木纳格葡萄谷胱甘肽-S-转移酶VvGST1基因克隆与序列分析
2023-09-12伊丽达娜迪力夏提
王 曼,张 政,伊丽达娜·迪力夏提,吴 斌
(1.新疆农业大学食品科学与药学学院,乌鲁木齐 830052;2.新疆农业科学院农产品贮藏加工研究所/新疆农产品加工与保鲜重点实验室,乌鲁木齐 830091)
0 引 言
【研究意义】木纳格(VitisviniferaL.cv Munage)是新疆主栽葡萄品种之一,其采后极易受到病原菌的侵染。随着分子生物学和生物技术的运用,许多抗真菌基因已经从多种植物中分离出来:如几丁质酶、β-1,3-葡聚糖酶和其它致病相关蛋白[1]。在植物与病原菌胁迫响应的过程中,植物进化出一系列复杂的机制来应对病原菌感染,包括病原菌感知、信号转导和诱导特定抗性相关基因[2]。当植物受到病原微生物侵染时主要是通过防御基因的表达、氧化应激反应和代谢系统获得抗性来抵抗病菌的入侵[3]。【前人研究进展】谷胱甘肽-S-转移酶(glutathione-S-transferase,GST)具有多功能性,在植物体内发挥解毒功能[4]。GST对于底物的特异性选择是由两个基本的功能域决定,分别是位于N端的GSH结合位点和位于C端的化合物底物的结合位点[5]。作为GSH途径中的关键酶之一,具有催化还原性GSH和其他物质(如疏水、亲电底物)产生共价化合物的作用[6]。GST是一个多样化的蛋白质家族编码的多基因家族。GSTs可作为替代底物催化各种反应,它们可参与生物解毒、应激响应和次级代谢等生理过程[7]。GST还是一种可溶性蛋白由两个亚基组成,可催化GSH与疏水性物质发生亲电取代反应,转运蛋白可将相关代谢物转运至液泡中,进而降解有毒有害物质,从而减少对细胞的毒害作用[8]。植物生长发育调节和次级代谢产物的运输等过程需要GST家族基因的参与,其作用表现在抵御病原微生物侵染、排除外来化合物、抑制自身代谢产生的有毒物质以及冷害等环境胁迫方面效果显著[9]。【本研究切入点】目前,已对小麦[3]、水稻[10]和烟草[11]等植物中的GST基因进行了生物学分析并发现GST基因在提高植物对生物和非生物胁迫的抗性中发挥着重要的作用[12]。然而,关于GST基因参与调控果实采后抗性的研究较少,尤其是葡萄中VvGST1的分子生物学的研究鲜有报道。需分析木纳格葡萄谷胱甘肽-S-转移酶VvGST1基因克隆与序列分析。【拟解决的关键问题】以木纳格葡萄为试材,对其VvGST1基因克隆并进行生物信息学分析,为研究VvGST1基因在鲜食葡萄中的抗病功能奠定分子基础。
