不同造林密度马尾松人工林分化特征及其对生产力的影响*
2023-11-05颜培栋杨章旗黄绥理周永斌零天旺
颜培栋 李 鹏 杨章旗 黄绥理 周永斌 零天旺
(1. 广西壮族自治区林业科学研究院 国家林业和草原局马尾松工程技术研究中心 广西马尾松工程技术研究中心 广西优良用材林资源培育重点实验室 南宁 530002;2. 广西横州市镇龙林场 横州 530327)
林木个体大小分化对森林生态系统的空间结构、群落演替、种间(内)关系以及林分结构和生产力水平均具有显著影响(Ali,2019;Levicket al.,2015),尤其在人工纯林生态系统中,如何减少林分内个体分化大小(Tschiederet al.,2012),对培育中大径材林分(谭健晖,2021)、提高林分生产力水平尤为重要。林木个体大小分化在空间结构上受光照、土壤水分和无机养分等供给能力的影响,林木对生长资源的获取和利用效率存在差别,进而发生生态位分化引起个体大小差异(Boydenet al.,2008;Schumeet al.,2004)。在时间上,林分生长早期阶段种间和种内竞争较弱(Oheimbet al.,2011),随着林龄增加,林木树冠开始互相接触遮蔽,根系依靠自身趋水和趋肥特性(Zhanget al.,2006),向相邻个体水肥条件较好的地方延伸,个体间竞争加剧,该过程中处于优势地位的个体占有更多生长资源,相比劣势个体更容易在林分中脱颖而出(Binkleyet al.,2010),从而导致林分内个体大小不断分化。当林分内个体竞争达到一定水平时,林分会发生自然稀疏(Duanet al.,2019),劣势被压木枯死,为优势木提供生长空间和养分,尤其是高密度种植时,因获取资源的非对称性竞争发生年龄更早(Stankovaet al.,2017;谌红辉等,2004)。林木个体大小分化直接导致林木径阶结构改变,林木径阶结构分布特征指林木个体按径阶大小的分布状况,是反映林分结构的重要指标(Yanget al.,2002),在人工林纯林发育过程中,探讨林木个体大小分化及其径阶分布特征对提高林分生产力和培育中大径材林木具有重要意义。
马尾松(Pinus massoniana)是我国亚热带地区特有的乡土树种和南方荒山造林先锋树种,也是重要的工业原料林树种,具有速生、丰产、适生性强、用途广等优点(孟祥江等,2018)。目前,关于马尾松人工林经营技术方面已开展施肥效应(胡炳堂等,2000)和密度调控(王建军等,2019)等研究,并取得了显著成果;然而在马尾松人工林经营过程中仍存在密集种植、采伐过早等问题,既不利于大径材培育,也造成林分生产力的浪费(李婷婷等,2015)。如何改进经营方法,培育中大径材马尾松人工林,提高林分生产力,尤其是在充分了解不同造林密度马尾松人工林生长发育过程的基础上,对马尾松人工林个体分化和径阶分布特征及其对林分生产力影响的探讨尤为重要。鉴于此,本研究基于南亚热带地区不同造林密度样地长期监测结果,分析4 种林分密度马尾松人工林个体大小分化、径阶分布特征及其对林分生产力的影响,阐明林龄、造林密度和自然稀疏过程对林木个体大小分化和生产力的影响规律,以期为确定适宜经营密度、培育马尾松林中大径材、提高林分生产力提供依据。
