芽变黄叶银杏光合和叶绿素荧光特性*
2023-09-02薛芳婷刘新亮刘玉华余鹏飞郁万文
薛芳婷,刘新亮,刘玉华,余鹏飞,郁万文
(1.南京林业大学 南方现代林业协同创新中心,江苏 南京 210037;2.江西省林业科学院 国家林业草原樟树工程技术研究中心,江西 南昌 330032;3.江苏农林职业技术学院 现代林业技术创新研究院,江苏 句容 212499;4.睢宁县润企投资有限公司,江苏 睢宁 221200)
叶色突变体是研究叶绿素代谢、叶绿体发育和光合作用的理想材料。它是园林绿化植物中常见的显性突变形式,其具有多样的突变类型,如白化、黄化、条纹、黄绿、绿黄、绿白、浅绿、翠绿等。这些叶色突变体不仅突变多样、性状明显,而且观赏价值高,为创制新种质和选育新品种提供了物质基础。叶片黄化植物根据其成色时期可分为春色叶、秋色叶和常色叶3类,其中常色叶在整个生长期内都呈现非绿叶色,通常是由于植物体内遗传物质发生改变,导致色素代谢失衡,从而引起叶色改变,叶色黄化仅仅是一种表观特征,在这种表型下,叶片结构、生理生化特性也会发生变化[1]。研究发现,不同黄化突变植物的叶绿素代谢及叶绿体发育的变异存在差异,其对光合作用的影响也各不相同。一般情况下,叶绿素缺乏会导致光合作用的捕光效率降低,从而减小天线系统的大小。大部分突变体中光系统II的光化学效率较野生型低,但个别叶绿素b缺乏的突变体表现较高[2]。叶绿素缺乏对光合效率的影响也有所差异,叶色突变体的光合效率通常较野生型低,但也有个别突变体表现较高[3-5]。目前,在园林绿化中广泛应用的黄化突变木本树种包括金叶银杏(Ginkgobilobacv.jinye)、金叶榆(Ulmuspumilacv.jinye)、金叶国槐(Sophorajaponicacv.jinye)、普林斯顿黄金枫(Acerplatanoides‘Princeton Gold’)、金叶复叶槭(Acernegundo‘Aurea’)、金叶白蜡(Fraxinuschinensis‘Aurea’)、金叶女贞(Ligustrumlucidum‘Vicaryi’)、金叶黄杨(Buxussempervives‘Latifolia Maculata’)等,这些黄化突变体是研究叶绿素代谢、叶绿体发育、光合作用及呈色机理的理想材料[6-7]。
芽变黄叶银杏(Ginkgobiloba)是一种具有优美叶色和长久观赏期的植物,具有较高的景观价值。学者们通过对黄叶银杏及其子代苗的生长[8]、叶色参数[8,10,12]、色素含量[8,10,12]和光合生理[9,11]年动态变化,以及其超微结构[12]、合成前体物代谢[13]、蛋白组学[14]等多个维度的研究,探索其呈色机理[12-15]。然而,目前对于黄叶银杏光合生理生态年动态特征的了解还不充分,这在一定程度上限制了黄叶银杏的栽培管理和生产应用。因此,本研究选取母株正常枝上绿叶和芽变枝上黄叶作为试材,比较两者的色素含量、超微结构、光合气体交换和叶绿素荧光参数的差异性,以探究黄化对银杏叶的光合和叶绿素荧光特性的影响机制,旨在揭示黄叶银杏的光合作用调控机理,为进一步利用黄叶银杏突变体提供基础。
