基于涡度系统的干旱区枣林能量平衡分析
2023-09-07张百和王腾飞
乔 英,张百和,邱 凯,王腾飞
(新疆理工学院,新疆 阿克苏 843100)
0 引 言
【研究意义】生态系统中地表能量是陆-气之间能量和物质传输、转化的基础,也是生态系统中一切过程和功能的动力,而地表能量研究的核心问题是以太阳能输入为驱动的有机化学能循环及能量交换规律[1]。所以,深入探究生态系统的能量循环规律,不仅有助于了解区域气候系统的能量和物质循环以及气候变化过程[2],而且还为维持生态系统的能量和水碳循环提供重要的科学依据[3]。【研究进展】能量平衡闭合率(Energy balance ratio,EBR),表示为生态系统的地表有效能量(潜热通量LE+显热通量H)与可利用能量(净辐射Rn+土壤热通量G)的比值[4],国内外学者们深入分析了多种生态系统的能量平衡,Wilson 等[4]对FLUXNET 的22 个通量站的50 个站点年进行能量平衡分析,结果显示EBR在53%~99%之间,平均值为79%;李正泉等[5]对ChinaFLUX 的8个通量站进行能量平衡分析,结果显示EBR平均约为73%。国内外学者们的研究结果均显示地表的能量均无法闭合,能量平衡闭合率均在52%~99%之间[4-8],生态系统种类包括落叶阔叶林[9]、农田生态系统(稻麦轮作)[10]、干旱区人工柠条林[11]、热带季雨林[12]等。
目前,涡度协方差技术(Eddy covariance technique,EC)作为监测生态系统能量交换过程的最有效、最准确的方法,已经得到了微气象学家和生态学家们的认可,成为检验其他测定方法、评价各类模型模拟精度的标准方法[13]。枣树因耐旱、喜光照、结果早,兼顾经济和生态效益[14]被成功引进新疆,目前种植面积已达50 万hm2,成为新疆特色经济林果业的支柱[15]。阿克苏地区枣树的种植面积(占全疆34.34%)和产量(占全疆54.70%)均为新疆首位[15]。【切入点】当前国内外学者对农田生态系统的研究多为一年生作物,对于人工经济林的研究相对较少,干旱区枣林生态系统的能量平衡分析及各能量分项的变化规律还需进一步探索。
【拟解决的关键问题】综上,本文以干旱区人工枣林生态系统为研究对象,利用涡度相关系统在田间开展2 a 的观测试验,采用REddyProc 包[16]插补通量缺失数据,计算干旱区人工枣林的热储存项,评价枣林能量平衡闭合率,并量化和分析枣林生态系统的能量变化规律和能量分配规律,为干旱区枣林的水碳循环研究和优化水碳管理模式提供参考。
1 材料与方法
1.1 研究区概况
研究区位于新疆阿克苏市农业示范园(80°18'—80°22'E,41°05'—41°07'N,海拔1 198 m),面积约170 hm2。研究区气候干旱少雨属温带沙漠气候,年均太阳总辐射量5 671.36 MJ/m2,年均日照时间2 911 h,年均降水量68.40 mm,年均气温11.20 ℃,年均蒸发量1 993 mm[17]。研究区土壤质地为砂土,砂粒量87.27%,土壤密度为1.63 g/cm3,田间持水率为8.93%,地下水埋深大于10 m。试验作物为枣树,品种为灰枣,株行距2 m×4 m,冠层高度为(4±0.2)m[18]。枣树每年4 月10 日—10 月20 日为生育期,10 月21日—次年4 月9 日为休眠期。