1 试验地概况与研究方法
1.1 试验地概况
试验林位于镇龙林场那歪分场4 林班10 小班,地处广西横州市境内(109°08′—109°19′E,23°02′—23°08′N),属南亚热带季风气候区,降雨充沛,年均降雨量1 477 mm,年均蒸发量1 056 mm,年均温度21.5 ℃,极端低温-1 ℃,极端高温39.2 ℃,年均日照时数1 758.9 h,无霜期336 天。试验地海拔450 m,坡度25°,坡向西南216°,中上坡位;土壤为紫色砂页岩发育而成的赤红壤,平均土层厚60 cm;林下灌草主要有山鸡椒(Litsea cubeba)、华南毛柃(Eurya ciliata)、桃金娘(Rhodomyrtus tomentosa)、山乌桕(Sapium discolor)、三叉苦(Euodia lepta)、鸭脚木(Schefflera octophylla)、黄毛榕(Ficus esquiroliana)、 大青(Clerodendrum cyrtophyllum)、棕叶芦(Thysanolaena maxima)、乌毛蕨(Blechnum orientale)和五节芒(Miscanthus floridulu)等。
1.2 样地设置与调查
试验林于1997 年5 月采用马尾松优良家系(桂MVF443)半年生营养杯苗(平均苗高25 cm、地径0.26 cm)造林,随机区组设计,设置3 个区组(即3 次重复),2 500(A)、3 333(B)、4 500(C)、6 000 株·hm-2(D)4 种造林密度,株行距分别为2 m×2 m、1.5 m×2 m、1.5 m×1.5 m、1 m×1.67 m。每个小区面积30 m×30 m,造林后前2 年进行常规割草抚育,其后采用近自然经营措施。每种密度选择坡度、坡向、土壤等立地因子基本一致的3 块20 m×20 m 标准地(样地间隔>50 m),共布设12 块固定样地(范志伟等,2012)。每年测定林木树高(tree height,H)和胸径(diameter at breast height,DBH)等生长指标,并记录林木存活状况。2006—2020 年连续15 年,每年年底进行定期测定,样地基本概况如表1 所示。
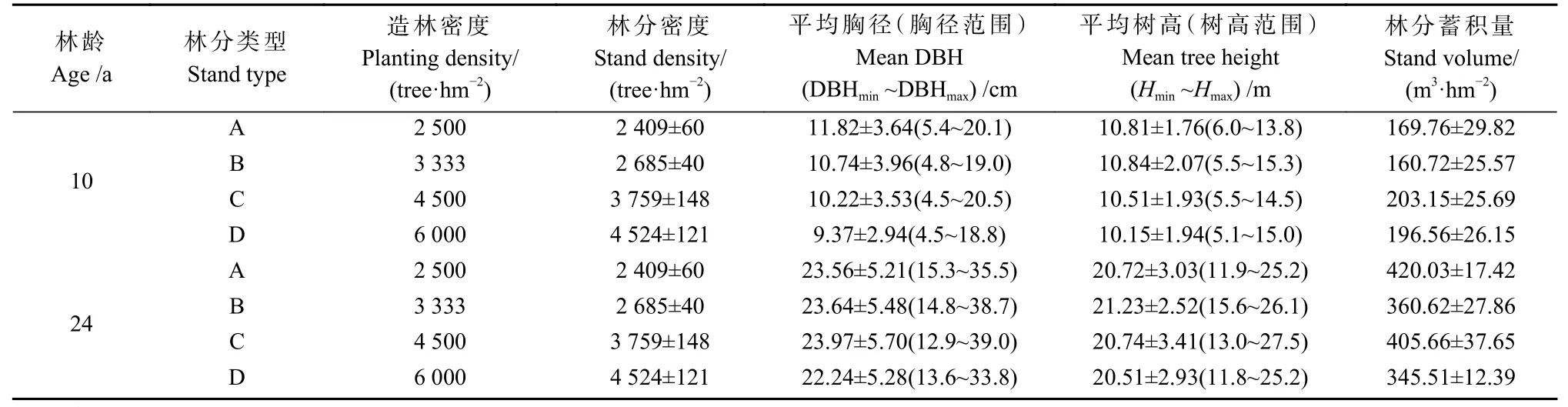
表1 样地基本概况①Tab. 1 Basic characteristics of the sampling plots
1.3 数据分析与计算方法
1.3.1 林分生产力计算 根据林分每木检尺结果,马尾松人工林活立木单株材积采用二元材积公式V=7.142 654 37×10-4×DBH1.867008H0.9014632计算,其中:V为单株材积,m3;DBH 为胸径,cm;H为树高,m。单位面积活立木总材积为活立木单株材积累加除以样地面积。林分生产力采用林分活立木总材积的平均年生长量(stock volume annual increment,VAI,m3·hm-2a-1)表示。
