毛竹对锰胁迫土壤的响应研究
2023-08-17蔡启航罗学刚
蔡启航,张 宇,罗学刚,
(1.西南科技大学生命科学与工程学院,四川 绵阳 621000;2.西南科技大学材料学教育部工程研究中心,四川 绵阳 621000)
【研究意义】锰(Mn)是动植物生长代谢中必需的微量营养元素之一,通常在土壤和水中的浓度较低,一般不被认为是潜在的有毒元素[1]。过去的采矿、加工业和农业等人类活动,破坏了Mn 含量的平衡,导致土壤、沉积物和水体中的Mn 污染增加[2]。研究表明,Mn 会对人体产生长期且隐性的危害,包括帕金森样症状以及生殖系统异常,也对免疫系统产生一定影响[3-4],Mn污染土壤修复已成为当前重要的环境问题[5]。传统的土壤修复方法主要为物理化学方法,如空气氧化法、氯氧化法和接触氧化法等[6-9],这些方法价格昂贵且易产生二次污染[6-7]。植物修复是一种经济、环保的修复方法,已被广泛用于治理重金属污染土壤。植物提取是植物修复技术最常用的方法,利用植物将土壤中的重金属元素富集到植物地上部分,再通过收割地上部分进行处理,能减少或除去土壤中的重金属污染物[8-9]。而植物修复污染土壤的关键在于筛选生物量大、生长周期短且重金属富集量高的新品种[10]。
【前人研究进展】目前,研究人员已对耐受Mn 的植物进行了广泛研究,取得良好效果[17]。研究表明,苍耳(Xanthium sibiricumPatrin ex Widder)等草本植物在污染土壤中显示出较强的Mn 吸收能力,但其生物量低、生长速度慢、根系浅,不适合大规模修复[11]。毛竹(Phyllostachys pubescens)是一种重要的经济竹种,我国种植面积超过527.76 万hm2,占全国竹子总面积的70%[12-13]。毛竹具有环境适应强、生长速度快、用途多、生态价值高等特性,地上部分生物量占全竹 90%以上,且根系能在地下远距离蔓延生长,具有吸附重金属的先天优势。水培和盆栽试验表明,毛竹在高质量浓度重金属胁迫下无明显的坏死和黄化等中毒症状[14]。张玲等[15]研究发现,生长在铅(Pb)、锌(Zn)尾矿周边的毛竹芽中Pb 含量高达1 182 mg/kg。Zhang 等[16]在某废弃矿区发现生长良好的毛竹人工林,矿区土壤Pb含量高达286 mg/kg,表明毛竹对Pb 胁迫具有较强的耐受能力。【本研究切入点】目前关于毛竹植物的研究大多在水培或盆栽试验条件进行,且只针对Pb、Zn 等重金属元素,对Mn 胁迫土壤的相关研究较少,无法充分了解毛竹对Mn 胁迫土壤的修复效果。【拟解决的关键问题】本文探究标准土柱内Mn 胁迫土壤3 年生毛竹的生理响应,为Mn 胁迫土壤的治理提供一定理论参考,为重金属污染土壤的修复提供新的植物资源。
1 材料与方法
1.1 试验材料
毛竹苗购自四川洪雅县廷富苗木有限公司。
1.2 试验方法
试验于2018 年6 月开始,通过五点取样法采 自西南科技大学周边农田土作为供试土壤,土壤粘性30.81%,质地为中壤。利用便携式X 射线荧光光谱(PXRF)技术[17]测定土壤中Cr、Mn、Fe、Ni、Cu 和Zn 含量分别为85.4、538.9、755.9、58.8、26.3、423.3 mg/kg,超出当地土壤背景值,本研究主要分析毛竹对上述6 种重金属的富集特性。毛竹(Phyllostachys pubescens)种植于西南科技大学试验田长、宽、深均为1 m 的土柱内,每个土柱装约1 000 kg 土壤(四川绵阳,海拔500 m,年均气温14.7~17.3 ℃,年降水量825.8~1 417 mm)。土壤设置3 个处理(Mn 浓度分别为1 000、2 000、4 000 mg/kg),以不添加Mn 的土壤为对照,每个处理3 次重复,每个重复面积为1 m2,采用五点法栽种5 株毛竹实生苗,自然光照和雨水灌溉,3 年后采样。