1 材料与方法
1.1 材料
江西省彭泽县的黄叶银杏母株(图1),树龄150年,雌株。根据叶色变化进程及发育时期(展叶至落叶),于4月27日(全展嫩叶)、5月28日(成熟叶)、8月2日上午830—900,避开雨天,选择南面向阳的同一高度的大小相似的一年生枝条上的黄叶和绿叶,液氮速冻后,带回实验室-80 ℃保存备用。

图1 芽变黄叶银杏母株
1.2 测定项目与方法
1.2.1 光合色素含量测定
参照唐延林等[16]的方法,用丙酮乙醇混合液(丙酮、无水乙醇、蒸馏水的体积比为4.5︰4.5︰1)提取,用UV-1900紫外可见分光光度计测定叶绿素a、叶绿素b 和类胡萝卜素的含量。每个处理3个生物学重复。取4月27日、5月28日、8月2日叶片,将叶片剪碎混匀,称取0.1 g 置于10 mL有盖离心管中,加入丙酮乙醇提取液10 mL,浸提24 h至叶片无色,期间震荡3次。以丙酮乙醇提取液作为对照,在665、649、470 nm 波长下联合测定OD 值A665、A649、A470。
叶绿素a 的浓度Ca(μg/mL)=13.95A665-6.88A649;
叶绿素b 的浓度Cb(μg/mL)=24.96A649-7.32A665;
类胡萝卜素的浓度Cc(μg/mL)=(1 000A470-2.05Ca-114.81Cb)。
1.2.2 电镜样品的制备和观察
将叶片用锋利刀片切取2 mm×1 mm小块,放入盛有2.5%的戊二醛固定液(pH=7.2,0.1 mol/L PBS配备)中,将瓶盖盖紧,用大口径注射器抽空叶片中气体,最终样品完全沉落到瓶底。4 ℃固定16 h,经0.1 mol/LPBS(pH=7.2)冲洗3次,然后在30%、50%、70%、80%、90%、100%的酒精梯度中逐渐脱水,每次20 min。其后环氧丙烷浸透两次,再用环氧丙烷︰包埋剂=1︰1溶液浸透6 h,1︰3溶液中浸透12 h,100%包埋剂浸透24 h后,用Epon 812环氧树脂包埋,用LKB-V型超薄切片机进行切片,经醋酸铅染色30 min及柠檬铅染色15 min后,在H-600型透射电镜下观察并拍照。以4月27日、5月28日和8月2日黄叶和绿叶为材料,观察其超微结构。
1.2.3 光合气体交换参数测定
选取固定枝条上部外观正常向阳的均匀一致叶片,于晴天上午900—1100用CIRAS-2便携式光合作用测定系统(美国,PP Systems),在光强1 500 μmol/(m2·s)、CO2浓度400 μmol/mol(由CO2钢瓶提供),温度与湿度为环境水平条件测定光合气体交换参数净光合速率(Pn)、蒸腾速率(E)、气孔导度(Gs)和胞间二氧化碳浓度(Ci)。每片叶读数3次,每次测定15片。计算公式如下:
瞬时水分利用效率WUE=Pn/Tr
①
瞬时羧化效率CE=Pn/Ci
②
气孔限制值Ls=1-Ci/Ca
③
式中:Ca为大气CO2浓度。
