新疆牛羊源金黄色葡萄球菌D353质粒pD353序列分析
2023-07-28贺腾飞刘英玉张柳青李泽亚蒋金豆祖力胡马尔艾力
贺腾飞,刘英玉,张柳青,陈 旺,李泽亚,胡 芸,蒋金豆,祖力胡马尔·艾力
(新疆农业大学动物医学学院,乌鲁木齐 830052)
0 引 言
【研究意义】质粒是染色体外的遗传物质,携带遗传信息,并能够通过接合的方式有效进行遗传信息的水平转移,进而增强细菌对不同环境的适应能力[1]。耐甲氧西林金黄色葡萄球菌(methicillin-resistantStaphylococcusaureus,MRSA)及耐万古霉素金黄色葡萄球菌(Vancomycin resistantStaphylococcusaureus,VRSA)的出现是有危害的。在当前具有感染性的金黄色葡萄球菌菌株中,抗生素的耐药性通常是由质粒编码的,这些质粒可以通过水平DNA转移机制在菌株之间传播。临床分离的金黄色葡萄球菌和凝固酶阴性葡萄球菌(coagulase negative staphylococci,CNS)通常携带一个或多个耐药质粒,这些质粒是基因转移的重要载体,有助于葡萄球菌耐药关键基因的获得、维持和传播。金黄色葡萄球菌D353对苯唑西林、万古霉素、红霉素、克拉霉素和诺氟沙星等12种抗生素耐药,类似这种耐多重抗生素菌株的出现和水平传播增加了人类治疗人畜共患疾病的困难,深入研究金黄色葡萄球菌质粒的复制方式、携带耐药基因及结构特点,对于揭示金黄色葡萄球菌对环境的适应机制、监测金黄色葡萄球菌的流行性并干预其质粒传播具有重要意义[2-4]。【前人研究进展】常见的有小滚圈复制的葡萄球菌质粒(1-5 kb),其生物学特性已经被详细分析,但除了它们携带抗性基因外,Theta复制质粒也携带抗性基因且片段较大,但对这种质粒的关注相对较少。目前已在葡萄球菌中识别出三类Theta复制质粒,即pSK639家族、多耐药质粒(重金属/β-内酰胺酶质粒和pSK1家族)和接合多耐药质粒(pSK41家族)[5]。了解质粒复制的详细分子机制可能会发现用于逆转或减缓耐药性发展的新方法[6]。【本研究切入点】在前期研究中,发现新疆牛羊源的金黄色葡萄球菌存在大量的耐药菌株和一些MRSA菌株[7],其中D353菌株对于苯唑西林、诺氟沙星、克拉霉素、红霉素和万古霉素等12种抗生素耐药。分析金黄色葡萄球菌耐甲氧西林菌株和耐万古霉素菌株的野生型质粒序列特征。【拟解决的关键问题】提取耐药菌株质粒并测序,分析质粒的结构和同源性,解释质粒复制转移能力和自身特性,对新疆牛羊源金黄色葡萄球菌质粒数据库提供理论依据。
1 材料与方法