1 材料与方法
1.1 材 料
1.1.1 葡 萄
木纳格葡萄采于新疆阿图什市。挑选果梗新鲜、成熟度一致(TSS≥18%)、大小均一、无表面损伤无病虫害的葡萄串。采收后的葡萄立即用冷链车运回实验室,用液氮速冻果肉后置于-80℃保存备用。
1.1.2 试 剂
琼脂糖、2×EasyTaq®PCR SuperMix (+dye),生工生物工程(上海)股份有限公司;HyPure浆果RNA提取试剂盒,北京君诺德生物技术有限公司;M5 DL2000 DNA Marker,北京聚合美生物科技有限公司;5X All-In-One RT MasterMix (with AccuRT Genomic DNA Removal Kit),ABM公司。
1.1.3 主要仪器设备
台式离心机,上海安亭科学仪器制造厂;PCR仪,美国Bio-Rad有限公司;漩涡混合器,海门市其林贝尔仪器制造有限公司;干式恒温器,杭州美盛仪器有限公司;核酸定量仪,杭州奥盛有限公司;电泳仪电源、电泳槽、凝胶成像系统,上海天能有限公司。
1.2 方 法
1.2.1 总RNA 提取与cDNA第一条链的合成
使用HyPure浆果RNA提取试剂盒,提取葡萄果实的总RNA。随后,通过琼脂糖凝胶电泳检测RNA的完整性。cDNA合成采用FastKing cDNA第一链合成试剂盒。
1.2.2 葡萄果实VvGST1全长克隆
根据GenBank数据库中公布的酿酒葡萄VvGST1基因核苷酸序列(序列号:NM_001281248.1),利用DNA man9.0 软件设计引物,VvGST1-F:AAGAGGCTGGCAAGAAGGAAT,VvGST1-R:CTGTAATCCACCACCAAGTCATAG,引物由上海生工合成。经PCR扩增试验,获得目的条带。将获得的目的条带回收和核酸电泳检测。纯化后的目的片段进行连接。12 h后在4℃条件下转化至DH5a,37℃过夜培养,挑取菌体正常的单克隆,将扩增的菌液进行PCR鉴定,鉴定完成后每片段挑选3例阳性单克隆菌液双向测序,测序列与参照序列进行对比,结果一致可用于后续分析。