1.2 试验方法
1.2.1 数据采集
实验仪器有涡度相关系统(IRGASON 型一体机)、四分量辐射计(CNR4 型)、土壤温度计(TCAV 型)、土壤热通量板(HFP01 型)、土壤水分探头(Hydra ProbeⅡ型)、数据采集器(CR3000 型)[14],详见表1。

表1 阿克苏站点实验仪器汇总Table 1 Summary of experimental instruments at the Aksu site
1.2.2 通量数据计算及数据插补质量控制
使用软件LoggerNet 4.5(Campbell Science Inc,USA)按照标准数据质量控制程序在线计算[19],并根据稳态测试、完整湍流特征、超声风速仪的水平定位等,按照微气象学经验将数据质量划分为0~9 级[14]。根据数据质量分级,将质量差的数据删除并使用R 语言的REddyPrco 包[16]进行数据插补,并采用交叉验证法来评价插补数据的质量[14];同时,使用能量闭合率来检验插补数据质量。
选择2018 年和2019 年的30 min 尺度数据,采用Kljun 等[20]和Kormann 等[21]的源区模型计算阿克苏站点的通量贡献区,90%的贡献区(通量源区)的圆形半径分别为356.50、350.00 m。
1.2.3 能量平衡计算
根据能量守恒定律,生态系统能量平衡可用式(1)表示[22],其中的热储存项S可用式(2)表示[23],热储存项的计算方法参见Dou 等[24]文献计算。能量平衡闭合率(Energy balance ratio,EBR)是指地表有效能量(潜热通量LE+显热通量H)与可利用能量(净辐射Rn-土壤热通量G)之比[25],EBR可用式(3)计算[26]。为了提高能量平衡闭合率,很多研究也将热储存项S记入可利用能量中,EBR可用式(4)计算[26]。
式中:Rn为太阳净辐射(W/m2);G为土壤热通量(W/m2);H为显热通量(W/m2);LE为潜热通量(W/m2);S为热储存项(W/m2);Q为其他来源的能量(忽略不计,W/m2);Ss为一定深度范围内的土壤热储存项(W/m2);Sa为大气显热储存项(W/m2);Se为大气潜热储存项(W/m2);Sc为冠层热储存项(W/m2);Sp为植物光合储存项(W/m2);St为树木枝干热储存项(W/m2)[23]。
2 结果与分析
2.1 气温、降水量和灌溉量的变化情况
