陆地棉磷高效基因GhMYB4的克隆与表达分析
2023-07-13耿翡翡孟超敏卿桂霞周佳敏张富厚刘逢举
耿翡翡,孟超敏,卿桂霞,周佳敏,张富厚,刘逢举
(1.河南科技大学农学院/洛阳市作物遗传改良与种质创新重点实验室,河南洛阳 471000;2.中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南安阳 455000)
0 引 言
【研究意义】磷是许多分子中的关键成分,如ATP,核酸,蛋白质和磷脂等[1]。磷参与植物能量代谢,光合作用和呼吸[2]。虽然土壤中的总磷含量很高,但磷是植物生长的限制因素之一,因其在许多自然和农业生态系统中被土壤成分迅速固定[3]。土壤中的磷主要以两种形式存在,即无机磷酸盐(Pi,主要是H2PO4-和HPO42-)和有机磷酸盐,但植物获得的大部分磷以无机磷酸盐的形式吸收[4-5]。为应对土壤磷酸盐可用性低的状况,植物已经进化出许多生理生化方面的改变来优化磷从土壤中的获取以及植物体内的磷活化,使植物能够有效地适应低磷环境[6-7]。发掘及利用磷诱导的相关基因对提高作物耐低磷能力具有重要意义[8]。【前人研究进展】MYB家族成员在植物的发育过程和抗逆反应中起着核心作用。MYB蛋白参与许多生理和生化过程,包括调节代谢,控制细胞发育和细胞周期,参与响应各种生物和非生物胁迫,激素合成和信号转导[9]。在植物中,MYB转录因子家族包含4种[10],其中R2R3型MYB基因代表了MYB结构域转录因子的主要类型,具有两个重复序列(R)[11-12],且此类基因能够调节植物中的多种发育和胁迫反应[13-15]。在拟南芥中,MYB30已被证明可以调节多种胁迫反应[16],包括通过信号通路调节植物在氧化和高温胁迫下的反应[17]。在拟南芥,番茄和水稻中,其他MYB转录因子,包括AtMYB68,LeAN2以及OsMYB55,已被发现参与高温、干旱胁迫[18]。小麦MYB80的表达已被证明与转基因拟南芥的耐热性有关[19]。在水稻中,OsMYB1过表达导致对热和盐度胁迫的耐受性增加[20]。【本研究切入点】培育耐低磷高效棉花品种对提高棉花新品种选育有重要意义,有助于提高棉花效益[21]。需研究陆地棉磷高效基因GhMYB4的克隆与表达分析。【拟解决的关键问题】以陆地棉新陆早19号为材料,克隆GhMYB4基因,采用生物信息学工具研究其编码蛋白理化性质,分析其表达特性,为研究棉花GhMYB4基因的生物学功能及培育磷高效利用的棉花新品种提供基因资源。
1 材料与方法
1.1 材 料
采用前期研究筛选出的磷高效棉花品种新陆早19号为材料,在河南科技大学农学院(34°59′ N,112°42′ E)进行大田种植,选取长势一致的新陆早19号植株采集根、茎、叶、花组织,用于分析GhMYB4基因在陆地棉新陆早19号植株4个不同组织中的表达情况。
1.2 方 法
1.2.1 引物设计
在NCBI网站的EST数据库中检索差异表达基因序列得到该基因的相似序列,将所得相似性序列(覆盖率>50%,相似度>90%)使用DNASTAR的Seqman进行拼接得到重叠群,将所得序列继续检索与拼接直至没有新的相似序列出现,所得即为结果序列conting,利用ORFfinder在线平台查找开放阅读框,进行目标基因GhMYB4的克隆与分析。
根据ORFfinder在线平台查找的开放阅读框,使用Primer 5.0软件设计引物,由生工生物工程有限公司合成。表1