1.2.3VvGST1基因的生物信息学分析
将测序结果通过NCBI数据库中的BLAST工具确定完整编码框(http://blast.ncbi.nlm.nih.gov);使用蛋白质查找开放阅读框ORF Finder在线对比(https://www.ncbi.nlm.nih.gov/orffinder);利用ProtParam软件分析蛋白质分子结构和理化特性(http://web.expasy.org/prot-param);利用Prot Scale软件分析蛋白质的疏水性(http://web.expasy.org/protscale/);使用SignalP软件对蛋白质信号肽分析(https://services.healthtech.dtu.dk/);使用TMHMM软件对蛋白质跨膜结构域分析(https://services.healthtech.dtu.dk/);通过Softberry在线软件的ProtComp 9.0预测VvGST1的亚细胞定位;使用SWISS-MODEL软件对蛋白质三级结构预测(http://swissmodel.Expasy.org/)。
2 结果与分析
2.1VvGST1保守序列的扩增及鉴定
研究表明,葡萄基因条带特异性好,清晰度高。所获得的VvGST1的片段长度为719 bp,目的基因长度与预测的结果一致。该条带满足测序反应的要求,已扩增到目的基因片段。随机选其中一个条带胶回收进行后续克隆实验。所有的测序峰图结果是较为单一的峰图,背景较弱无杂峰乱峰,可以用于后续的分析。图1
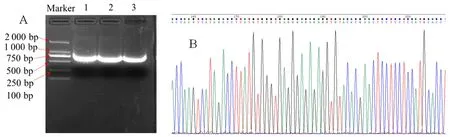
图1 葡萄VvGST1基因PCR扩增电泳图、测序峰图和对比
2.2VvGST1氨基酸序列比对
研究表明,木纳格葡萄中的VvGST1的氨基酸序列为保守序列。该序列与酿酒葡萄GST1(NM_001281248.1)的同源性高达97%,与梅(Prunus mume,XM_008228042.2)、甜杏仁(Prunus dulcis,XM_034357965.1)、桃(Prunus persica,XM_020561748.1)相似度均超过70%,分别为71.94%、71.78%和71.29%。克隆获得的GST是葡萄果实中GST1基因。进一步与其他物种比对氨基酸序列,各物种间的GST基因具有较高的同源性。图2