观测期(2018、2019 年)日平均气温、降水量、灌溉量如图1 所示。观测期间,日平均气温在-12.72~29.16 ℃之间变化,年平均气温为10.99 ℃,其中月平均气温最低出现在2018 年12 月(-8.36 ℃)和2019 年1 月(-8.45 ℃),月平均气温最高出现在2018 年7 月(24.07 ℃)和2019 年7 月(24.74 ℃),6—8 月的月平均气温在20 ℃以上。降水集中在4—10 月(生育期)内,2018、2019 年年降水量分别为81.50、57.40 mm,单日最大降水量为13.20 mm。灌溉集中在4—10 月,2018、2019 年灌溉量分别为847.15 mm(灌溉31 次)、848.25 mm(灌溉30 次)。
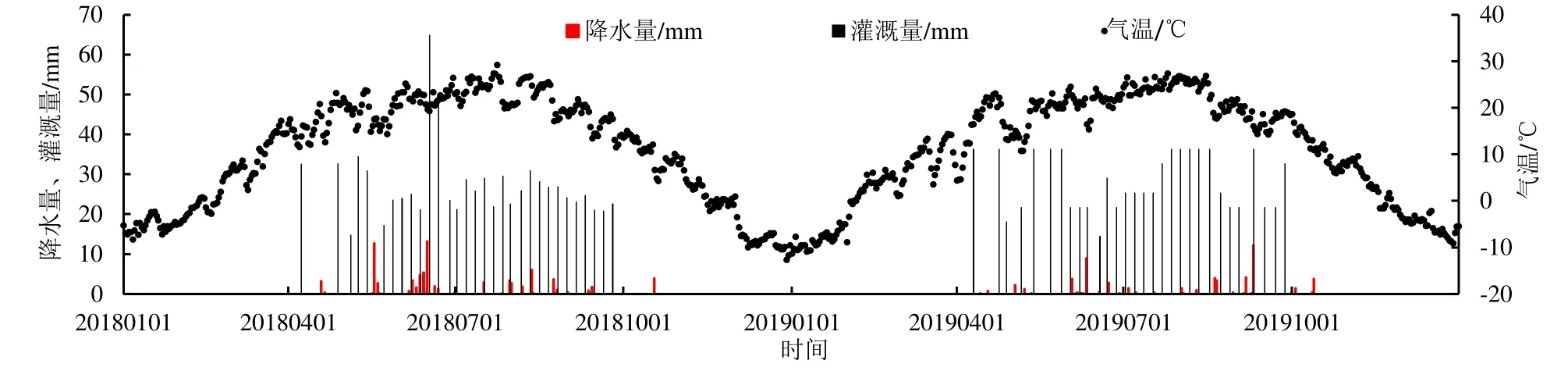
图1 研究区2018年和2019年气温、降水量及灌溉量Fig.1 Distribution of temperature, precipitation and irrigation of the study area in 2018 and 2019
2.2 能量通量值
使用REddyPrco 包插补后的通量数据及式(2)计算了干旱区枣林的能量分项,详见表2。由表2 可知,2018、2019年净辐射Rn年度合计为1 475 227.47、1 481 026.93 W/m2,在生育期明显大于休眠期;2018、2019 年潜热通量LE年度合计为898 349.68、922 381.08 W/m2,潜热通量主要集中在枣树生育期,在休眠期较小;2018、2019年显热通量H年度合计为205 778.37、183 786.36 W/m2,主要集中在休眠期,在生育期较小;2018、2019 年土壤热通量G年度合计为-15 839.17、-13 877.22 W/m2,在生育期为正,休眠期为负;且Rn、LE、H、G在2018 年和2019 年情况相似。由表2 可知,显热热储存项Sa为S中的最大项,2018、2019年Sa年度合计分别为22 053.05、17 847.04 W/m2,占净辐射Rn的1.49%、1.21%;其次是植物光合热储能Sp,2018、2019 年Sp年度合计分别为13 153.85、11 345.06 W/m2,占净辐射Rn的0.89%、0.77%;土壤热储存项Ss、冠层热储存项Sc、潜热热储存项Se在2018 年和2019 年的年度合计均较小。
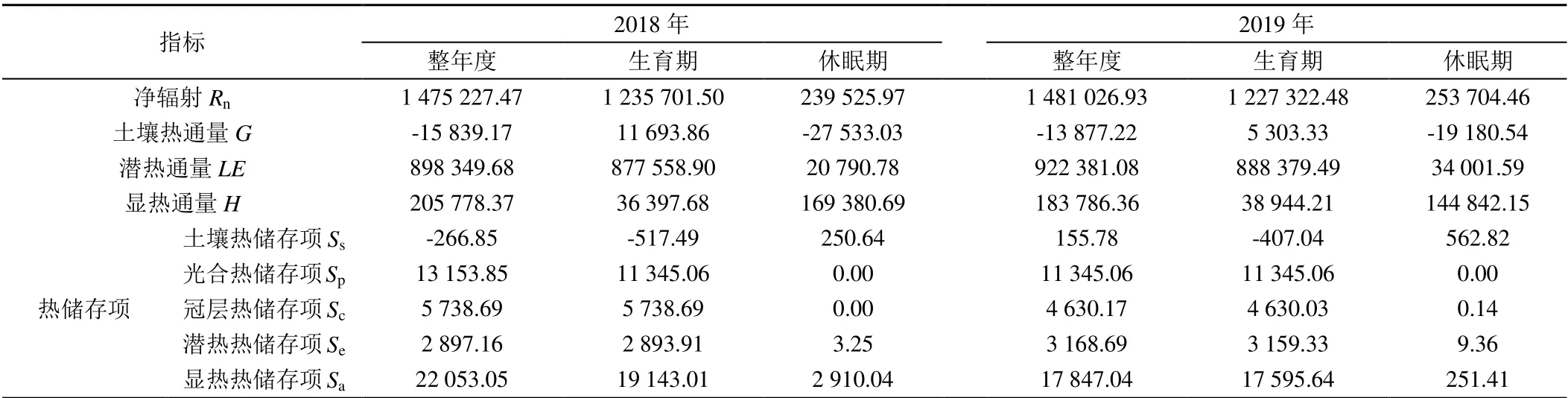
表2 干旱区枣林的能量分项Table 2 Energy subdivision of date jujube forests in arid areas W/m2
2.3 能量通量的月平均日变化特征
2018、2019 年REddyProc 插补后的能量通量(净辐射Rn、潜热通量LE、显热通量H、土壤热通量G)的月平均日变化规律见图2。由图2 可知,Rn、LE、H、G的月平均日变化均呈单峰变化趋势,2018 年和2019 年各月的能量变化趋势相似。Rn、LE、H、G在白天为正,夜间为负,正负转换的交点与白昼变化的时间一致。各月Rn的最小值均为负,在-74.16~-47.51 W/m2之间波动;Rn在夜间为负值,在白天为正值,说明在夜间地表损失热量,而在白天吸收热量。