1.2.4 叶绿素荧光动力学参数的测定
参照Xu等[17]的方法。于晴天上午900—1100采用FMS-2脉冲调制式荧光仪(Hansatech,英国)测定光适应后的最大荧光(Fm′)、光下初始荧光(Fo′)、稳态荧光(Fs),计算荧光参数Fv′(光下最大可变荧光强度,Fv′=Fm′-Fo′)、Fv′/Fm′(PS II有效光化学量子产量);在充分遮光暗适应(1 h以上)后,测定初始荧光(Fo)、最大荧光(Fm),计算Fv(黑暗中最大可变荧光强度,Fv=Fm-Fo)、Fv/Fm(最大光化学效率)、φPS II(PS II实际光化学反应量子效率)、qP(光化学淬灭系数)和ETR(表观光合电子传递效率)等叶绿素荧光参数。计算公式为qP=(Fm’-Fs)/(Fm′-Fo′);φPSII=(Fm′-Fs)/Fm′;NPQ=(Fm-Fm′)/Fm′;ETR=φPSII×PAR×0.5×α,式中PAR为光通量密度,α为叶片吸光系数,一般为0.84。
参照Demmig-Adam等[18]提出的公式计算叶片吸收光后的光能分配。光化学反应的份额(P)、天线热耗散的份额(D)、用于反应中心由非光化学反应耗散的份额(Ex)的计算公式分别为:
P=Fv′/Fm′×qP
④
D=1-Fv′/Fm′
⑤
Ex=Fv′/Fm′×(1-qP)
⑥
1.3 数据分析
采用Microsoft Excel 2021进行数据处理和制表,并采用SPSS 17.0软件进行统计分析和显著性检验(Duncan’s新复极差法),数据结果以平均值±标准差表示。
2 结果与分析
2.1 光合色素含量的变化
由表1可知,4月和5月黄叶中叶绿素a(Chla)、叶绿素b(Chlb)和总叶绿素(Chl)含量均极显著低于绿叶(P<0.01),8月黄叶中Chla、Chlb和Chl含量则显著低于绿叶(P<0.05),其中Chla含量降低21.3%~69.5%,Chl b含量降低80.0%~91.4%,Chl含量比绿叶降低54.0%~78.7%,表明黄叶叶绿素极为缺乏,且在4月份尤为显著。3个时期的黄叶Chla、Chlb、Chl含量均呈升高的变化趋势,叶色逐渐转绿,其中8月Chla和Chl含量极显著高于4月和5月(P<0.01);而绿叶Chla含量降低,Chlb含量升高,Chl含量差异不显著。4月黄叶类胡萝卜素(Car)含量与绿叶差异不显著,而5月和8月的Car含量极显著高于绿叶(P<0.01)。3个时期的黄叶Car含量呈升高的变化趋势,但升幅不大;而绿叶Car含量则极显著降低(P<0.01)。值得注意的是,3个时期黄叶的叶绿素a/b(Chla/b)极显著高于绿叶(P<0.01),且呈现降低的变化趋势,说明黄叶叶绿素b含量相对于绿叶的降幅较大,且高于叶绿素a的下降幅度,表明黄叶中叶绿素b的合成受阻更为严重。3个时期的黄叶和绿叶类胡萝卜素与叶绿素的比值(Car/Chl)均呈下降的变化趋势,且黄叶Car/Chl均极显著高于绿叶(P<0.01)。

表1 三个时期的黄叶和绿叶银杏光合色素含量