2.3VvGST1与其它植物GST氨基酸序列系统进化树
研究表明,构建棉花(XM01260636.1)、桃(XM020561748.1)、梅(XM008228041.2)、樱桃(XM021977290.1)、甜杏仁(XM034357965.1)和板栗(DQ673317.1)等植物的GST氨基酸序列进行系统进化树,克隆的VvGST1基因编码蛋白与棉花属GST的亲缘关系最近且属于为Tua类GST基因。图3
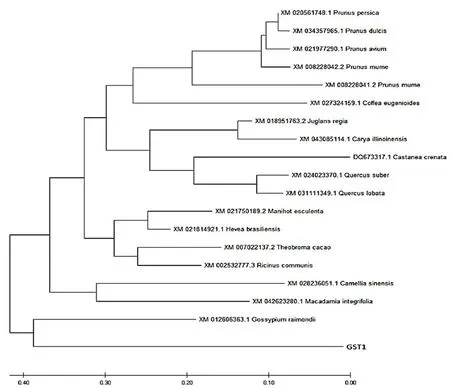
图3 VvGST1与其它植物GST氨基酸序列系统进化树
2.4 VvGST1蛋白质的氨基酸理化特性
研究表明,VvGST1蛋白质分子式为C1178H1799N293O321S11;相对分子质量为25.55 kDa;原子总数3 602;等电点为6.32;推导半衰期在30 h左右。其中,不稳定指数39.22,该蛋白质具有稳定性。VvGST1氨基酸数量为221个,氨基酸种类为20种。相对含量较高为亮氨酸(9.5%)、赖氨酸(9.0%)和谷氨酸(8.1%);氨基酸相对含量较低的是色氨酸(2.3%)和谷氨酰胺(1.8%)。其中带负电荷的天冬氨酸和谷氨酰胺残基总数是30,带正电荷的精氨酸和赖氨酸残基总数是28。图4
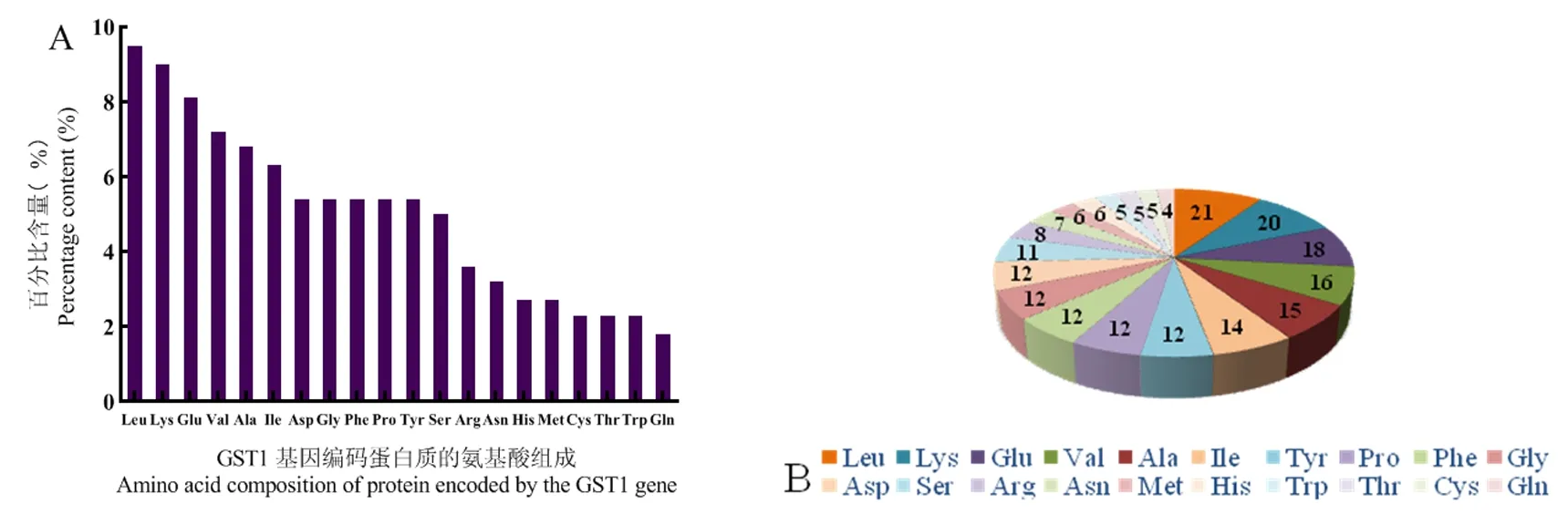
图4 VvGST1基因编码蛋白质的氨基酸组成
2.5 VvGST1蛋白亲疏水性
研究表明,在整个肽链中大多数氨基酸数值在0 ~ -2.5,分布均匀,负值越小,表示该氨基酸疏水性越强,且总亲水性平均数为-0.175,脂肪指数为89.55,该葡萄果实中VvGST1为亲水性蛋白。图5

图5 VvGST1蛋白的亲疏水性
2.6 VvGST1蛋白信号肽预测
研究表明,木纳格葡萄VvGST1没有明显的区段存在信号肽,该序列没有预测到信号肽,意味VvGST1不会产生蛋白运转。图6

图6 VvGST1蛋白信号肽预测
2.7 VvGST1蛋白跨膜结构域预测
研究表明,木纳格葡萄VvGST1蛋白整条链上所有位点均没有存在跨膜结构的可能性,整个蛋白的所有氨基酸膜外的概率均在0.9以上,存在于细胞基质中,说明该蛋白亚单位不含跨膜结构域,是膜外蛋白,为典型的基质蛋白。图7
