ALKBH5 在肾透明细胞癌中的表达水平及临床意义
2023-11-06朴松哲蔡仙国柯莽郑兰周高波陈超前
朴松哲 蔡仙国 柯莽 郑兰 周高波 陈超前
肾癌是泌尿系统常见的恶性肿瘤之一。肾透明细胞癌(kidney renal clear cell carcinoma,KIRC)是肾癌最常见的病理类型,约占80%,细胞组织形态学特征表现为细胞内脂滴积聚。N6-甲基腺嘌呤(N6-methyladenosine,m6A)修饰是近年来肿瘤领域研究的热点,研究表明m6A 修饰是真核细胞中丰度最高的RNA 修饰方式,参与基因表达调控中的多个环节,如RNA 剪切、成熟、出核、翻译及降解,并影响多种生理、病理进程,如胚胎发育、代谢稳态维持和肿瘤发生、发展。越来越多的研究显示m6A 修饰在肾癌,尤其是KIRC 演进过程中发挥重要作用。Wang 等[1]发现在KIRC 中抑制甲基转移酶(methyltransferase,METTL)14 基因表达可以诱导肿瘤生长,且METTL14 mRNA 高表达患者总生存期(overall survival,OS)较低表达患者明显延长。Li等[2]发现METTL13 基因和蛋白在KIRC 组织中呈低水平表达,敲低METTL3 可明显促进肾癌细胞增殖、迁移和侵袭功能,并诱导肾癌细胞发生G0/G1细胞周期停滞。然而烷烃羟化酶同源5 基因(alpha-ketoglutarate-dependent dioxygenase homolog 5,ALKBH5)在KIRC 中的相关研究较少。最新研究发现,ALKBH5 作为m6A 去甲基化酶与肾癌代谢重编程和肿瘤细胞线粒体含量调节有关[3-4]。本研究利用生物信息学探讨ALKBH5 mRNA 和蛋白在KIRC 组织中的表达情况,分析ALKBH5 与KIRC 患者临床病理特征及预后的关系,以期为寻找KIRC 新的治疗靶点提供理论基础和依据。
1 材料和方法
1.1 数据获取 本研究基于肿瘤基因组图谱(the cancer genome atlas,TCGA)数据库与基因型和基因表达量关联数据库(genotype-tissue expression,GTEx)数据库。登陆UCSC Xena 数据库(http://www.xebabrowser.net),下载经Toil 流程[5]统一处理的KIRC(531 例KIRC组织和72 例癌旁组织)、肾乳头状细胞癌(kidney papillary renal cell carcinoma,KIRP)(291 例KIRP 组织和32例癌旁组织)及嫌色细胞癌(kidney chromophobe renal carcinoma,KICH)(66 例KICH 组织和25 例癌旁组织)项目的转录组测序(RNAseq)数据、患者临床病理参数和OS 资料;登陆GTEx 数据库,下载28 例正常肾组织RNAseq 数据。
1.2 表达差异分析 利用R 软件(V4.0.0)“limma”和“ggplot2”程序包比较KIRC 组织和癌旁组织中ALKBH5 mRNA 表达水平。应用阿拉巴马大学伯明翰分校癌症数据分析数据库(the University of Alabama at Birmingham cancer data analysis portal,UALCAN)(http://ualcan.path.uab.edu/ index.html)分析KIRC(110 例)和癌旁组织(84 例)中ALKBH5 蛋白的表达水平。
1.3 临床信息分组 根据TCGA 数据库中KIRC 患者临床资料对患者分组,去除无意义、缺失或重复的数据,根据年龄分为≤60 岁(271 例)和>60 岁(275 例);根据性别分为男(355 例)、女(191 例);根据种族分为白人(474 例)和有色人种(亚裔+非洲裔,65 例);根据美国癌症分期联合委员会(American joint committee on cancer,AJCC)2007 分期分为Ⅰ~Ⅳ期,其中Ⅰ期276例,Ⅱ期59 例,Ⅲ期125 例,Ⅳ期83 例;根据组织学分级分为G1~G4级,其中G1级14 例,G2级238 例,G3级208例,G4级78 例。登陆UALCAN 数据库,根据KIRC 分子亚型分为A 亚型(205 例)和B 亚型(175 例)[6]。酵母交换型转换/蔗糖不发酵(yeast switch in mating type/sucrose non fermentation,SWI/SNF)是一种ATP 依赖的染色质重塑复合物,SWI/SNF 染色质重塑复合物缺陷被认为与肾癌的发生、发展密切相关[7-9]。根据SWI/SNF染色质重塑复合物是否突变分为突变组(98 例)和未突变组(84例),并比较两组ALKBH5蛋白的表达水平。