1.3 毛竹生长特性测定
1.3.1采样与前处理 2021 年7 月,每个处理选取长势一致的3 株毛竹,采集毛竹中部(从下往上数6~7 节间)的茎与侧枝条发育较好的叶片,在土壤深度40 cm 处采集根系。清理叶片和茎上杂物、根系上的土壤,放入液氮速冻,置于-80oC 冰箱保存,备用。
1.3.2表观数据 每组分别选取长势一致、叶片大小均一、生长状况相同的3 株毛竹进行测定。叶长、叶宽、株高用卷尺测量。利用游标卡尺测量竹基部外径。每个处理5 次重复。
1.3.3纤维形态 利用LDA02 型纤维质量浓度分析仪测定毛竹纤维形态[18]。选取部分茎杆切成火柴杆大小,置于10 mL 离心管中,使用离析液(10%硝酸与10%铬酸)浸没样品,室温放置2 d。将离析后的样品置于玻璃皿中,用清水洗至无色后进行测定。每个处理3 次重复,每个重复3 个样品,每个样品测定5 000 根纤维。
1.3.4毛竹重金属元素累积量 利用破碎机将各组干燥后的根、茎和叶粉碎,通过微波消化法分别提取根、茎和叶中的元素。在微波反应系统中将10 mL 硝酸与12 mL 过氧化氢的混合物对0.100 g 样品进行消解。微波消解方法:升温10 min,120 ℃加热5 min;升温10 min,150 ℃加热10 min;升温10 min,180 ℃加热30 min;待消解管冷却后,通过加热去酸。用硝酸溶液(体积分数为1%)冲洗3 次,并将溶液转移到50 mL 容量瓶中 。提取物通过0.45 μm 滤膜过滤,并在提取后3 d 内采用电感耦合等离子体质谱法测定元素质量浓度。每个处理3 次重复。
1.4 毛竹光合参数与叶绿素参数测定
1.4.1光合参数 毛竹的光合参数利用Li-6400 系列便携式光合作用测定仪测定[19]。包括净光合速率(A,mmol H2O/m2·s)、蒸腾速率(E,mmol H2O/m2·s)、胞间CO2浓度(Ci,ppm)、水分利用率(WUE,g/kg)、叶片温度(Tleaf,℃)和叶片水汽压亏缺(VpdL,kPa)。每个处理9 次重复。
1.4.2叶绿素含量 采用乙醇丙酮(3∶1,V/V)萃取法测定叶绿素含量[20]。称取毛竹叶片0.1 g,加入5 mL色素提取物,快速研磨和浸出5 h(25 ℃)后,用分光光度计分别在645、663 nm 波长下测量。每个处理5 次重复。
1.4.3叶绿素荧光参数 叶绿素荧光参数利用MIN-PAM 仪器(Walz,Inc,德国)测定[21]。叶绿素荧光参数包括最大荧光产量(Fm)、可变荧光(Fv)、PS II的最大光化学量子产率(Fv/Fm)、叶绿素荧光指标Y(II)、相对电子传递速率(ETR)、光合有效辐射(PAR)、光化学淬灭系数(qP)和非光化学淬灭系数(qN)。每个处理9 次重复。
1.5 抗氧化代谢酶 活性与丙二醛含量测定
使用试剂盒(Solarbio Science &Technology Ltd.,北京)测量抗氧化酶活性与丙二醛(MDA)含量。抗氧化酶包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)。每个处理5 次重复。
1.6 统计分析