2.2 叶绿体超微结构的比较
如图2a所示,在4月底,黄叶和绿叶叶绿体迅速增大,细胞和叶绿体大小相似,并开始出现少量嗜锇颗粒;黄叶形成的中央大液泡较绿叶小,片层结构较绿叶排列疏松;黄叶叶绿体中开始大量出现淀粉粒,但较绿叶小。如图2b所示,在5月底黄叶和绿叶细胞结构类似,叶绿体形态没有明显差异,均呈现为椭圆形,淀粉粒较大,二者数量和形态没有明显差异,存在少量嗜锇颗粒。如图2c所示,8月时,黄叶和绿叶叶绿体内淀粉粒大量减少,嗜锇颗粒明显增多,片层结构变得疏松或不清晰,叶片开始衰老;黄叶叶绿体形状较绿叶变得不规则,内囊体片层结构减少程度较绿叶严重,嗜锇颗粒较多,说明黄叶的类囊体降解早于绿叶,衰老时间早。

a
2.3 光合气体交换参数的比较
如表2所示,3个时期的黄叶和绿叶的净光合速率(Pn)均呈现升高-降低的变化趋势,其中5月的Pn极显著高于4月和8月(P<0.01);4月和8月的黄叶Pn显著或极显著低于绿叶(P4月<0.05和P8月<0.01),而5月的黄叶Pn均显著高于绿叶(P<0.05)。3个时期黄叶和绿叶气孔导度(Gs)和胞间二氧化碳浓度(Ci)均呈升高的趋势,且不同月份间差异极显著(P<0.01);4月和5月黄叶Gs与绿叶差异均不显著(P>0.05),8月黄叶Gs极显著低于绿叶(P<0.01),同期黄叶的Ci与绿叶差异不显著(P>0.05)。3个时期黄叶和绿叶蒸腾速率(E)均呈“降低-升高”的趋势,且不同月份间差异极显著(P<0.01);4月和8月黄叶E显著或极显著低于绿叶差异(P4月<0.05和P8月<0.01),5月黄叶E与绿叶差异不显著(P>0.05)。黄叶和绿叶瞬时水分利用效率(WUE)均在5月达到最高水平,分别为16.52和14.93 μmol/mmol,并在8月降至最低,分别为1.37和1.65 μmol/mmol;3个时期黄叶WUE均与绿叶差异不显著(P>0.05)。3个时期的黄叶和绿叶瞬时羧化效率(CE)和气孔限制值(Ls)均呈下降的变化趋势,3个时期黄叶CE和Ls均与绿叶差异不显著(P>0.05)。
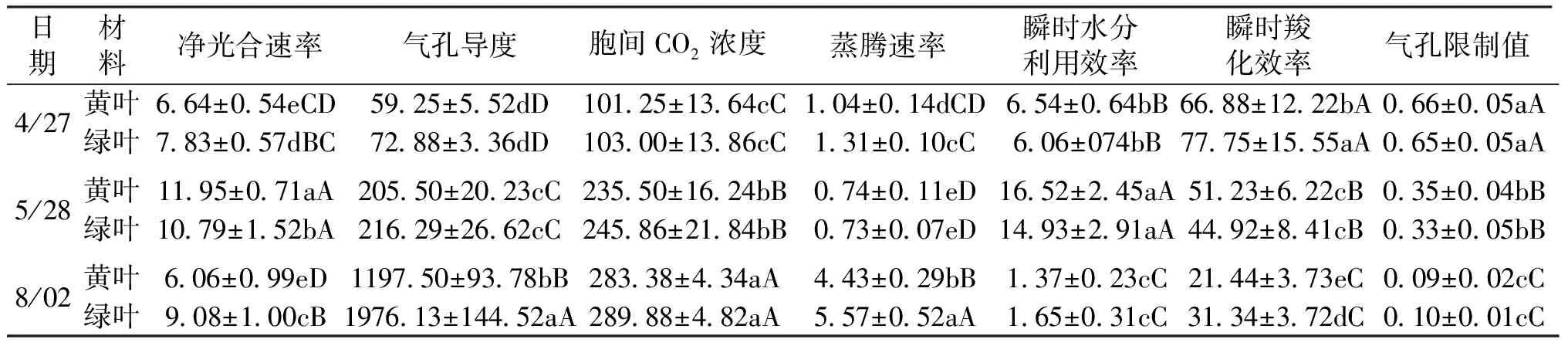
表2 黄叶和绿叶光合特性的比较
2.4 叶绿素荧光动力学参数的变化
如表3所示,黄叶的Fo、Fo′、Fm、Fm′、Fs和ETR在3个时期均极显著低于绿叶(P<0.01),这与黄叶叶绿素色素含量显著低于绿叶的结果一致。4月时,黄叶的Fv/Fm和NPQ显著高于绿叶(P<0.01),表明其ΦPS II有较高的光能转换效率,而且此时其天线色素吸收的光能以热形式耗散较多;黄叶的Fv′/Fm′和φPS II显著低于绿叶,说明其φPS II天线色素吸收光能并用于光合电子传递的量子产额较小,φPSII原初光能捕获效率较低;二者qP差异不显著,说明二者的电子传递活性相似。5月时,黄叶的φPS II和qP显著低于绿叶(P<0.01),说明此时黄叶的φPS II天线色素吸收光能并用于光合电子传递的量子产额较小,其电子传递活性明显低于绿叶;黄叶的Fv/Fm显著高于绿叶(P<0.05),Fv′/Fm′和NPQ与绿叶差异不显著,说明此时其φPS II原初光能捕获效率和天线色素吸收光能的热耗散基本一致。8月时,除Fo、Fo′、Fm、Fm′、Fs和ETR 6个指标外,黄叶的Fv/Fm、Fv′/Fm′、NPQ、φPS II和qP与绿叶差异均不显著,说明此时二者的ΦPS II活性并没有明显的差异。