1.4 ALKBH5 mRNA 表达水平对KIRC 患者预后的影响 从UALCAN 数据库获取KIRC 患者信息,去除正常组织、无临床信息及重复的数据,根据ALKBH5 mRNA 中位表达水平分为高、低表达组;按性别将KIRC 患者分为不同亚组(男性高表达组和男性低表达组、女性高表达组和女性低表达组),对不同亚组进行生存分析,采用log-rank 检验比较不同组间的生存率。
1.5 基因文库富集分析(gene set enrichment analysis,GSEA) 使用LinkedOmics 数据库(http://www.linkedomics.org/login.php)的LinkFinder 模块分析TCGA 数据库KIRC 队列(479 例)中与ALKBH5 相关的差异表达基因。结果采用Spearman 秩相关分析。使用Linked-Omics 的Link-Interpreter 模块对差异表达基因进行通路和网络分析。采用GSEA 对LinkFinder 结果数据进行基因功能富集分析,显著富集的阈值为错误发现率(false discovery rate,FDR)<0.25 且P<0.05。
1.6 统计学处理 使用R(4.0.3)软件,分别使用ggplot2(3.3.6)、stats(4.2.1)、car、pROC(1.18.0)、survival用于统计和图形分析。非正态分布的计量资料采用Mann-WhitneyU检验或Kruskal-WallisH检验进行组间比较,绘制ROC 曲线评估ALKBH5 mRNA 表达水平对KIRC、KIRP 和KICH 的诊断价值;采用Kaplan-Meier法及log-rank 检验生存曲线并评估患者总生存率。P<0.05 为差异有统计学意义。
2 结果
2.1 ALKBH5 mRNA 和蛋白在KIRC 组织和癌旁组织中表达的比较 与癌旁组织比较,KIRC 组织中ALKBH5 mRNA 显著高表达(P<0.01),见图1A。与癌旁组织比较,KIRC 组织中ALKBH5 蛋白也显著高表达(P<0.01),见图1B。

图1 ALKBH5 mRNA 和蛋白在KIRC 组织和癌旁组织中表达的比较(A:基于FPKM 的ALKBH5 mRNA 表达水平比较;B:基于CPTAC z 值的ALKBH5 蛋白表达水平比较)
2.2 不同临床病理特征KIRC 患者KIRC 组织中ALKBH5 mRNA 和蛋白表达的比较 不同年龄、种族、病理分期、组织学分级的KIRC 患者KIRC 组织中ALKBH5 mRNA 表达水平比较,差异均无统计学意义(均P>0.05)。相比男性KIRC 患者,女性KIRC 患者KIRC组织中ALKBH5 mRNA 表达水平显著升高(P<0.05)。相比B 亚型KIRC 患者,A 亚型KIRC 患者KIRC 组织中ALKBH5 mRNA 表达水平显著升高(P<0.01)。相比SWI/SNF 染色质重塑复合物未突变组KRIC 患者,SWI/SNF 复合物突变组患者ALKBH5 蛋白表达水平显著升高(P<0.01),见图2。

图2 不同临床病理特征KIRC 患者KIRC 组织中ALKBH5 mRNA 和蛋白表达的比较(A:年龄;B:种族;C:病理分期;D:性别;E:组织学分级;F:分子亚型;G:SWI/SNF 染色质重塑复合物突变情况)
2.3 ALKBH5 mRNA 表达水平对不同类型肾癌的诊断价值 ROC 曲线分析显示,ALKBH5 mRNA 表达水平诊断KIRC 的AUC 为0.813(95%CI:0.775~0.850),最佳截断值为5.583,灵敏度为0.667,特异度为0.900;诊断KIRP 的AUC 为0.605(95%CI:0.530~0.680),最佳截断值为6.110,灵敏度为0.218,特异度为0.950;诊断KICH的AUC 为0.707(95%CI:0.607~0.807),最佳截断值为5.086,灵敏度为0.818,特异度为0.604。ALKBH5 mRNA 表达水平对KIRC 的诊断价值最大,见图3。

图3 ALKBH5 mRNA 表达水平诊断不同类型肾癌的ROC 曲线