利用OmicShare工具对数据进行标准化处理,对毛竹代谢的6 种元素(Mn、Fe、Ni、Cr、Zn和Cu)进行主成分分析(PCA)和权重网络分析。选择权重在0.5~1.0 之间的元素揭示元素间的相互作用。利用SPSS 版本20.0 软件计算数据的平均值与标准偏差,进行单因素方差分析,多重比较采用邓肯检验。转移系数(TF)为地上部的重金属质量浓度除以根的重金属质量浓度。
2 结果与分析
2.1 土壤中 不同浓度锰胁迫对毛竹植物学性状的影响
如图1、图2 所示,与对照相比,在土壤4 000 mg/kg Mn 胁迫浓度下,毛竹的株高、叶长、叶宽和基径分别下降22.6%、25.9%、21.3%和26.7%,毛竹的生长发育受到抑制。

图1 土壤中不同浓度锰胁迫下的毛竹生长形态Fig.1 Growth morphology of moso bamboo under different concentrations of Mn stress in soil
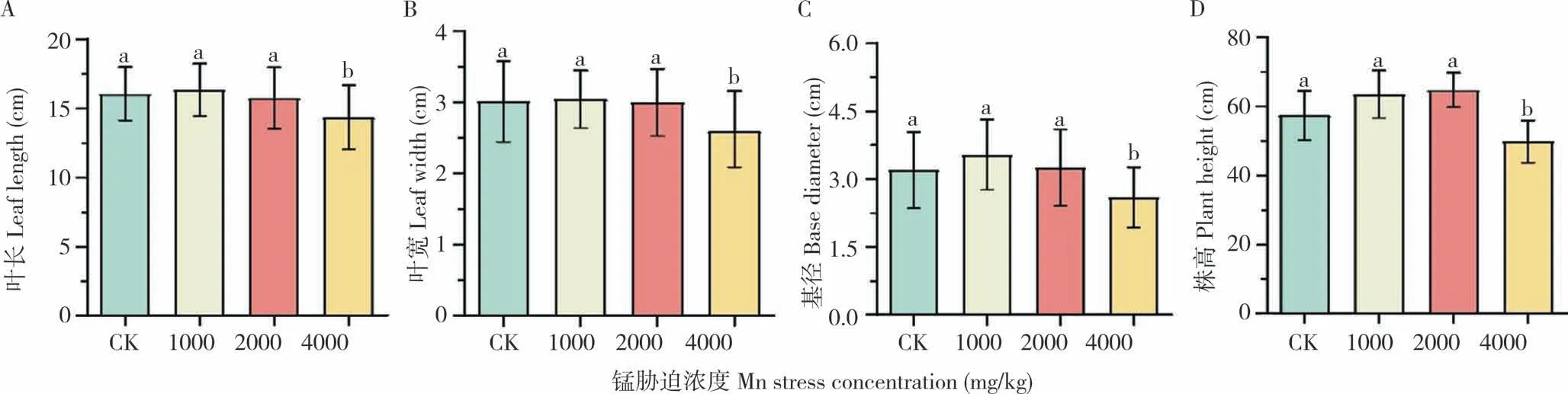
图2 土壤中不同浓度锰胁迫对毛竹生长指标的影响Fig.2 Effects of different concentrations of Mn stress in soil on growth index of moso bamboo
2.2 土壤中不同浓度锰胁迫对毛竹纤维形态的影响
如表1 所示,与对照相比,在土壤2 000 mg/kg Mn 胁迫浓度下,毛竹的细小纤维长度、宽度分别增加15.6%、20.6%,在2 000、4 000 mg/kg Mn 胁迫浓度下,纤维的扭结指数分别增加26.6%、47.5%,在1 000 mg/kg Mn 胁迫浓度下,毛竹的扭结角度和每毫米扭结数分别降低22.3%、21.5%,对毛竹纤维的平均长度和加权平均长度无显著影响。

表1 土壤中不同浓度锰胁迫下的毛竹纤维形态Table1 Fiber morphology of moso bamboo under different concentrations of Mn stress in soil
2.3 土壤中不同浓度Mn 胁迫对毛竹重金属富集特性的影响
2.3.1毛竹地上部与地下部对重金属的富集特性 如图3 所示,随着Mn 胁迫浓度的增加,毛竹地上部与地下部对Mn 的吸收量逐渐增大。在0、1 000、2 000 和4 000 mg/kg Mn 胁迫浓度下,毛竹对Mn 的平均富集量分别为1 098.6、2 336.4、3 530.9、5 370.4 mg/kg,对Mn 的转移系数分别为1.59、3.83、3.56 和4.17(表2),表明毛竹对土壤中的Mn 有较强的富集能力。与对照相比,在土壤4 000 mg/kg Mn 胁迫浓度下,Fe、Ni、Cr 和Zn 的转移系数分别降低38.1%、17.9%、28.5%、47.8%。表明毛竹对各类重金属有不同的富集特性,高浓度Mn 胁迫阻碍重金属元素向毛竹地上部转移。

表2 土壤中不同浓度锰胁迫下毛竹各重金属转移系数Table 2 Transfer coefficients of heavy metals in moso bamboo under different concentrations of Mn stress in soil
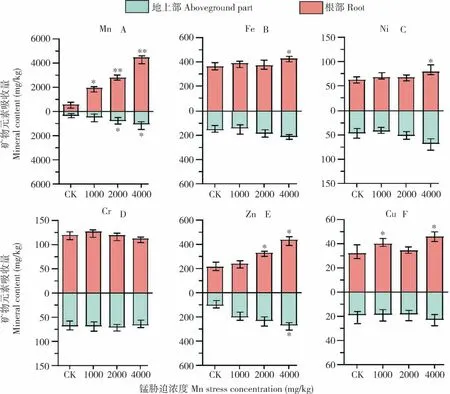
图3 土 壤中不同浓度锰胁迫对毛竹地上部与根部重金属吸收量的影响Fig.3 Effects of different concentrations of Mn stress in soil on the absorption of heavy metals in the aboveground parts and root of moso bamboo