2018年Rn的最大值为653.05 W/m2,出现在7 月;2019年Rn的最大值为638.98 W/m2,出现在6 月;2 a 最大值均出现在15:30。2018 年和2019 年LE的最大值均出现在8 月,最大值分别为331.85、290.20 W/m2,且最大值出现时间和Rn一致,均在15:30。LE在生育期明显大于休眠期,在5—9 月,LE为Rn的主要分配项。H在2018 年和2019 年的变化幅度相似,在-80.51~180.45 W/m2之间波动,2018 年H的最大值为180.45 W/m2,出现在4 月;2019 年H的最大值为165.06 W/m2,出现在5 月。H在1—4 月和10—12月为Rn的主要分配项。G在2018 年和2019 年的变化幅度相似,在-24.09~71.16 W/m2之间波动,2018年G的最大值为34.52 W/m2,出现在4 月;2019 年G的最大值为71.16 W/m2,出现在3 月;G占Rn的比值较小。

图2 2018、2019年干旱区枣林1—12月能量的月平均日变化规律Fig.2 Monthly average daily variation of energy in jujube forests in arid areas from January to December 2018 and 2019
2.4 干旱区枣林生态系统的能量平衡分析
2018、2019 年干旱区枣林生态系统各月能量平衡闭合线性回归参数和能量平衡闭合率情况见表3。表3 显示,未计入热储存项时,各月的有效能量和可利用能量之间的统计回归斜率k为0.32~0.69,2018年k值最低为0.37,出现在1 月,2019 年k值最低为0.32,出现在12 月;各月能量平衡闭合线性回归参数决定系数R2为0.60~0.92;斜率k和R2在3—10 月之间变化较平缓,极小值出现在11—12 月、1—2 月。各月的能量平衡闭合率在34.05%~80.78%之间,闭合率最高为80.78%(2019 年9 月);闭合率最低为34.05%(2018 年12 月);未计入热储存项时,表明各月的能量平衡闭合程度较好。由于受太阳高度、枣林生育期中地表覆盖度和叶面积指数、灌溉、气象等因素变化的影响,各月的线性回归斜率和能量平衡闭合比率存在一定的波动。计入热储存项后,各月的回归斜率k、R2、能量平衡闭合率均有所提高,但增加幅度较小。表3 显示,枣树生育期的能量平衡闭合率明显高于休眠期,同时线性回归斜率和R2也是生育期明显大于休眠期;计入热存储项后,k、R2、EBR也均有提高。
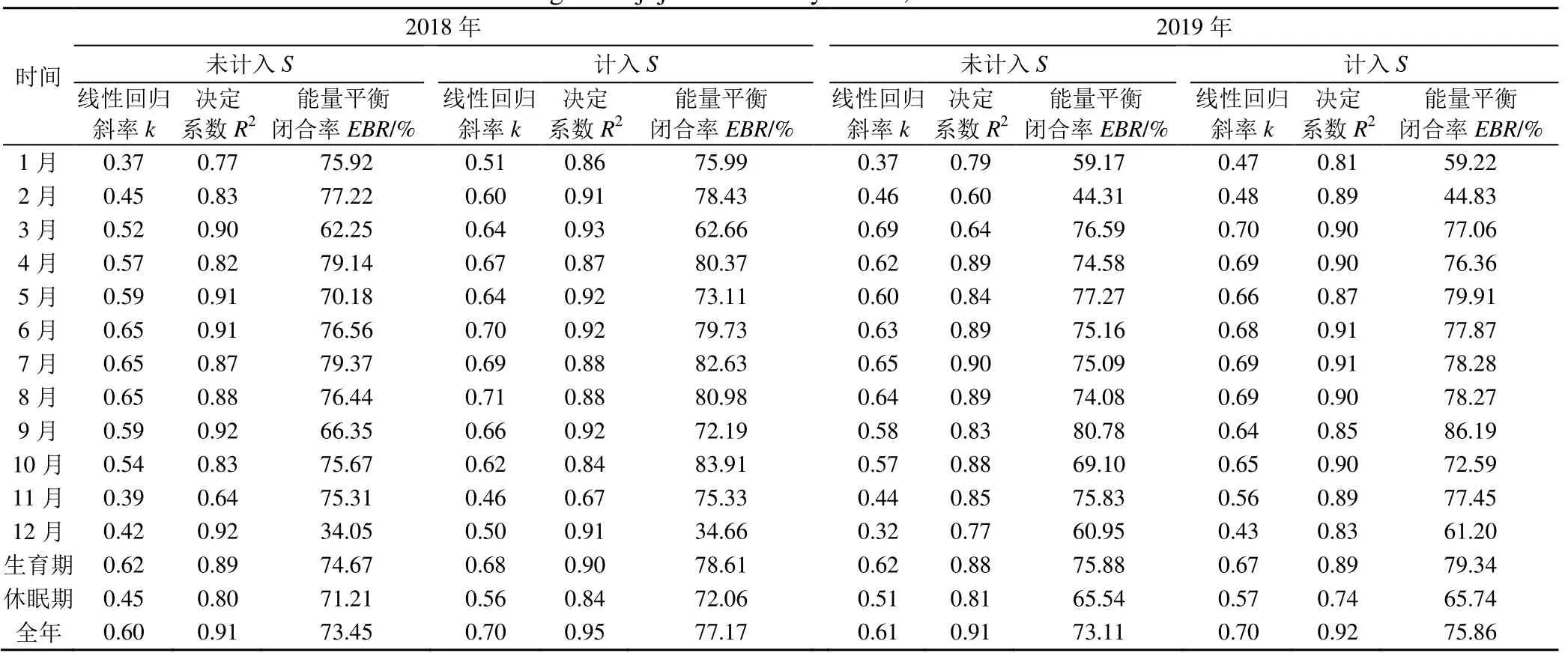
表3 2018、2019年干旱区枣林各月能量平衡闭合线性回归参数和能量平衡闭合率Table 3 Energy balance closure linear regression parameters and energy balance closure rates in arid region of jujube forests by month, 2018 and 2019