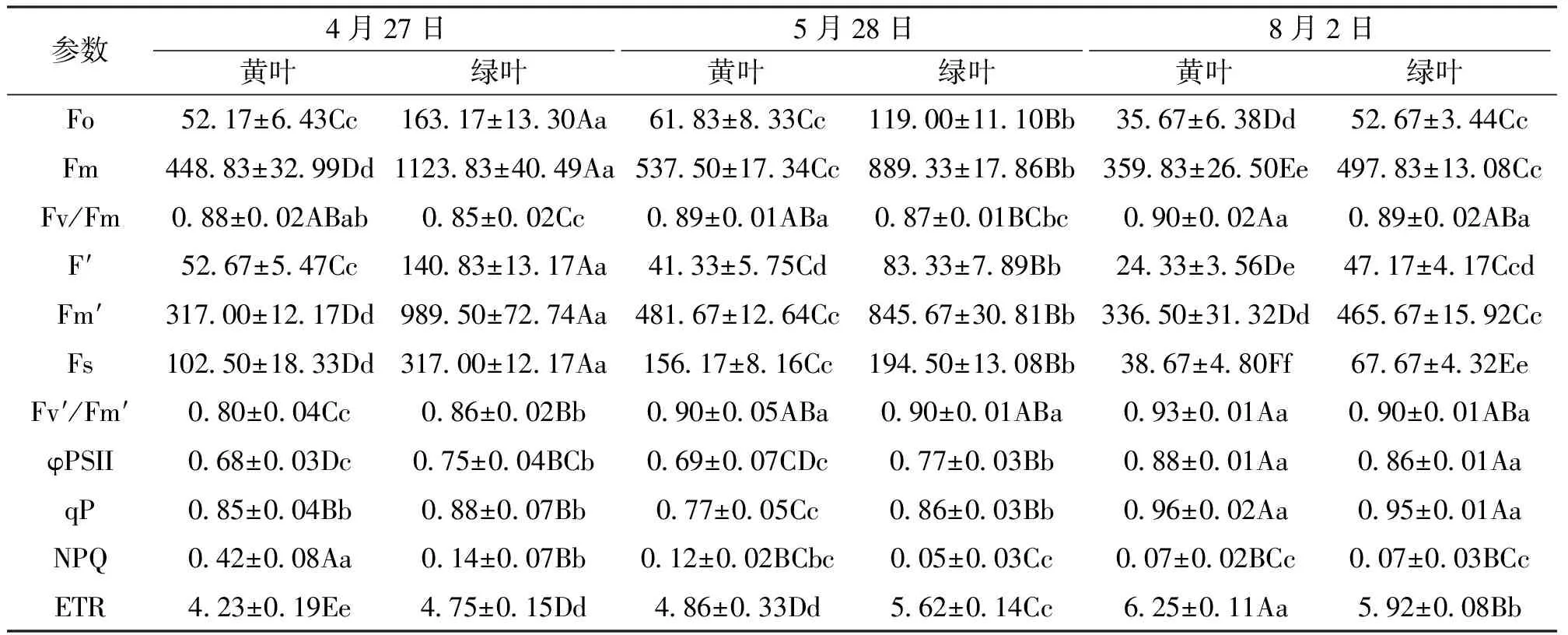
表3 黄叶和绿叶叶绿素荧光动力学参数的比较
2.5 叶片吸收光能分配的差异
如表4所示,4月时,黄叶的P显著低于绿叶(P<0.01),说明此时黄叶吸收的光能用在光化学反应上的比例小于绿叶;随着叶片的发育,黄叶的P开始增加,至8月时,与绿叶处于相同的水平。4月黄叶的D显著高于绿叶,说明此时黄叶吸收的光能在捕光天线热耗散的比例较高;5月时二者处于相同水平,8月时黄叶显著低于绿叶(P<0.01)。4月和8月黄叶的Ex与绿叶处于同一水平上。5月时黄叶显著高于绿叶(P<0.01),二者Ex水平均显著增加,黄叶的增加幅度高于绿叶。这说明5月时黄叶光合机构中的过剩光能高于绿叶,由于这部分光能没有被光化学反应利用和捕光天线耗散,可能导致体内活性氧含量增加,从而造成光合机构的破坏。

表4 3个时期黄叶和绿叶光能吸收与分配
3 讨论与结论
植物叶色突变体叶色变异的直接原因是叶中色素含量、比例或分布发生了改变[19-21]。本研究发现,3个时期芽变银杏叶中叶绿素含量均极显著低于绿叶(P<0.01),推测该突变为总叶绿素缺乏型。最佳观赏期(4月27日)黄叶的Chl含量显著低于绿叶(P<0.01),仅为绿叶的21.27%,且Chla/b(4.914)显著高于绿叶(1.365),Car含量与绿叶差异不显著(P>0.05),但Car/Chl(0.365)极显著高于绿叶(0.088)(P<0.01);8月黄叶Car/Chl(0.233)仍显著高于绿叶(0.033);黄叶Car含量比4月黄叶显著升高,而8月绿叶中Car含量比4月底绿叶显著降低。综上可知,黄叶银杏黄化的主要原因是由叶绿素含量及其所占比例显著降低所致,而与类胡萝卜素含量关系不大,与潘晶晶等[22]、李艺迪等[23]、潘晶晶[24]的研究结果一致。
由于叶绿素主要结合在类囊体膜上,叶绿素的生物合成与类囊体膜的建成密切相关,与叶绿体的发育程度有直接的关系[25]。研究表明,叶色突变会导致叶绿体发育异常,主要表现为叶绿体发育不完全,结构表现不正常,如叶绿体形状不规则,片层结构较少、较薄(疏松)、排列不规则,类囊体表现为不同程度的膨胀,含有较少的淀粉粒等。本研究黄叶叶绿体发育和类囊体膜结构建成明显晚于绿叶,即黄叶叶绿体的发育滞后,使黄叶中叶绿体数和叶绿体片层数减少,导致叶绿素合成受限,含量降低,使叶片黄化。叶绿体基质中嗜锇颗粒的出现通常被认为是脂质物质积累的标志,当叶绿体在衰老阶段时,嗜锇颗粒的增大是片层结构开始解体的重要标志。黄叶叶绿体在8月出现较多的嗜锇颗粒,表明其衰老的开始时期较绿叶早。总之,根据叶绿体结构分析,黄叶叶绿体内部结构发育缺陷,使叶绿素合成代谢基础较弱,导致叶绿素含量降低,造成其光合能力较弱。