1.3.2 林木个体分化程度表征 参考杨桂娟等(2019)方法,采用基尼(Gini)系数表征林分内林木个体大小分化程度。Gini 系数最早由Lorenz 在1905 年提出(Lorenz,1905),用于社会科学中分析人群收入的差异程度,后被引入表征植物种群内个体大小的差异程度,并被广泛应用。本研究中Gini 系数采用分组计算法,其计算公式为:
式中:n为样地内活立木株数;Yi为活立木单株断面积。
1.3.3 林分自然稀疏效应 林分自然稀疏程度(selfthinning rate)采用林木当年死亡数量除以总株数乘以100%表示。为确定林分自然稀疏过程中死亡单株对林分整体水平的影响,采用每年林分自然稀疏的个体平均断面积与林分死亡发生前林分整体平均断面积(mean basal area,BA)的比值(BAdeath: BAtotal)反映,BAdeath: BAtotal>1 表示死亡单株平均大小大于林分整体水平,BAdeath: BAtotal<1 表示死亡单株平均大小小于林分整体水平。
1.3.4 径阶分布划分 按径阶将马尾松分为大径材(胸径在26 cm 径阶及以上)、中径材(胸径在18~24 cm径阶之间)和小径材(胸径在16 cm 径阶及以下),以2 cm 为1 个径阶。
采用Excel 2016 和SPSS 20.0 软件对数据进行统计分析,所用数据均以均值±标准差表示。运用重复测量方差分析(repeated measure ANOVE)检验造林密度和林龄对马尾松人工林个体大小分化、径阶分布特征和林分生产力影响的差异,差异显著性多重比较采用Tukey’s 检验(n=3),显著性水平α=0.05。利用R 语言的Random Forest 程序包进行随机森林分析,判定林龄、造林密度和自然稀疏过程对林木个体大小分化和林分生产力的影响。应用Smart PLS 软件构建偏最小二乘法-结构方程进行Gini 系数和林分生产力的影响路径分析。
2 结果与分析
2.1 马尾松人工林个体大小分化特征
Gini 系数随林龄增加显著减小(F=20.598,P<0.001),个体间分化程度随造林密度增加显著增大(F=6.172,P<0.05),林龄与造林密度的交互作用对Gini 系数无显著影响(图1)。不同造林密度下Gini 系数与林龄间呈极显著线性负相关(P<0.001)。
由图2 可知,林分自然稀疏过程中,随林龄增加活立木株数显著减少(F=324.168,P<0.001),由造林时的2 500~6 000 株·hm-2降至10 年生时的2 344~4 790 株·hm-2,并逐渐趋于一致,到24 年生时为588~1 396 株·hm-2。造林密度越大,林分内存活株数减小越快,减少数量越多,随自然稀疏进程持续趋于一致,到17 年生后不同造林密度活立木株数间无显著差异,林龄与造林密度的交互作用对活立木株数具有显著影响(F=8.596,P<0.001)。不同造林密度下活立木株数与林龄呈极显著的指数函数变化趋势(P<0.001)。随林龄增加林分自然稀疏率存在显著差异(F=4.389,P<0.01),造林后头9 年,A(2 500 株·hm-2)密度自然稀疏率最小为3.65%,其次为C(4 500 株·hm-2)密度(16.20%),而B(3 333 株·hm-2)和D 密度(6 000 株·hm-2)自然稀疏率较大,分别为19.44%和24.60%,从整体来看,林分自然稀疏率较大的林龄为13~14 年和20 年生时,表明在这2 个时间点林分内个体竞争程度较大,竞争剧烈。不同造林密度间林分自然稀疏率存在显著差异,高密度自然稀疏率大于低密度,而林龄与造林密度的交互作用对林分自然稀疏率无显著影响。