干旱区枣林生态系统30 min 尺度的能量平衡如图3 所示。由图3 和表3 可知,未计入热储存项的干旱区枣林生态系统2018、2019 年30 min 尺度上的有效能量(H+LE)与可利用能量(Rn-G)的直线拟合斜率为0.60(R2=0.91)和0.61(R2=0.91),能量平衡闭合率分别为73.45%、73.11%。将热储存项计入可利用能量,2018 年和2019 年(H+LE)与(Rn-G-S)的直线拟合斜率为0.70(R2=0.95)和0.70(R2=0.92),记入热储存项后能量平衡闭合率有所提高,干旱区枣林2018 年和2019 年的能量平衡闭合率分别为77.17%、75.86%,且2 a 的闭合率相似。学者们[5-10]认为生态系统能量平衡不闭合程度在10%~30%之间,本文研究结果在此范围内,说明涡度相关法在干旱区枣林生态系统通量观测中可靠性较高,下垫面符合观测范围;同时,也证明REddyPrco 包插补的通量数据质量较好。

图3 干旱区枣林生态系统30 min尺度的能量平衡Fig.3 Energy balance of date Jujube forests ecosystems in arid areas at the half-hourly scale
2.5 生育期和休眠期的热储存项分项分析
土壤热储存项Ss、显热热储存项Sa、潜热热储存项Se、冠层热储存项Sc和光合热储存项Sp在枣树生育期和休眠期的日平均变化见图4。

图4 枣林的能量分项在生育期和休眠期的日平均变化Fig.4 Average daily changes in the energy subdivision of date Jujube forests during the reproductive and dormant periods
图4(a)显示,2018 年和2019 年土壤热储存项Ss日平均变化在生育期和休眠期规律相似,差异不大,均在-51.17~69.15 W/m2之间呈单峰变化,与土壤热通量变化规律相似。Ss在白天为正值,表示为吸收热量;而在夜间为负值,表示为释放热量。图4(b)显示,2018 年和2019 年潜热热储存项Se日平均变化规律在生育期和休眠期不同:在休眠期潜热热储存项的变化范围在0 附近,变化幅度较小,为±0.05 W/m2,呈单峰变化;在生育期潜热热储存项在-0.23~0.58 W/m2之间呈双峰变化规律,在09:00 和20:00 均有峰值,说明在昼夜交替的时候潜热热储存项变化较大。光合热储存项Sp和冠层热储存项Sc在休眠期均为0,所以图4(c)只绘制了生育期的光合热储存项和冠层热储存项的平均日变化。图4(c)显示,光合热储存项Sp呈单峰变化规律,在夜间为0,处于0~2.95 W/m2之间,Sp随着净辐射的变化而变化,当净辐射在15:00达到峰值时,Sp继续升高在18:00 左右达到峰值,并随着净辐射的减小而迅速降低。2 a 的冠层热存储项Sc均较小,在-0.39~1.55 W/m2之间呈单峰型变化。