基数与倍性所占比例图Fig. 2 Chromosome cardinal number and ploidy ratio chart of Salvia
2.3 系统发育分析结果
通过构建国产鼠尾草属62种分子进化树(图3),将鼠尾草属植物的染色体核型数据与系统发育树进行匹配,发现国产鼠尾草属植物染色体基数以x=8为主,倍性以二倍体为主。分子进化树在弧隔鼠尾草亚属(Subg.Salvia)和美洲鼠尾草亚属(Subg.Jungia)这两分支对于形态学分类的支持度最高,除三叶鼠尾草(S.trijuga)外,均属于弧隔鼠尾草亚属, 美洲鼠尾草亚属全部物种单独聚为一支; 而荔枝草亚属(Subg.Sclarea) 是支持率较低的亚属。在弧隔鼠尾草亚属这一分支上的染色体基数,已报道的除栗色鼠尾草(S.castanea)有x=8和x=11两种染色体基数外,其余的物种均为x=8,出现染色体组加倍的物种有雪山鼠尾草、短唇鼠尾草(S.brevilabra)和甘西鼠尾草,其中雪山鼠尾草又同时拥有二倍体物种。另外,雪山鼠尾草、三叶鼠尾草、圆苞鼠尾草(S.cyclostegia)、暗红鼠尾草(S.atrorubra)和新疆鼠尾草(S.deserta)这5个物种在进化树上表现出在各自的大进化支内都处在相对独立的单一分支中,与各自分支中的大部分物种具有相对较远的亲缘关系,且三叶鼠尾草与新疆鼠尾草没有与形态学分类中所属亚属的大部分物种聚为一支。美洲鼠尾草亚属的染色体基数和倍性均呈现出多样性的特点。而鼠尾草亚属(Subg.Allagospadonopsis)和荔枝草亚属中的染色体报道相对较少,且在与分子系统树匹配上也没有很好的支持,亟需进行细胞学的研究并且进行更加准确的分子进化树的构建。

黑色箭头表示非标识亚属物种; *表示Flora of China中的6种鼠尾草属植物,但未记录在《中国植物志》中;加粗标识物种表示本研究中进行细胞学分析的物种。Black arrow marks mean non-subgenus species; * means six Salvia species in Flora of China (FOC), but not recorded in Flora Reipublicae Popularis Sinicae; species identified in bold mean the species for cytological analysis in this study.图3 中国地区分布鼠尾草属植物基于叶绿体DNA联合数据构建的分子系统树(分支节点数字代表可能性)Fig. 3 Molecular phylogenetic tree of Salvia from China based on combined chloroplast DNA data (branch node numbers represent possibilities)
3 讨论与结论
鼠尾草属作为唇形科的冠部进化分支,在世界各地广泛分布,与其具有较高的适应能力密不可分。鼠尾草属植物的适应进化特性是由遗传物质对环境的适应所造成,因此鼠尾草属植物的进化特征也与其核型进化有关(黄艳波等, 2014)。本研究中通过收集全世界范围内的鼠尾草属染色体数据报道,统计得出,全世界范围内,鼠尾草属植物染色体数据报道率约为23%,中国地区报道率为32.10%,横断山区报道率为40.54%。收集整理中国地区的染色体核型数据并通过NCBI下载叶绿体基因来构建中国地区鼠尾草属植物的分子进化树,并对横断山区的6种8居群鼠尾草属植物进行细胞学实验获得核型数据,从细胞学水平探讨鼠尾草属下分类关系,补充鼠尾草属的染色体数据。本研究西藏鼠尾草核型为首次报道,核型公式为2n=2x=16=2m(2sat)+12sm+2st。雪山鼠尾草在之前的研究中存在二倍体和四倍体两种倍性的个体(Chen et al., 2018),本研究中采自云南德钦的居群为二倍体,其余4种核型结果与前人研究一致,甘西鼠尾草为四倍体物种,黄花鼠尾草、毛地黄鼠尾草、粘毛鼠尾草为二倍体物种,且6种鼠尾草属植物染色体基数均为x=8,为属内出现频率最高的基数类型,染色体数目也为最普遍的16条与32条(Hu et al., 2016)。另外,通过统计染色体倍性,我们发现鼠尾草属中海拔相对较高的横断山区的多倍化比例与中国地区的多倍化比例基本持平,这说明多倍化可能并不是植物适应高山环境的唯一机制(聂泽龙等, 2004; 杨扬和孙航, 2006)。对于染色体倍性随着统计范围的减小,二倍体比例逐渐降低,推测是由于数据统计量级上巨大差异所造成。
本研究构建的鼠尾草属分子进化树建树结果与Hu等(2018)的研究结果相似,可以较为清晰地在分子进化树分支上划分出4个亚属,但各个亚属会有个别物种掺杂在其他亚属的分支中,特别是荔枝草亚属下物种的分支关系还有待进一步研究。弧隔鼠尾草亚属的分支最为清晰且分为两支,但此分子系统分类学下的亚属关系与传统分类学关系不一致。本研究中6种鼠尾草属植物除粘毛鼠尾草为一年生亚组,其余5种均为多年生亚组,分散于4个不同的支系,分别为隶属于短冠鼠尾草系的雪山鼠尾草、栗色鼠尾草系的黄花鼠尾草、毛地黄鼠尾草系的毛地黄鼠尾草和甘西鼠尾草、西藏鼠尾草系的向鼠尾草。分子系统构建表明粘毛鼠尾草、黄花鼠尾草、毛地黄鼠尾草以及雪山鼠尾草聚为一支,但雪山鼠尾草亲缘关系相对较远;甘西鼠尾草与西藏鼠尾草聚为一支。粘毛鼠尾草与多年生亚组的3种鼠尾草属植物(黄花鼠尾草、毛地黄鼠尾草、雪山鼠尾草)较甘西鼠尾草和西藏鼠尾草亲缘关系较近,而同属一系的毛地黄鼠尾草与甘西鼠尾草却亲缘关系较远,推测可能与甘西鼠尾草染色体组加倍有关。Chen等(2018)研究发现雪山鼠尾草既有二倍体个体又有四倍体个体,但本研究中雪山鼠尾草为二倍体个体,从分子系统树上看,无论是基部类群还是末类群,鼠尾草属染色体大都是二倍体(2n=2x=16),主要是在二倍体水平上的进化,且在染色体倍性的统计中,二倍体物种具有绝对的比例优势,徐波等(2020a, b)的研究也表明在石竹科(Caryophyllaceae)中有多种高山植物维持二倍体进化,并推测二倍体水平上的染色体结构和核型进化是青藏高原地区物种进化的一个重要机制,因此本研究认为多倍化不是鼠尾草属的主要进化途径。
本研究丰富了横断山区鼠尾草属物种的染色体核型数据,验证了多倍化可能并不是植物适应高山环境的唯一机制,论证了多倍化可能不是鼠尾草属的主要进化途径,构建的进化树支持前人分类学结果,分子系统树与染色体数据的结合分析为今后深入研究该属物种的核型进化提供了探索,为开展祖先物种染色体基数推演分析补充了基础数据,但横断山地区的鼠尾草染色体数据仍旧不充分,这可能会造成统计学意义较差,因此染色体数据的补充仍旧十分重要。