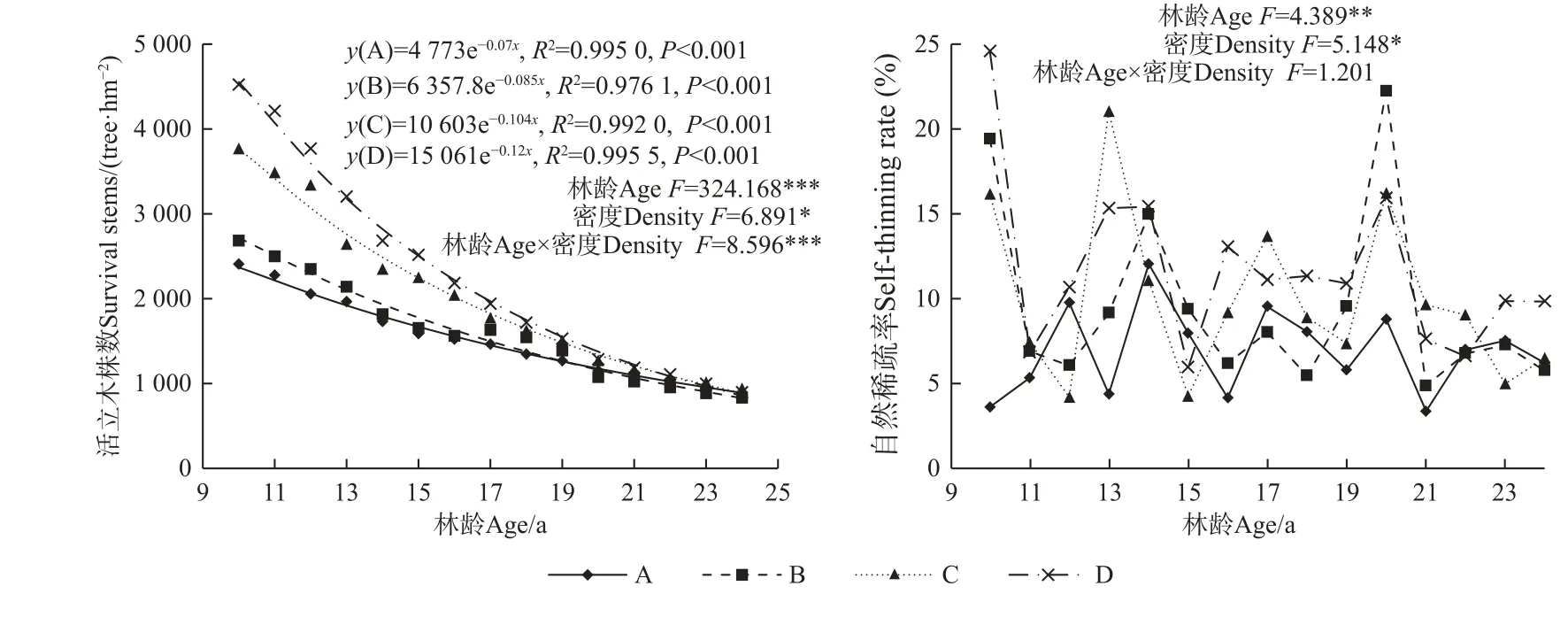
图2 马尾松人工林活立木数量和自然稀疏率的变化趋势Fig. 2 Dynamics of surviving stems and self-thinning rate of P. massoniana plantations
活立木株数与Gini 系数间呈对数函数分布(P<0.001),且随活立木株数增加Gini 系数增大,并逐渐趋于水平,而自然稀疏率与Gini 系数间无显著相关(图3)。
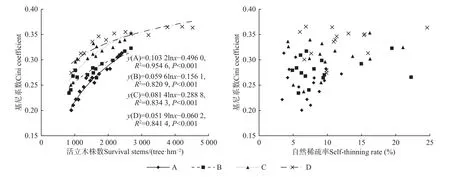
图3 马尾松人工林活立木数量和自然稀疏率与Gini 系数的关系Fig. 3 The relationship between the surviving stems and self-thinning rate and Gini coefficient of P. massoniana plantations
林分自然稀疏过程中,林分死亡个体断面积与林分平均断面积的比值(BAdeath: BAtotal)随林龄增加显著增大(F=2.668,P<0.05)(图4),整体呈上升趋势,且在19 年生后均有出现>1 的情况,表明林分自然稀疏过程中,随林龄增加死亡单株平均大小大于林分整体水平,死亡单株对林分整体的影响随林龄增加而加大。不同造林密度下BAdeath: BAtotal具有显著差异(F=3.018,P<0.01),随造林密度增加BAdeath: BAtotal增大,林龄和造林密度的交互作用对BAdeath: BAtotal无显著影响。不同造林密度下BAdeath: BAtotal与林龄间均呈幂函数分布特征(P<0.01)。
2.2 马尾松人工林径阶分布特征
马尾松人工林大、中和小径材株数随林龄增加均具有显著性差异(P<0.01)(图5),其中大径材株数随林龄增加呈近似“S”形变化,14 年开始出现大径材,14~17 年间大径材株数增加较慢,18~22 年间大径材株数增加最快,23 年后逐渐平稳;22~24 年间不同造林密度大径材株数具有显著差异(P<0.05),均呈现C密度显著高于其他密度。中径材株数随林龄增加呈先增加后减少趋势,均在17~19 年间出现极大值;10~19 年间马尾松人工林中径材株数均在A 密度下最多,表明A 密度适用于中径材马尾松速生丰产林培育,且主伐年龄在19 年后适宜。小径材株数随林龄增加呈显著减少趋势,10~17 年间均呈现D 密度显著高于他密度,而随林龄增加不同造林密度马尾松人工林小径材株数逐渐减小并趋于一致。径阶分布特征三元相图表明,马尾松人工林大径材株数占比在25%及以上包括A 密度在21~24 年、B 和C 密度均在22~24 年、D 密度在24 年;中径材株数占比在50%及以上包括A 密度在17~24 年、B 密度在20~24 年、C 密度在24 年、D 密度在23 年;小径材株数占比在50%以下包括A 密度在17~24 年、B 密度在19~24 年、C 和D密度均在20~24 年。