Sc在夜间变化平缓,变化幅度小;白天随着净辐射的增大而增大,到达峰值时间与净辐射一致。图4(d)显示,2018 年和2019 年显热热储存项Sa在生育期和休眠期变化规律不同。生育期的显热热储存项在-0.80~6.50 W/m2之间呈单峰变化;Sa在夜间变化平缓幅度小,白天随着净辐射的增大而迅速变大,在12:00左右达到峰值,峰值持续6 h 左右,在18:00 之后缓慢降低。休眠期的显热热储存项在-2.74~3.35 W/m2之间变化,其变化幅度比生育期小,在夜间为负,白天为正;日出后,随着净辐射迅速升高,Sa也迅速增大,在12:00 左右达到峰值,然后逐渐减小。
3 讨 论
3.1 枣林生态系统的能量分配特征
在太阳辐射能量的驱动下,农田生态系统内进行能量流动、物质合成转移和水碳循环等运动过程。因下垫面的作物类型、种植密度、生育期、季节、昼夜、气象、人为干扰等差异,会导致净辐射进入生态系统后,各个能量分项等在系统内的分配变化存在差异[27]。同时,生态系统的能量分配变化规律也可以反映下垫面的干湿状况[28]。
5—10 月,干旱区枣林生态系统的LE/Rn的比值大于50%,LE/Rn的比值最大出现在2018 年8 月(92.10%)和2019 年9 月(96.43%)。在4 月和11月,LE/Rn比值在17.33%~35.18%之间,在1—3 月和12 月,LE/Rn的占比均较小,在3.35%~12.25%之间。分析其原因,本文研究区是干旱区,年降水量较小,作物生长依赖于灌溉,枣林只在生育期进行灌溉,时间为每年的4—10 月,灌溉后,土壤湿润,土壤蒸发较大,所以5—10 月干旱区枣林LE/Rn的占比大于50%,说明净辐射主要用于下垫面的LE,即蒸散(包括植株蒸腾和土壤蒸发)。而4 月的LE/Rn为27.23%和33.41%,是因为4 月枣林虽然进行了灌溉,土壤湿润导致土壤蒸发较大,但是枣树处于萌芽期,叶面积指数较小导致植株蒸腾较小,综合起来表现为LE/Rn比3 月有明显增加(增幅为21.57%和21.16%)。LE/Rn在8—9月最大,处于86.47%~96.43%之间,此时也是枣林灌溉量最大、叶面积指数最大的时期。这表明在枣林的生育期(5—10 月)能量分配由LE占据主导。
在1—4 月和11—12 月,枣林生态系统中的H为Rn的最大支出项;在1—3 月和11、12 月,H/Rn占比均大于50%,在4 月H/Rn占比接近50%(2018 年为47.59%,2019 年为45.19%),这表明,在枣树休眠期,能量分配由H占据主导。Trenberth 等[29]分析了全球能量收支,认为地表吸收的能量有近1/2 消耗在潜热,同时,能量分配方式与生态系统中的水分平衡受大气水分供需影响。也有学者[2,6,9]进一步研究表明,植被覆盖较好的生态系统(农田、森林和草甸等)能量分配由LE占据主导,这也验证了本文的研究结果,对于干旱区的枣林生态系统能量分配来说,水分条件是决定能量分配的关键因素,若生态系统水分充足,能量分配则以潜热通量为主,反之则以感热通量为主[30-31]。干旱区枣林的能量分配方案由水分条件决定,受到水分条件、植被类型和气候特征的影响[32]。