2.4 ALKBH5 mRNA 表达水平与KIRC 患者预后的关系 Kaplan-Meier 生存曲线表明,ALKBH5 mRNA 高表达组患者总生存率高于低表达组(P<0.01)。亚组分析显示,男性KIRC 患者ALKBH5 mRNA 表达水平对预后无显著影响(P>0.05);但对女性KIRC 患者,与ALKBH5 mRNA 低表达组比较,ALKBH5 mRNA 高表达组总生存率显著升高(P<0.05),见图4。
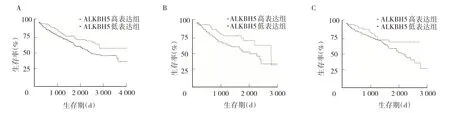
图4 ALKBH5 mRNA 表达水平与KIRC 患者预后的关系(A:KIRC 患者总生存率分析;B:女性亚组总生存率分析;C:男性亚组总生存率分析)
2.5 ALKBH5 在KIRC 中可能参与的信号通路GSEA 富集分析结果显示,ALKBH5 mRNA 在糖酵解通路中的标准化富集分数为2.30,FDR<0.01,P<0.01;在果糖和甘露糖代谢通路中的标准化富集分数为2.27,FDR<0.01,P<0.01;在氨基酸生物合成通路中的标准化富集分数为2.18,FDR<0.01,P<0.01,见图5。

图5 ALKBH5 在KIRC 中可能参与的信号通路(A:糖酵解;B:果糖和甘露糖代谢;C:氨基酸生物合成)
3 讨论
ALKBH5 基因位于染色体17p11.2,其编码蛋白含有394 个氨基酸残基,分子量约43 kD,属于非血红素铁(Ⅱ)/双加氧酶的α-烷烃羟化酶(alpha-ketoglutarate-dependent dioxygenase,ALKB)家族,是高度保守的mRNA 结合蛋白。至今为止在哺乳动物体内共发现9个ALKB 家族成员,分别为ALKBH1~8 和fat mass-and obesity-associated(FTO),其中ALKBH5 和FTO 同为m6A 去甲基化酶。不同于FTO,ALKBH5 对N6,2-O-二甲基腺苷(N6,2-O-dimethyladenosine,m6Am)无活性,可直接催化m6A 甲基化腺苷去除甲基,从而降低靶基因的m6A 水平,调控基因表达参与细胞生命活动及功能调节,且不产生中间产物。目前研究认为ALKBH5 在肿瘤细胞中既可发挥促癌作用亦可发挥抑癌作用。Chao 等[10]研究发现ALKBH5 在肺腺癌组织中高表达,通过人叉头框蛋白M1(forkhead box M1,FOXM1)mRNA 翻译效率来提高肿瘤细胞的增殖和侵袭能力。在乳腺癌中,低氧刺激可上调乳腺癌细胞内ALKBH5 mRNA 水平,介导肿瘤微环境中乳腺癌干细胞的富集[11]。相反,ALKBH5 在胰腺癌中低表达,能够提高胰腺导管腺癌细胞对吉西他滨的敏感性,抑制肿瘤细胞的增殖和迁移[12]。但目前ALKBH5 在肾癌中的研究相对较少。
本研究基于TCGA 数据库探索ALKBH5 基因在KIRC 中的表达情况,结果发现KIRC 组织中ALKBH5 mRNA 和蛋白表达水平均高于癌旁组织,与以往研究结果一致[3,13]。Guimarães-Teixeira 等[13]研究发现ALKBH5 mRNA 在KIRC 中的表达水平高于KIRP 和KICH。本研究发现,ALKBH5 诊断KIRC 的AUC 为0.813,高于KIRP(AUC=0.633)和KICH(AUC=0.707),说明ALKBH5 mRNA 表达水平可较好区分KIRC 和其他类型肾细胞癌,有望成为KIRC 诊断的生物标志物。结合KIRC 患者的临床资料进行生存分析,结果发现女性KIRC 患者肿瘤组织中ALKBH5 mRNA 和蛋白表达水平均高于男性患者,提示ALKBH5 基因和蛋白表达水平可能与性别、性激素水平等因素有关。值得注意的是,KIRC 具有显著的异质性。Brooks 等[6]基于34 个基因将KIRC 分成低风险(A 型)和高风险(B 型)两种分子亚型。本研究发现,A 型KIRC 组织ALKBH5 mRNA 表达水平显著高于B 型KIRC 组织,且生存分析发现ALKBH5 mRNA 低表达的KIRC 患者预后不良,与Brooks 等[6]的研究结果一致。SWI/SNF染色质重塑复合物被证明与KIRC 进展关系密切[14-15]。本研究中SWI/SNF 染色质重塑复合物突变组ALKBH5 蛋白表达水平高于SWI/SNF 复合物未突变组。