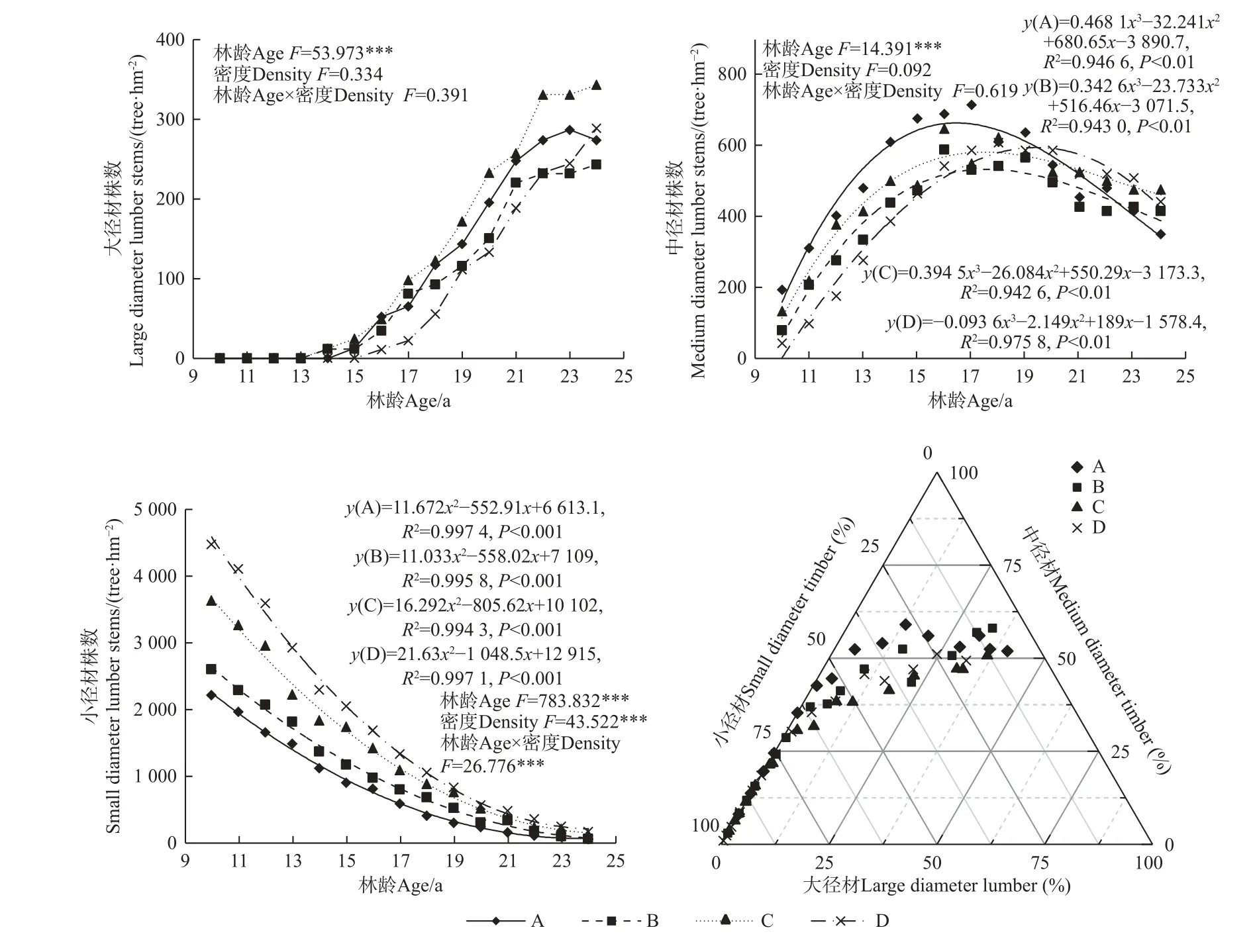
图5 不同造林密度马尾松人工林大、中和小径材株数变化及分布特征Fig. 5 Changes and distribution characteristics of large, medium and small-diameter trees with different densities of P. massoniana plantations
2.3 马尾松人工林林分生产力
重复测量方差分析(图6)表明,马尾松人工林林分蓄积量和林分生产力随林龄变化均存在极显著差异(P<0.001),造林密度、林龄与造林密度的交互作用对林分蓄积量和林分生产力无显著影响。不同造林密度马尾松人工林林分蓄积量随林龄增加呈对数函数增加,其中A 密度下林分蓄积量变化幅度最大,从9 年生的183 m3·hm-2至24 年生的420.03 m3·hm-2,15年间增加237.03 m3·hm-2,其次为C 和B 密度,分别增加202.51 和186.76 m3·hm-2,D 密度下林分蓄积量增加幅度最小,为148.95 m3·hm-2。不同造林密度马尾松人工林林分生产力随林龄增加整体呈先增加后减小并逐渐趋于稳定,其中A 和B 密度在13 年生时最大,分别为19.31 和18.76 m3·hm-2,C 和D 密度均在11 年生时最大,分别为20.48 和19.67 m3·hm-2。不同造林密度林分生产力的大小顺序在不同林龄阶段呈现不同变化趋势,其中10~11 年林分生产力为C > D > A > B,12~15 年林分生产力为C > A > B > D,16~24 年林分生产力为A > C > B > D。
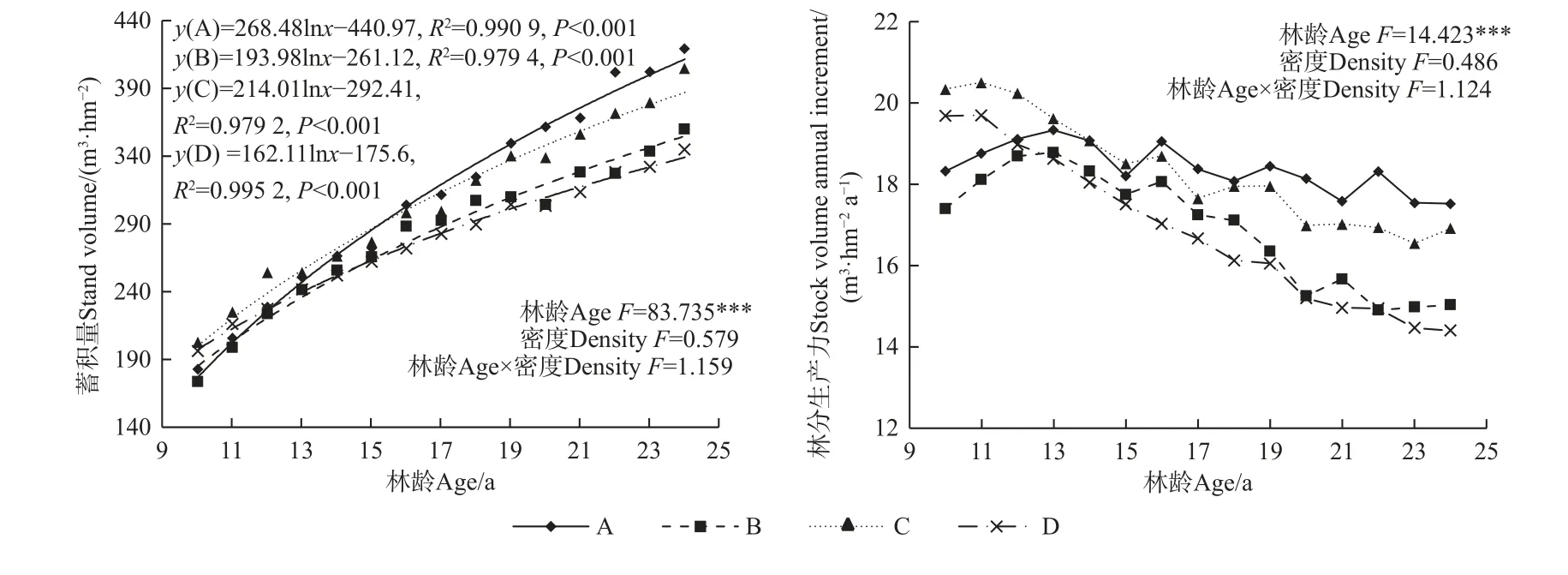
图6 不同造林密度马尾松人工林林分生产力变化Fig. 6 Changes in stock volume and stock volume annual increment with different densities of P. massoniana plantations
马尾松人工林个体分化程度Gini 系数与林分蓄积量呈显著负相关(P<0.001),与林分生产力呈显著正相关(P<0.01),且Gini 系数越大林分蓄积量越小,林分生产力越大(图7)。马尾松人工林活立木株数与林分蓄积量呈显著负相关(P<0.001),A 和B 密度林分生产力随活立木株数增多呈先增加后减小的二次函数关系,均在2 000~2 500 株·hm-2时达到极大值,C 和D密度林分生产力随活立木株数增多呈显著正相关(P<0.001)。马尾松人工林自然稀疏率与林分蓄积量和生产力仅在D 密度下呈显著正和负相关(P<0.001),而A、B、C 密度无显著相关。
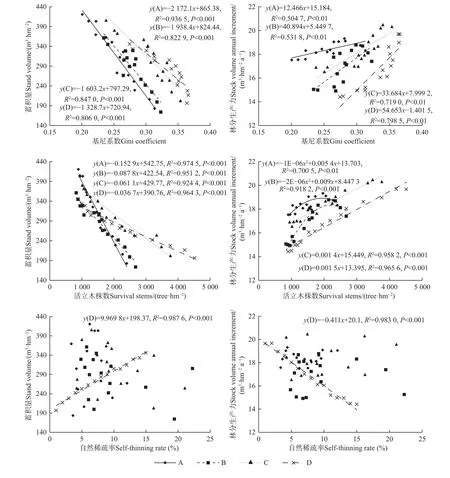