1.1 材 料
金黄色葡萄球菌D353[7],来源于新疆库尔勒牛羊肉农贸市场的工具拭子,样品获取时间为2018年10月,菌液与60%甘油1∶ 1,-20℃保存于新疆农业大学畜产品质量安全实验室。
质粒提取试剂盒(TIANGEN公司);溶菌酶(Solarbio公司);7.5% NaCl 肉汤(青岛海博生物技术公司);Baird-Parker琼脂基础(青岛海博生物技术公司);10×Loading Buffer(TaKaRa公司);1 kb Ladder(康为世纪公司)。
生物安全柜(美国LABCONCO公司);高速离心机D2012 plus(SCILOGEX公司);恒温振荡器(上海一恒科学仪器有限公司);电热恒温培养箱(上海一恒科学仪器有限公司);微量分光光度计SMA4000(Merinton 仪器公司);梯度PCR仪(美国伯乐公司)。
1.2 方 法
1.2.1 菌株培养
将实验室保存的金黄色葡萄球菌D353和7.5% NaCl 肉汤液体培养基按照1∶ 9混合,放入恒温振荡器,150 r/min 培养18 h ,划线接种 Baird-Parker 平板,放入电热恒温培养箱37℃培养24 h。
1.2.2 菌株16S rRNA鉴定
取1 mL菌液提取基因组DNA,16S rRNA的扩增引物序列为F:AGAGTTTGATCCTGGCTCAG,R:ACGGCTACCTTGTTACGACTT,扩增反应条件为94℃预变性5 min,94℃变性30 s,55℃退火30 s,72℃延伸90 s,30个循环,72℃再延伸5 min,4℃保存,将PCR产物送上海生物工程有限公司测序,鉴定菌株类别。
1.2.3 质粒提取
按照质粒提取试剂盒方法提取质粒,取1~5 mL菌液离心,去上清,先用溶菌酶100 mg/mL,37℃,20 min恒温处理菌体,按照试剂盒说明书提取质粒。
1.2.4 质粒浓度
用微量分光光度计测定提取的质粒原液浓度,获得质粒浓度。
1.2.5 质粒测序与组装
将提取的质粒送至西安擎科生物技术公司测序,采用Miseq测序法,之后利用 plasmidid(version 1.6.5)进行组装。
1.2.6 生物信息学
利用NCBI网站上的ORF Finder寻找质粒的开放阅读框(open reading frame,ORF)以及BLAST进行初步注释,利用Snap Gene(version 4.2.4)制作质粒图谱,使用MEGA 11软件制作16s rRNA序列和质粒复制起始蛋白repB的Neighbor-Joining进化树,并利用Mauve软件进一步分析几株质粒间的亲缘关系。