人类SWI/SNF 染色质重塑复合体由15 个组分组成,包括1 个催化ATP 酶亚基[SWI/SNF related,matrix associated,actin dependent regulator of chromatin,subfamily A,member 4,(SMARCA4)或SMARCA2]和其他3 个 核 心 亚 基(SMARCB1、SMARCC1 和SMARCC2)[16]。双植物同源结构域(plant homeodomain,PHD)锌指家族3(double PHD fingers 3,DPF3)属于D4 蛋白家族,是SWI/SNF 复合体的非催化亚基。DPF3 表达DPF3a 和DPF3b 两种剪接变体。Cui等[17]通过体内及体外实验证实DPF3a 可促进肾癌细胞的迁移,这与临床观察到的DPF3a 在KIRC 转移患者中显著上调的结果一致[18]。在机制上,过表达DPF3a 可上调TGF-β 信号通路相关的基因表达[17],而TGF-β 是上皮细胞-间充质转化(epithelial-mesenchymal transition,EMT)表型的主要诱导因子,是促进癌症转移的信号通路。有研究发现在非小细胞癌(non-small-cell lung cancer,NSCLC)中,过表达ALKBH5 可抑制TGF-β诱导的EMT,并增强NSCLC 细胞的侵袭能力[19]。然而在KIRC 细胞系769-P 中敲低ALKBH5 可下调间质化分子标志物蛋白波形蛋白(Vimentin)的表达水平[20]。推测ALKBH5 在不同组织中、在肿瘤细胞不同的恶性条件下、不同的细胞内外通路中对EMT 的调控作用是有差别的。此外,Colli 等[21]和Protze 等[18]发现位于DPF3 第1 个内含子内的风险位点(rs4903064)可产生一种缺氧反应增强子,其活性受Von Hippel-Lindau(VHL)肿瘤抑制基因调节,从而影响体外KIRC 细胞的增殖能力。在KIRC 患者的VHL 突变较为常见。DPF3 在KIRC 患者中显著上调,在存在VHL 突变的患者中尤为明显。VHL 的缺失导致缺氧诱导因子-1α(hypoxia-inducible factor-1α,HIF-1α)蛋白水平上调,进而激活参与增殖、凋亡和转移的基因的转录。值得注意的是,最新研究发现ALKBH5 启动子区存在HIF-1α 的结合位点,其核心序列为5'-RCGTG-3',被称为缺氧反应元件chypoxia-responsive element,HRE),分别位于-486/-482(HRE A)和+265/+269(HRE B)。染色质免疫沉淀实验和荧光素酶实验均证实ALKBH5 HRE A 与HIF-1α 结合,未与HIF-2α 结合[22]。低氧条件可诱导肾癌细胞中的ALKBH5 基因表达水平上调,且该过程是HIF-1α 依赖性的。因此,本研究推测在KIRC 中SWI/SNF 复合物突变(DPF3 基因rs4903064 位点)产生缺氧反应增强子,通过HIF-1α 上调ALKBH5的表达,进而促进EMT 过程。
ALKBH5 mRNA 和蛋白高表达可能在KIRC 的演进中发挥重要作用。本研究生存分析显示,ALKBH5 mRNA 高表达的KIRC 患者总生存率高于低表达者。Guimarães-Teixeira 等[13]研究结果显示ALKBH5 mRNA表达水平与患者预后无关。Zhang 等[3]研究发现ALKBH5 高表达与KIRC 肿瘤体积较大、高恶性度、预后较差相关。上诉结果与本研究分析结果不同,可能与样本量及研究方法不同有关。本研究进一步按性别进行亚组分析,结果显示在女性KIRC 患者中,与ALKBH5 低表达组比较,ALKBH5 高表达组患者总生存率显著增高;然而,在男性KIRC 患者中,ALKBH5 mRNA表达水平对预后无显著影响。提示ALKBH5 mRNA 表达水平可成为女性KIRC 患者判断预后的有效标志物,但具体机制尚不清楚。
本研究的GSEA 富集分析结果显示ALKBH5 mRNA 可能参与糖酵解、果糖和甘露糖代谢、氨基酸生物合成等信号通路。研究发现ALKBH5 启动子区-486/-482 存在HIF-1α 的结合位点,在多种细胞系中,DNA 损伤和缺氧环境可以激活ALKBH 5 基因的转录[23]。以上结果说明ALKBH5 可能通过调控KIRC 的糖酵解过程在肾癌的发生、发展及转移等过程中发挥重要作用。
综上可知,ALKBH5 作为促瘤因子在KIRC 发生、发展、转移及预后评估过程中发挥重要作用,推测ALKNH5 有成为KIRC 诊断标志物和治疗靶点的潜在可能,同时为临床评估患者预后提供理论依据。