图7 林分生产力与个体大小分化和自然稀疏效应的关系Fig. 7 The relationship between stock volume annual increment and individual size differentiation and self-thinning effect
2.4 林分生产力和Gini 系数的影响效应分析
随机森林分析(图8A)表明,林龄、造林密度和活立木株数对Gini 系数均具有显著影响(P<0.01),自然稀疏率对Gini 系数无显著影响,其中造林密度的影响效应最大(38.41%),其次是活立木株数(28.92%)和林龄(28.15%)。林龄、造林密度、活立木株数和Gini 系数对林分生产力均具有显著影响(P<0.01)(图8B),自然稀疏率对林分生产力无显著影响,其中活立木株数的影响效应最大(32.03%),其次是造林密度(26.25%)、Gini 系数(24.64%)和林龄(23.23%)。
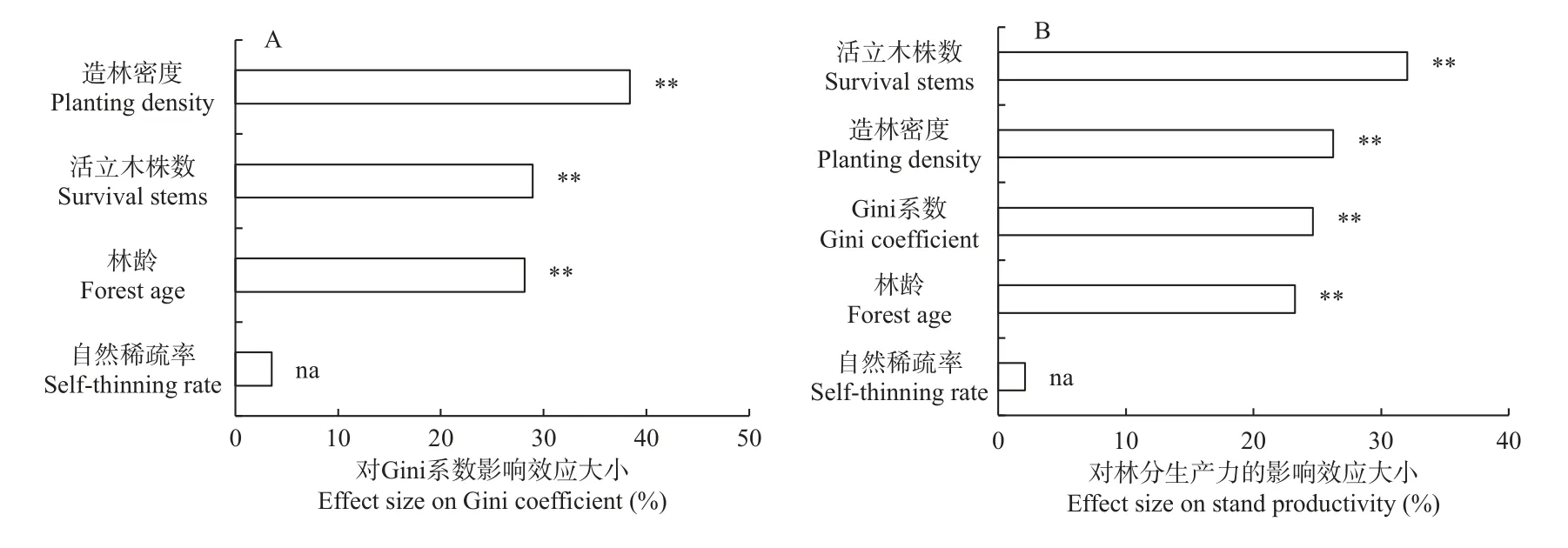
图8 林龄、造林密度和自然稀疏对Gini 系数和林分生产力的影响效应分析Fig. 8 Effect analysis of forest age, planting density and self-thinning on Gini coefficient and stand productivity
结构方程显示(图9),林龄对马尾松人工林活立木株数和Gini 系数均具有显著负向影响(路径系数分别为-0.798 和-0.464),对自然稀疏率和林分生产力具有负向影响(路径系数分别为-0.163 和-0.072),活立木株数对林分生产力具有显著正向影响(路径系数为0.953),表明马尾松人工林林分生产力随林龄的变化主要是通过影响活立木株数变化的间接作用,即随林龄增加经过自然稀疏过程,导致林分活立木株数改变而间接影响林分生产力。造林密度对Gini 系数具有显著正向影响,对自然稀疏率、活立木株数均具有正向影响(路径系数分别为0.252 和0.274),对林分生产力具有负向影响(路径系数为-0.083),Gini 系数对林分生产力具有显著负向影响(路径系数为-0.63),表明造林密度对林分生产力产生的影响主要是通过不同造林密度条件对马尾松人工林林分个体大小分化(Gini 系数)影响,从而间接影响林分生产力。
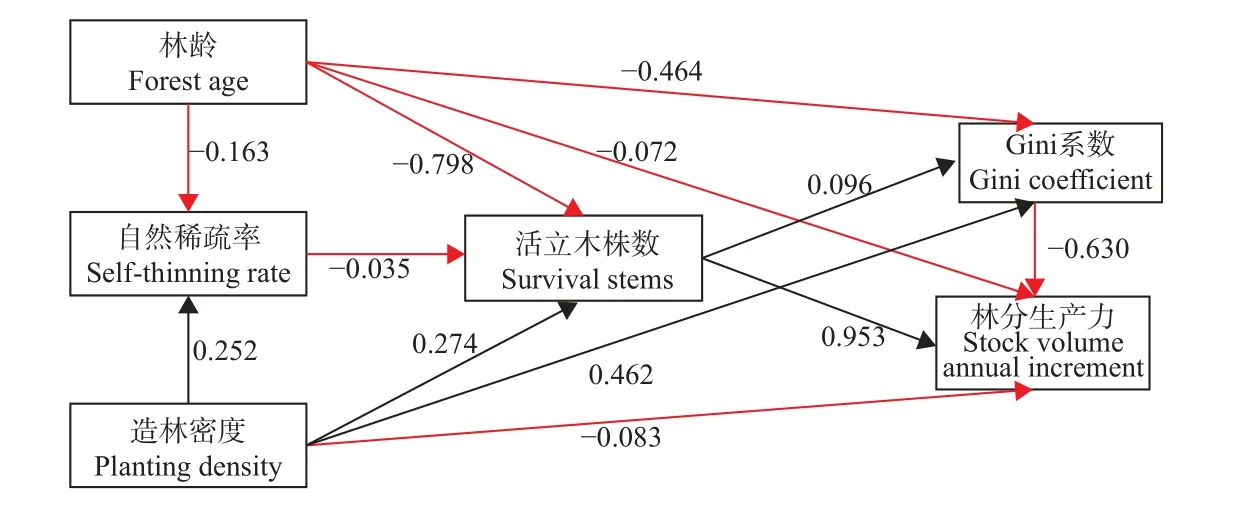
图9 Gini 系数和林分生产力的影响路径分析Fig. 9 Analysis of the impact path of Gini coefficient and stock volume annual increment
3 讨论
林分内个体大小分化反映其对生长资源的获取和利用差异,初始造林密度越大,林分进入竞争阶段时间越早,进而导致林分内个体间分化程度越大(谌红辉等,2011),与本研究马尾松人工林个体间分化程度随造林密度增加显著增大的结果一致,均呈现D>C>B>A,表明A 密度条件具有保持较低林分分化水平。杨桂娟等(2019)对杉木(Cunninghamia lanceolata)人工林个体分化大小研究表明,个体间分化程度与林龄以及是否发生自然稀疏无关,与本研究结果相一致。