目前对叶色突变体光合特性研究较多,叶绿素缺乏对突变体光合能力的影响不尽相同。在一般情况下,突变体内叶绿素含量降低导致其光合能力降低,但净光合速率与叶绿素含量之间很少显著性相关[7,26]。此外,由于光合作用是在叶绿体中进行的复杂的生物过程,而且光合机构对光能的吸收和转化主要发生在类囊体膜上,因此叶绿体的发育程度直接决定了植株光合能力的大小[27]。本研究中Pn与叶绿素含量的变化趋势并不相同,其差异程度远小于叶绿素含量的差异,而且黄叶5月的Pn显著高于绿叶,说明叶绿素含量在超出一定范围后,并不是Pn高低的决定因素。结合之前叶绿体超微结构的观察结果,黄叶叶绿体内部结构发育缺陷,是造成其光合能力较弱的主要原因。
一般认为,叶色突变减少了捕光色素蛋白复合体的含量,因而影响到光系统Ⅱ供体侧的稳定性,使突变体对光强和高温的耐受性比野生型低[28]。本研究发现,黄叶的Fo、Fo′、Fm、Fm′和Fs在各个时期均显著低于绿叶,与叶绿素含量显著降低的现象一致。4月底时,黄叶的Pn、Gs、E、CE均显著低于绿叶,而Ci和WUE与绿叶处于相同水平上,这是由于此时黄叶叶绿素含量水平较低和叶绿体发育不完全引起的;叶绿素荧光参数Fo、Fo′、Fm、Fm′、Fs、φPS II、Fv′/Fm′和ETR,但是此时Fv/Fm和NPQ显著高于绿叶(P<0.01),说明黄叶PS II反应中心内有较高的光能转换效率和热耗散。黄叶中高比例的叶绿素a有利于提高其PSII光化学效率,较高的光能转换效率是其Pn与绿叶差异不大的原因(为绿叶的0.85倍),是对低叶绿素水平限制光能捕获的适应性调节作用。植物光合速率降低的因素有气孔限制因素和非气孔限制因素,只有Pn与Ci同时下降且Ls同时增加时,才能判断气孔限制因素是光合速率下降的主要原因。虽然黄叶在8月时的Pn和Gs值显著低于绿叶,但其Ci、Ls并没有显著差异,因此可以判断黄叶Pn的下降是由非气孔因素引起的。8月时,黄叶和绿叶的Fv/Fm、Fv′/Fm′、φPS II、qP、P等参数均无显著差异,说明此时黄叶和绿叶PS II反应中心活性相似,反应中心吸收的光能用在光化学反应和耗散上的比例相似。
综上所述,芽变黄化银杏叶极低的叶绿素含量及其占比和叶绿体发育不健全是其黄化呈色的物质基础和结构基础,并影响了光系统发育、光能吸收与捕获、电子传递、激发能分配等活动,具体表现为降低了光合能力、光能吸收与捕获、电子传递速率,提高了热耗散水平和原初光能转换效率。在4月黄叶有较高的热耗散,对光合机构起到一定的保护作用,从而维持正常的光合作用;同时有较高水平的光能转换效率;8月黄叶和绿叶净光合速率主要受非气孔因素限制,且黄叶Pn受高温的影响较绿叶严重,叶绿体开始衰老。植物叶色的突变是生理和遗传因素共同作用的结果,关于黄叶叶色变异的分子机制有待进一步深入研究。