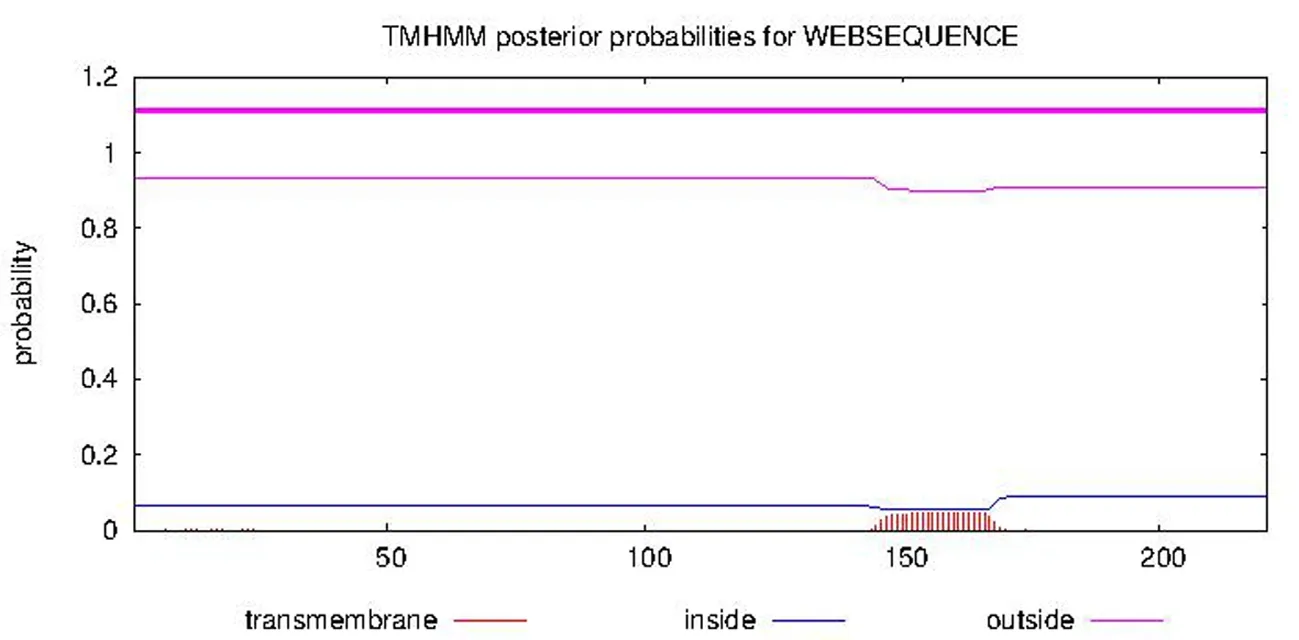
注:膜外(outside)、膜内(inside)和跨膜区域(transmembrane)
2.8 VvGST1蛋白亚细胞定位预测
研究表明,由神经网络预测在细胞外的分值为1.0,预测VvGST1蛋白整体在细胞质的分值为8.64,该蛋白可能是多定位的。利用MultiLocDB中进行SBLAST搜索,该蛋白质的位置可能在细胞质和细胞核中。表1
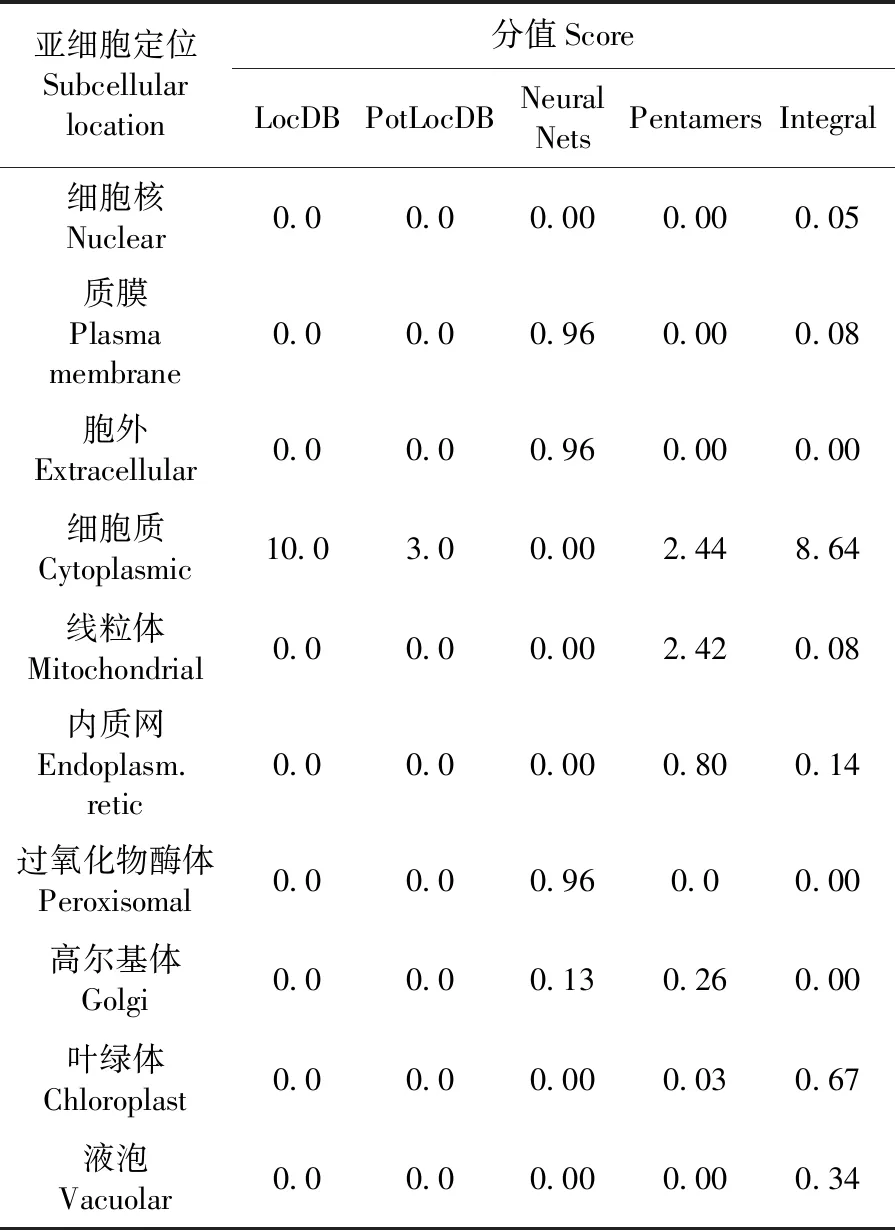
表1 VvGST1亚细胞定位预测
2.9 VvGST1蛋白二级和三级结构的构建
研究表明,VvGST1蛋白主要由4种二级结构组成,包括无规则卷曲、α-螺旋、延伸链和β-折叠等,其中α-螺旋是主要的二级结构占51.58%,其次无规则卷曲占29.84%,延伸链占13.75%,β-折叠占4.98%。图8

注:蓝色:α-螺旋;绿色:β-折叠;黄色:无规则卷曲;红色:延伸链
VvGST1蛋白质的三维结构模型类似山峰型,蛋白三维结构模型与模板的一致性为60.37%,全球性模型质量评估(GMQE)值为0.81,越接近1,该蛋白三级结构建模质量较好。该蛋白的三级结构主要是由α-螺旋组成,该蛋白具有稳定的结果。图9

图9 VvGST1蛋白的三级结构预测
3 讨 论
蛋白二级结构是指沿一定方向折叠或盘旋而形成的特定多肽结构,包括α-螺旋、β-折叠、无规则卷曲和延伸链等[13]。灰霉病是由灰葡萄孢引起的一种真菌性采后病害,是葡萄采后贮藏期间引起腐烂的主要病原菌之一,引起葡萄浆果采后品质的快速下降[14]。开展抗灰霉病相关基因的挖掘对于葡萄采后抵御病原菌侵染具有重要作用。GST基因在植物响应生物与非生物胁迫方面发挥着重要作用,并具有一定的耐受能力[15]。GST家族可根据结构特征和序列的同源性分类分别是Phi、Tua、Lambda、Theta、Zeta、DHAR和TCHQD,其中植物所特有的是Phi和Tua类基因[16]。