2.3.2矿物元素的主成分分析和加权网络分析 为阐明Mn 与其他元素的相互作用,对毛竹矿物元素进行主成分分析和加权网络分析。由图4可知,不同浓度Mn 胁迫下毛竹的矿物元素簇状位置差异显著,表明高浓度的Mn 胁迫对毛竹的矿物代谢有显著干扰。Mn与Ni、Cu和Fe呈正相关关系,与Cr 和 Zn 呈现负相关关系,与Fe 的相关性最强。可见,高浓度的Mn 胁迫导致毛竹的矿物元素代谢发生变化,是生长异常的主要原因之一。

图4 毛竹重金属元素代谢的主成分(A)和加权网络(B)分析Fig.4 Principal component (A) and weighted network(B) analysis of heavy metal element metabolism in moso bamboo
2.4 土壤中不同浓度Mn 胁迫对毛竹光合参数和叶绿素含量的影响
由图5 可知,与对照相比,不同浓度Mn 胁迫对毛竹净光合速率的蒸腾速率、胞间CO2浓度、叶片温度和叶片水汽压亏缺无显著影响,而水分利用率显著降低30.6%,高浓度的Mn 胁迫对毛竹的光合色素影响不显著。

图5 土壤中不同浓度锰胁迫对毛竹光合参数的影响Fig.5 Effects of different concentrations of Mn stress in soil on photosynthetic parameters of moso bamboo
2.5 土壤中不同浓度锰胁迫对毛竹叶绿素荧光参数的影响
如图6 所示,不同浓度Mn 胁迫对毛竹叶片的叶绿素荧光参数影响显著。与对照相比,在1 000 mg/kg Mn 胁迫下,最大荧光产量、可变荧光量显著降低11.6%、16.9%。2 000、4 000 mg/kg Mn 胁迫下的光化学淬灭指数分别下降19.8%和29.8%,而非化学淬灭指数分别下降23.5%和20.7%。PS II 的最大光化学量子产率、叶绿素荧光指标、光合有效辐射和相对电子传递速率没有显著变化 。表明高浓度Mn 胁迫使光化学淬灭能力和非光化学淬灭能力受到抑制,光合效率降低。

图6 土壤中不同浓度锰胁迫对毛竹叶绿素荧光参数的影响Fig.6 Effects of different concentrations of Mn stress in soil on chlorophyll fluorescence parameters of moso bamboo
2.6 土壤中不同浓度锰胁迫对毛竹抗氧化代谢酶活性与丙二醛含量的影响
如表3 所示,与对 照相比,1 000 mg/kg Mn胁迫浓度下,毛竹的SOD 和CAT 活性显著增加37.8%、18.4%;2 000 mg/kg Mn 胁迫浓度下,SOD 活性增加15.2%,而POD 活性下降31.5%;在4 000 mg/kg Mn 胁迫浓度下,POD 与CAT 活性分别下降39.4%、22.6%。不同浓度Mn 胁迫对毛竹的抗氧化代谢酶的活性影响不同,对MDA含量无显著影响。

表3 土壤中不同浓度锰胁迫下的毛竹抗氧化酶活性(U/mg)和丙二醛含量(nmol/g)Table 3 Antioxidant enzyme activity and malondialdehyde content under different concentrations of Mn stress in soil