2 结果与分析
2.1 菌株16S rRNA鉴定结果
研究表明,以葡萄球菌属S91和表皮葡萄球菌SW-7/FSE的16S rRNA基因为参照,建立ML树。分析可知,D353与金黄色葡萄球菌 NRLFFD288以及 B0021-01R亲缘关系较近,且与金黄色葡萄球菌DFI-23聚为一支。确定菌株D353为金黄色葡萄球菌。图1

图1 质粒pD353 16S rRNA系统发育树
2.2 质粒浓度和序列
研究表明,利用微量分光光度计测定pD353浓度为40.29 ng/μL,将pD353测序拼接序列上传NCBI的Gene Bank数据库,序列号为NZ_CP080004,利用NCBI网站上的BLAST分析,其中质粒pSaD592、pIT1-S和pIT4-R与pD353同属于金黄色葡萄球菌质粒,而质粒pfas4-2和pSF2来自于海氏肠球菌和粪肠球菌。
质粒pD353长度34063 bp,其(G+C)mol含量为30.42%。pD353质粒有47个超过200个核苷酸的ORF。ORF110编码314 aa的复制蛋白repA,repA含有复制起始蛋白A的N端结构域属于超家族cl21459和复制起始蛋白A的C 端结构域是超家族cl39431。ORF139编码286 aa的复制蛋白repB属于COG5527超家族结构域。ORF41编码一个类似于IS21元件的ISBmu3家族转座酶istA基因,ORF13编码一个类似于IS21元件的 IS5376 家族辅助ATPaseistB基因。在pD353发现了5种转座酶,分别是IS3家族转座酶、类IS6 ISSau6家族转座酶、IS21家族转座酶、IS30家族转座酶和IS1182家族转座酶。图2
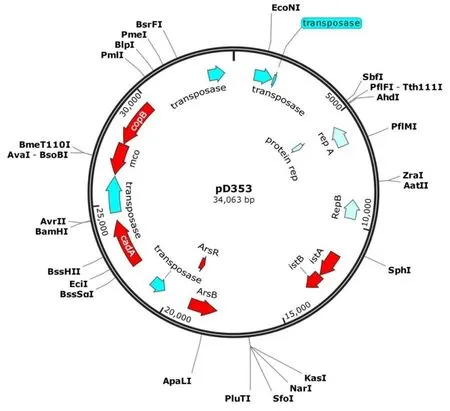
图2 pD353质粒圈
2.3 质粒进化同源
研究表明,pD353与金黄色葡萄球菌质粒pIT1-S和pIT4-R结构组成基本相同,都仅有一个模块发生逆转。与海氏肠球菌质粒pfas4-2和粪肠球菌质粒pSF2都有3个相似模块,但也有未配对序列无相似性,所以质粒pD353与此两株质粒亲缘相对较近。而与金黄色葡萄球菌质粒pSaD592相比,虽然两株质粒都来自于金黄色葡萄球菌,但两株质粒含有很多的不同模块,两株质粒亲缘较远,差异较大。pD353与pIT1-S和pIT4-R可能共同起源于海氏肠球菌和粪肠球菌质粒,由于pD353与pIT1-S和pIT4-R有更近的亲缘关系,pD353由意大利人源金黄色葡萄球菌质粒进化而来。图3

A pD353与pIT1-S,pIT4-R的亲缘关系比对