试验从木纳格葡萄果实中成功克隆了VvGST1全长序列。该序列为完整的开放阅读框,保守的编码长度是719 bp,其氨基酸蛋白编码长度与山葡萄[17]、荔枝[18]相似。VvGST1与其他物种氨基酸序列多重对比和进化树系统构建可看出,研究克隆的VvGST1基因编码蛋白与棉花属GST的亲缘关系最近且属于Tua类GST基因。不同物种间GST的氨基酸序列在进化过程中是保守的。研究表明,氨基酸保守性越高,就越可能是功能位点,其突变对功能影响就越大[19]。
理化特性分析表明VvGST1共编码氨基酸221个,包含20种氨基酸,pI 6.32;相对分子质量为25.55 kDa。VvGST1蛋白不稳定指数为39.22,表明该蛋白质具有稳定性。分子的亲水性是指其有能力形成氢键的位点,并且和非极性或其他疏水性溶液对比,亲水性分子更易溶解在水里面[20]。VvGST1蛋白是具有稳定性的亲水性蛋白,该蛋白不能进行膜内外运动,不含跨膜结构域,没有预测到信号肽。从亚细胞定位可以发现该蛋白存在细胞质中与跨膜结构域预测结果一致。三级结构是指蛋白质通过充分旋转折叠的、并具有生物活性的、能完成生物代谢功能的一个完整球蛋白的三维立体结构[20]。VvGST1蛋白的二级结构和三维结构模型主要由α-螺旋组成,其中三级结构模型中GMQE值为0.81,当越接近1时建模质量越好。VvGST1基因编码蛋白质的二级和三级结构十分稳定。这与蓖麻克隆GST蛋白三维结构相似,该蛋白可能与蓖麻蛋白具有的功能相似[21]。葡萄VvGST1含有丰富的α-螺旋结构,其可能指导VvGST1生物合成能够响应于生物胁迫因子或非生物胁迫因子的诱导,参与细胞对外界胁迫的抗性反应[5]。苹果受到轮纹病侵染时VvGST1基因表达上调表达并克隆了抗苹果轮纹病相关的MdGSTU1[9]。在水稻和小麦中分别分离出了参与植物抗病胁迫响应的GST相关基因[22];拟南芥与番茄中的多个GST被证明了能够对真菌侵染做出响应提高抗性能力[23]。植物GSTs的表达受到非生物和生物胁迫的激活。
4 结 论
在木纳格葡萄中克隆了1个GST1基因,该基因片段长度为719 bp,具有完整的开放阅读框。VvGST1基因编码蛋白质的相对分子质量为25.55 kDa;理论等电点为6.32;分子式为C1178H1799N293O321S11;原子总数3 602;理论推导半衰期为30 h左右;不稳定指数为39.22。VvGST1基因编码的氨基酸种类及个数分别为20种和221个。VvGST1蛋白的二级和三级结构构建分析主要是由α-螺旋组成,该蛋白具有十分稳定的结构。