3 讨论
Mn 在植物代谢和生长过程中起关键作用[22],参与光合作用、呼吸作用,苹果酸酶、硝酸还原酶等酶的生物合成,也是三羧酸循环中许多酶的激活剂[23]。研究表明,超过一定浓度范围,重金属会对植物的生长发育产生危害。重金属污染通过取代必需元素破坏电子传递链、细胞水分失衡、光合色素减少、胞内CO2含量的减少、叶绿素含量下降等[24]。毛竹作为具有一定修复能力的植物,具有调节机制,保护光合作用免受重金属诱导的胁迫,如信号通路、运输网络、活性氧清除、叶绿素相互转化、基因表达等[25]。
毛竹是一种生长快速的植物,高浓度Mn 胁迫对毛竹的矿物代谢有显著干扰,损伤毛竹的根系、抑制毛竹叶片的水分利用能力,降低毛竹的生物学指标与生物量等。与对照相比,4 000 mg/kg Mn 胁迫浓度下,毛竹的株高、叶长、叶宽和基径分别下降22.6%、25.9%、21.3%和26.7%。在实际应用中,在毛竹发育早期可利用分枝、分层、扦插或枝条扦插的无性繁殖等方法适应重金属胁迫[26]。
细胞质膜是植物受到重金属胁迫时的第一道保护屏障,当重金属浓度超过植物耐受范围时,会造成氧化应激和损害,出现脂质过氧化现象。本研究中,与对照相比,4 000 mg/kg Mn 胁迫下,POD 活性与CAT 活性分别下降39.4%、22.6%,对MDA 含量没有显著影响。表明不同浓度Mn 胁迫对毛竹的抗氧化代谢酶的活性影响显著,但对毛竹的膜系统的过氧化损伤有限。毛竹的存活率与生长状况较好,没有叶片枯死黄化、茎秆干枯损伤等现象,说明毛竹具有很强的重金属耐受性。Wang 等[27]研究发现,水培试验中,Mn 胁迫下栾树叶片光合作用下降,导致植物生物量和叶绿素含量显著下降。
与对照相比,在4 000 mg/kgMn 胁迫浓度下的光化学淬灭指数下降29.8%,而非化学淬灭指数下降20.7%,表明高浓度Mn 胁迫抑制毛竹光合作用的效率。光合作用期间产生的能量会用于防御重金属的诱导产生的氧化应激,以维持自身的生长发育[28]。在后续应用中可以调控毛竹的生长条件,如增加光照强度、增加CO2浓度、保持土壤水分等,用于增加毛竹的生物量,使之达到预期的修复效果[29]。
本研究中,毛竹虽没有达到超富集植物的标准,但在4 000 mg/kgMn 胁迫浓度下,对Mn 的转移系数为4.17,表明毛竹具有较强的富集能力[30]。而毛竹纤维形态数据表明,Mn 胁迫不会影响毛竹的纤维长度,修复后毛竹依旧能作为纸浆加工的原材料。今后,我们将进一步从生理层面深入研究毛竹对Mn 的耐受性,探讨重金属在毛竹体内的转移机制,揭示毛竹对重金属的解毒机理。可以将毛竹与其他超富集植物联合,修复更多的目标重金属土壤,提升空间利用率。还可探究在添加EDTA、槐糖脂等外源物质后毛竹对重金属污染的响应机制,为实现植物修复研究领域提供新的思路[31-33]。
4 结论
本文以毛竹为研究对象,进行3 年Mn 胁迫土柱试验,对毛竹植物学性状、光合参数、叶绿素荧光参数等指标进行测定,阐明毛竹在不同浓度Mn 胁迫下的毒性响应机制。结果表明,高浓度的Mn 胁迫对毛竹的植物学性状指标有一定抑制作用,显著降低毛竹光化学淬灭能力和非光化学淬灭能力,影响毛竹的相关抗氧化代谢酶的活性。但在4 000 mg/kgMn 胁迫下毛竹依旧生长,没有叶片枯死黄化等现象,对Mn 的转移系数达到4.17,对Mn 胁迫土壤有很好的富集作用。毛竹易于栽培、生物量大、环境适应能力较强,可作为一种新的Mn 胁迫土壤的植物修复品种。该研究为今后重金属污染治理的应用与植物修复领域的研究提供参考。