而马尾松人工林个体大小分化随林龄增加显著减小与杉木研究规律(杨桂娟等,2019)并不一致,其原因一方面是本研究采用的材料为同一家系,具有相对分化程度较小等优良特性,而杉木研究为种子园混系,个体差异较大;另一方面可能是因树种不同,马尾松具有向阳性、不耐阴,随着林龄增加,林分内个体间竞争强度增大,林分优势个体对相邻劣势个体产生冠层“欺压”,根系伸展受限,被压的个体很快发生自然稀疏而枯死(Sunet al., 2011),加之马尾松活枝枝下高较高,林木存留可供光合作用针叶数量较少,林分内劣势被压木大量死亡(马尾松自然稀疏率平均达10%),从而导致个体大小分化随林龄增加而减小,这与本研究林分自然稀疏过程中活立木株数随林龄增加显著减小并逐渐趋于一致、个体大小分化随活立木株数增加而增大的对数函数分布特征结果相一致。此外,本研究还发现林分自然稀疏过程中,林分死亡个体断面积与林分平均断面积比值(BAdeath: BAtotal)随林龄增加显著增大,且在19 年后均有出现>1 的情况,表明随林龄增加林分自然稀疏过程中,死亡单株平均大小大于林分整体水平,死亡单株对林分整体影响随林龄增加而加大。
林木大小径阶分布特征可用于评估林分结构,了解林分不同径阶水平的变化规律,可指导林分中大径材的培育(Zasada,2013)。本研究表明,林龄对马尾松人工林大、中和小径材株数变化均具有显著影响(P<0.01),径阶分布特征中14 年生马尾松人工林开始出现大径材,以A 密度(21 年生和17 年生)出现25%大径材和50%中径材最优,加之A 密度下林分蓄积量变化幅度最大,15 年间增加237.03 m3·hm-2,且16年后林分生产力均处于较大水平,综合林分生长和中、大径材演变的林龄效应来看,A 密度适用于马尾松中大径材人工林培育。此外,本研究发现C 密度是最晚出现小径材低于50%的林分(20 年生),且15 年前均以C 密度下马尾松人工林生产力水平最佳,表明C 密度可作为马尾松短周期工业原料林培育模式,与陈旋等(2020)对马尾松优良家系人工林生长及直径分布的影响结果相一致。随林龄增加林木进行光合作用,吸收利用养分累积生物量,导致林分蓄积量不断增加(Jagodzińskiet al., 2018),与本研究马尾松人工林林分蓄积量随林龄呈对数函数显著增加的结果相一致。不同林龄马尾松人工林林分年均蓄积量(林分生产力)随林龄增加呈现波动性变化,主要是由于林木生长受水、温、干旱等自然环境长期作用的结果(冯源等,2019)以及林木生长大小年对生物量累积的效应,但整体呈先增加后减小并逐渐趋于稳定,主要是由于马尾松人工林生长初期属于快速生长阶段,其快速累积生物量导致林分生产力水平快速增加,林木生长后期由于养分供应和种内竞争加剧,导致林分生产力有所下降并逐渐趋于稳定(吴鹏等,2011),而不同造林密度达到生产力峰值的时间并不一致,其中A 和B 密度在13 年生,C 和D 密度在11 年生,这主要是由于不同造林密度林分种内进入竞争的时间大小不一致,产生生态位林分化时间差异引起的(谌红辉等,2011;丁贵杰,2003),林分密度越大,越早进入竞争阶段,生产力达到峰值时间越短,与本研究结果相似。