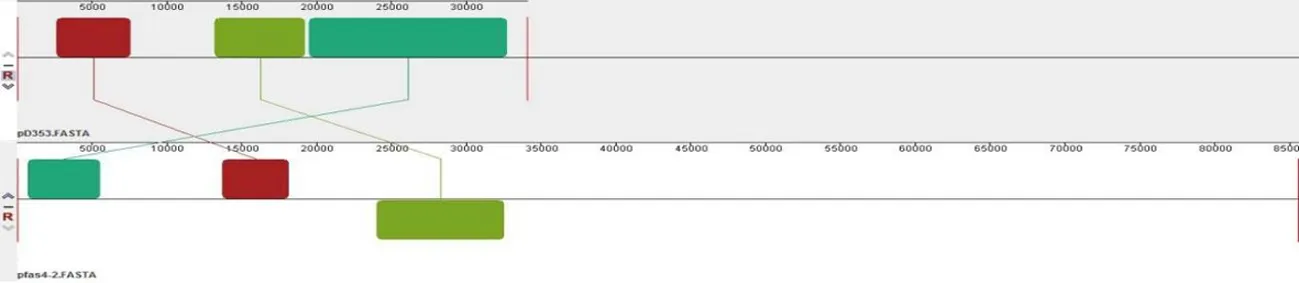
B pD353与pfas4-2的亲缘关系比对
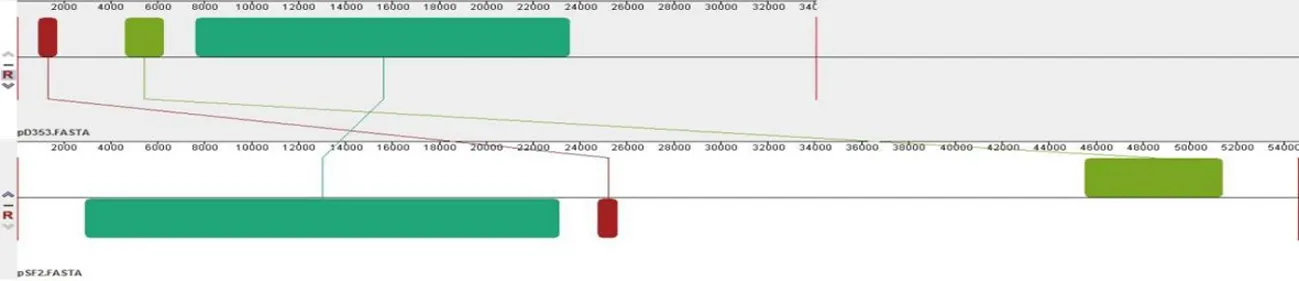
C pD353与pSF2的亲缘关系比对
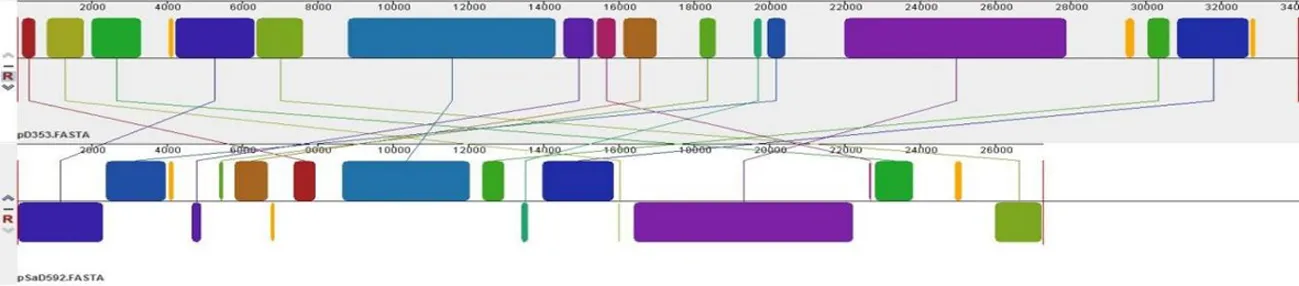
D pD353与pSaD592的亲缘关系比对
2.4 质粒复制起点
研究表明,pD353质粒的repB与pN315质粒的rep蛋白属于同种蛋白,两者都与质粒pSK639近源,预测质粒pD353和pN315同属于pSK639家族质粒,采用Theta型复制。图4
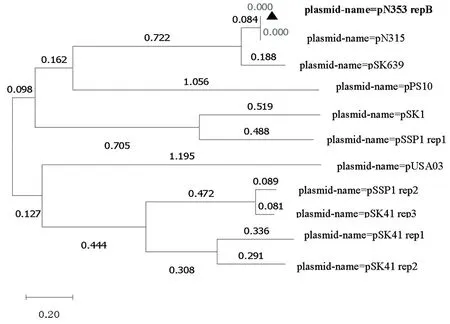
图4 质粒pD353 repB 系统发育树
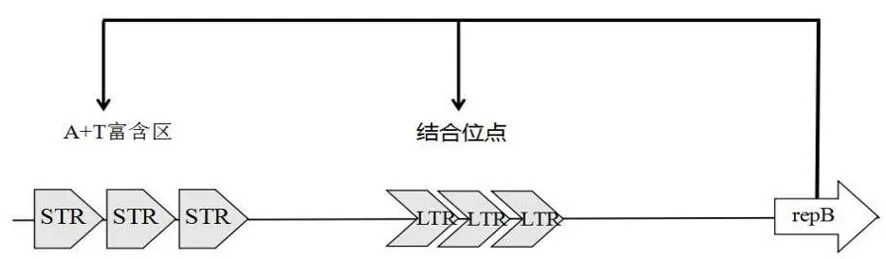
图5 质粒pD353复制起点
2.5 重金属相关基因
研究表明,pD353上含有5种的重金属转运蛋白基因。其中在22 088~24 268 bp的ORF49编码镉转运 ATP 酶cadA与已上传的金黄色葡萄球菌MRSA252菌株染色体的cadA基因相似率为99.9%。17 780~19 069 bp的ORF100编码亚砷酸盐外排转运蛋白膜亚基arsB基因,19 383~19 069 bp的ORF131编码一个As(III) 感应金属调节转录阻遏物arsR基因,这2个基因与已上传的金黄色葡萄球菌UP322染色体的arsB和arsR基因的相似率为100%。26 344~27 777 bp的ORF120编码一个多铜氧化酶mco基因,与已上传的金黄色葡萄球菌ATCC 12 600染色体的mco基因相似率为99.79%。27 792~29 921 bp的ORF150编码一个铜/银转运 P 型 ATP 酶copB基因,与已上传的金黄色葡萄球菌S3染色体的copB相似率为100%。
3 讨 论
小于10 kb的质粒使用RC机制,大于14.5 kb的质粒使用Theta模式复制,中等10至14.5 kb大小范围内的质粒使用这2个机制中的任一种[6]。近年来,金黄色葡萄球菌的耐药性日益严重[8,9],试验通过16S rRNA基因引物[10,11]确定D353为金黄色葡萄球菌之后,提取D353质粒测序,测序结果进行序列比对分析,在序列上研究发现了大量的酶切位点及丰富的重金属转运蛋白,将测得的质粒pD353归类于重金属质粒。类似的重金属质粒也被白凤岚的牦牛源MASR菌株和张赫的黄河污泥源的金黄色葡萄球菌报道[12, 13]。