3.2 能量平衡闭合率
干旱区枣林生态系统在2018 年和2019 年的全年能量平衡闭合率为73.45%、73.11%,这与前人[4-12]在其他的生态系统中观测的能量平衡闭合率相似,在合理的范围内(能量平衡不闭合率处于10%~30%之间)。根据Kljun 等[20]和Kormann 等[21]的源区模型计算出的下垫面90%通量贡献区的圆形半径分别为356.50、350.00 m,而实际试验区的面积覆盖90%通量贡献区;试验区的下垫面范围是合适的,涡度仪器的观测高度也是合适的,这说明在干旱区枣林使用涡度相关观测系统测量能量是可行的,得到的观测数据可信度高。
干旱区枣林生态系统的全年能量平衡闭合程度不高,这可能是由于忽略了热存储项,于是本文利用式(4)重新计算了能量平衡闭合率,结果显示记入热存储项后,2018 年和2019 年的EBR提高了3.72%、2.75%,达到了77.17%、75.86%,与美国温带落叶林[33]、加拿大北部杨树林[34]、西双版纳热带季节雨林[24]的增幅范围相似;但是比黄土高原柠条林[23]、黑河中游生育期的春小麦农田[22]等的增幅低。多位学者指出[13,22-23],下垫面土壤热储量Ss为热储存项中的最大占比,忽略Ss是造成地表能量平衡不闭合的主要原因。但是根据表2 显示,Ss占净辐射的比例小于1%,这与岳平等[2]、高红贝等[22]研究结果不一致。这可能是因为本文研究区处于干旱区,作物需要灌溉,枣林使用深井水进行滴灌,井水温度低于灌溉时的气温和土壤温度,导致枣林每次灌溉时降低了土壤热储量Ss,故本文虽然计算了Ss,但是EBR提高幅度小,Ss占净辐射的比例小于1%。
4 结 论
1)Rn、LE、H、G的日均值均呈单峰变化趋势,2018 年和2019 年的变化趋势相似。
2)未计入热储存项时,各月的有效能量和可利用能量之间的统计回归斜率为0.32~0.70,决定系数R2为0.64~0.94,能量平衡闭合率在34.05%~80.78%之间;计入热储存项后,各月的有效能量和可利用能量之间的统计回归斜率、决定系数、能量平衡闭合率均有所提高,但增幅较小。
3)2018 年和2019 年未计入热储存项的全年能量平衡闭合率分别为73.45%、73.11%;计入热储存项后,能量平衡闭合率有所提高,但增幅较小。
4)干旱区枣林生育期的能量平衡闭合率高于休眠期;计入热存储项后,生育期能量平衡闭合率的增幅也明显大于休眠期。
5)土壤热储存项的日平均变化在生育期和休眠期规律相似;潜热热储存项的日平均变化在休眠期呈单峰变化,生育期呈双峰变化;光合热储存项在生育期呈单峰变化;冠层热存储项在生育期呈单峰变化;显热热储存项在生育期呈单峰变化,休眠期波动幅度较小。
6)干旱区枣林的能量分配受水分条件、植被类型和气候特征的影响,且能量分配方案由水分条件决定,若生态系统水分充足,能量分配则以潜热通量为主,反之则以感热通量为主。
(作者声明本文无实际或潜在的利益冲突)