随机森林分析表明,林分生产力和Gini 系数均受林龄、造林密度、活立木株数的显著影响,受自然稀疏率的影响并不显著,其中Gini 系数受造林密度的影响效应最大,达38.41%,表明造林密度对林木个体大小分化影响效应较大,选择适宜造林密度减小林木个体分化尤为重要。活立木株数对林分生产力的影响效应最大(32.03%),表明林分生产力主要由在竞争过程中具有优势个体的生长状况决定,发生自然稀疏被淘汰的劣势个体对林分生产力和个体大小分化的影响较小。通过构建结构方程模型进一步明晰林龄、造林密度和自然稀疏过程对林分生产力和Gini 系数的直接或间接影响,可为明确马尾松人工林受林龄、造林密度和自然稀疏的调控机制提供理论依据(Zhuet al., 2019)。结构方程模型显示,林龄对马尾松人工林活立木株数和Gini 系数均具有显著负向影响,造林密度对Gini 系数具有显著正向影响,这是因为随着林龄增加,林分发生自然稀疏效应,活立木株数逐渐减小,优势木所占有生态位加大,林分个体大小分化减小,而林分密度增大必然会引起林木生态位分化增大,个体大小分化增加。林龄对生产力的影响通过“林龄-活立木株数-林分生产力”的间接作用产生,其原因是随林龄增加,林分内活立木株数改变,而林分生产力又由林分内优势个体生长状况决定(Fleminget al.,2014),从而导致林龄与生产力间的间接作用。造林密度对林分生产力的影响通过“造林密度-Gini 系数-林分生产力”的间接作用产生,表明不同造林密度通过调控林分内个体大小分化方式,进而调控林分生产力水平。因此,在生产实践过程中,当林分生态位分化达到一定程度,通过抚育间伐方式,伐除被压木既可以减小个体分化维持林分生产力不下降,又可以作为土壤养分的来源,成为优势木生长的重要养分来源,维持土壤地力。
4 结论
马尾松人工林林木个体分化随林龄增加呈显著线性负相关,随造林密度增加显著增大。林分自然稀疏过程中,活立木株数呈指数函数显著减小,且造林密度越大林分内存活株数减小越快,减少数量越多;林分自然稀疏率较大的林龄为13~14 年和20 年生,表明在这2 个时间点林分内个体竞争程度较大,竞争剧烈;随林龄增加死亡单株对林分整体影响逐渐加大,尤其是19 年生死亡单株平均大小大于林分整体水平。径阶分布特征中以A 密度最早出现25%大径材和50%中径材(21 年生和17 年生),C 密度最晚出现小径材低于50%的林分(20 年生),且15 年间林分蓄积量增加顺序为A(237.03 m3·hm-2)>C(202.51 m3·hm-2)>B(186.76 m3·hm-2)>D(148.95 m3·hm-2),综合林龄效应和径阶分布来看A 密度适宜中大径材马尾松人工林培育。林龄、造林密度、活立木株数对林分个体分化和生产力均具有显著影响,其中造林密度对林分个体的影响效应最大(38.41%),活立木株数对林分生产力的影响效应最大(32.03%),林龄和造林密度对林分生产力影响主要通过“林龄-活立木株数-林分生产力、造林密度-Gini 系数-林分生产力”的间接作用。本研究主要明晰林龄、造林密度和自然稀疏过程对马尾松人工林个体分化特征的影响及其与林分生产力的关系,并通过构建各因素间结构通路为调控个体分化特征、提高林分生产力、确定适宜南亚热带地区马尾松人工林经营密度提供依据。