20世纪60年代和70年代的金黄色葡萄球菌菌株通常含有对β-内酰胺类抗生素和重金属或其他无机离子产生抗药性的质粒[14]。大量的重金属抗性基因能够帮助细菌更好的适应恶劣的环境[15]。序列共线性分析结果表明,测得的质粒pD353与海氏肠球菌和粪肠球菌质粒存在一定的序列同源性,溯源分析发现质粒pD353与意大利金黄色葡萄球菌质粒[16]的起源相同。通过对质粒复制起始蛋白repB的研究,了解到该质粒与pSK639家族质粒复制起始蛋白同源[17-23],质粒的复制起点普遍具有3个特点:支持质粒自主复制的最小的顺式作用元件;前导链合成的起始位置;打开 DNA 双链,启动复制机制的区域[24,25]。Theta型复制质粒复制起点还包括 AT 丰富区和 Rep 蛋白结合位点,后者含有相邻或是被插入序列间隔的串联重复序列[26],所以串联重复序列是复制的活性起始点所必需的[27],分析复制起始蛋白上游的串联重复序列,判断出质粒pD353为重金属Theta型复制质粒。
序列的基因分析发现在金黄色葡萄球菌质粒pD353上存在一定数量的转座酶和插入序列,其中包括IS3家族转座酶、类IS6 ISSau6家族转座酶、IS21家族转座酶、IS30家族转座酶和IS1182家族转座酶。介导DNA细胞内运动的插入序列和转座子,以及促进细胞间DNA移动的质粒、噬菌体和整合接合元件之间的相互作用[28]。这些序列元件促进了细胞内和细胞间的基因迁移,使得基因水平之间的交换成为可能[19],并且还被发现在调节基因表达和基因组重排方面发挥着额外的作用[29],但其具体的作用机制尚需进一步的实验研究。
串联重复序列对质粒拷贝数的调控具有重要意义,例如质粒R6K的复制起点γ中移除7个串联重复序列中的1个,对复制并无影响,但是当删除2个串联重复序列会降低DNA的复制效率,删除3个或以上就会停止质粒的复制,这似乎使得阻断耐药质粒的传播获得了可能,分析金黄色葡萄球菌质粒的rep蛋白复制方式,具有一定的指导意义。
4 结 论
质粒pD353与Genebank库中的意大利人源金黄色葡萄球菌质粒pIT1-S,pIT4-R有较少的模块逆转,具有非常近的亲缘关系。pD353的repB与pSK639的repB近源,trf串联重复序列发现pD353的repB序列前存在Theta型复制结构的LTR和sTR,由此鉴定质粒pD353为Theta型复制pSK639家族质粒,富含3种复制起始蛋白和丰富的重金属转运蛋白。未在该质粒发现任何的抗生素抗性基因,但该质粒特殊的Theta型复制结构对于阻断基因迁移具有较高的研究价值。