表1 所用引物信息
1.2.2 基因组DNA的克隆
1.2.2.1 基因组DNA的提取
取液氮速冻整株新陆早19号植株后在研钵中迅速研磨成粉末转移至离心管,利用CTAB法提取陆地棉新陆早19号的基因组DNA,向其中添加200 μL的TE缓冲液溶解后置于-20℃下保存备用。
1.2.2.2 基因克隆与测序
以新陆早19的基因组DNA为模板设计如下PCR扩增反应体系(20 μL),冰上操作:2X M5 HiPer plusTaqHiFi PCR mix (with blue dye) 10 μL,Sense primer(10 μM) 0.5 μL,Anti-sense primer(10 μM) 0.5 μL,Template DNA 0.5 μL,Nuclease-free ddH2O 8.5 μL。反应程序为95℃ 3 min;94℃ 25 s,56℃ 25 s,72℃ 40 s,32个循环;72℃ 5 min;-4℃低温保存。120 V,25 min,1%琼脂糖凝胶电泳检测。将目的DNA片段进行胶回收纯化后连接pTOPO-T载体,体系(10 μL):M5 HiPer pTOPO-TA Vector 1 μL,10×Enhancer 1 μL,纯化后的PCR产物3 μL,灭菌水5 μL。取5 μL连接产物转化50 μL大肠杆菌DH5ɑ感受态细胞,将菌液涂布在含AMP的LB固体培养基上,37℃培养过夜。随机挑取单菌落,扩大培养4 h。以所得菌液作模板进行菌液PCR反应,体系(20 μL):2× M5 HiPer plusTaqHiFi PCR mix (with blue dye) 10 μL,M13F 0.5 μL,M13R 0.5 μL,Template DNA 0.5 μL,Nuclease-free ddH2O 8.5 μL。反应程序同上,检测符合后送至生工生物工程有限公司测序。
1.2.3 cDNA的克隆
1.2.3.1 总RNA的提取及cDNA的合成
取液氮速冻整株新陆早19号植株后在研钵中迅速充分研磨成粉末,后续步骤依照所用试剂盒说明书进行,150 V,15 min,1%琼脂糖凝胶电泳检测。详细步骤参考试剂盒说明书完成cDNA的合成,-20℃保存备用。
1.2.3.2 基因克隆与测序
以新陆早19号的cDNA为模板设计如下PCR扩增反应体系(20 μL),冰上操作:2× M5 HiPer plusTaqHiFi PCR mix (with blue dye) 10 μL,Sense primer(10μM) 0.5 μL,Anti-sense primer(10 μM) 0.5 μL,cDNA 1 μL,Nuclease-free ddH2O 8 μL。PCR反应程序为:95℃ 3 min;94℃ 25 s,56℃ 25 s,72℃ 35 s,35个循环;72℃ 5 min;-4℃低温保存。120 V,25 min,1%琼脂糖凝胶电泳检测。将目的DNA片段进行胶回收纯化后连接pTOPO-T载体,体系(5 μL):M5 HiPer pTOPO-TA Vector 0.5 μL,10×Enhancer 0.5 μL,纯化后的PCR产物1.8 μL,灭菌水2.2 μL。取5 μL连接产物转化50 μL大肠杆菌DH5ɑ感受态细胞,培养后随机挑取单菌落扩大培养后进行菌液PCR反应,电泳检测符合后送至生工生物工程有限公司进行测序。
1.2.4 生物信息学
使用在线平台及软件对基因进行生物信息学分析,预测分析基因的结构、性质等。
1.2.5 半定量RT-PCR
利用半定量RT-PCR技术,分析GhMYB4基因在根、茎、叶、花4个不同组织中的表达状况。以新陆早19号各组织的cDNA为模板,GhActin作内参基因。设计PCR扩增反应体系(20 μL),冰上操作:2×M5 HiPer plusTaqHiFi PCR mix (with blue dye) 10 μL,GhMYB4-F(10 μM) 0.5 μL,GhMYB4-R(10 μM) 0.5 μL,cDNA 1 μL,Nuclease-free ddH2O 8 μL。PCR反应程序为95℃ 3 min;94℃ 25 s,60℃ 25 s,72℃ 10 s,35个循环;72℃ 5 min;-4℃低温保存。120 V,25 min,1%琼脂糖凝胶电泳检测。表1
1.2.6 荧光定量PCR
采用实时荧光定量PCR技术检测基因在根、茎、叶、花不同的4个组织中的表达情况。以陆地棉新陆早19号植株的4个不同组织的cDNA为模板,GhActin作为内参基因,进行定量分析。采用SYBR®Green ProTaqHS预混型qPCR试剂盒进行荧光定量,使用的荧光定量PCR仪器型号是CFX96,数据分析采用2-ΔΔcT法。表1
2 结果与分析
2.1 低磷胁迫差异表达序列延伸
研究表明,共检索发现并下载了16条相似序列,利用DNASTAR将所得序列全部进行拼接,得到了一个新的conting重叠群,序列长度为1 052 bp。
2.2 GhMYB4基因的克隆
研究表明,克隆得到GhMYB4基因的CDS,其条带符合目的条带大小。通过无缝克隆连接到pTOPO-T载体上,挑选阳性克隆测序。获得DNA序列1 000 bp左右与conting序列的一致性较高,为97.08%。获得cDNA序列800 bp左右与congting序列比对一致性为99.88%。开放阅读框为长度为774 bp,共编码257个氨基酸。图1

注:M:Marker 2000;1:DNA的PCR扩增片段;2:cDNA的PCR扩增片段
2.3 GhMYB4基因结构
研究表明,该基因编码序列包含5′与3′非编码序列,仅含有1个内含子,2个外显子。图2

图2. GhMYB4基因结构
2.4 生物信息学
2.4.1 基因编码蛋白的理化性质
研究表明,该基因共编码了257个氨基酸,该基因所编码蛋白的分子式是C1262H2005N375O382S13,脂肪族氨基酸指数是72.92,分子量是28 959.86D等电点为8.87,属于碱性蛋白。其中亮氨酸的含量占比最大,为10.1%,共26个。在该蛋白中带负电荷的氨基酸有31个,另外有38个带正电荷。氨基酸组分会直接影响蛋白的亲疏水性,总亲水性平均系数为-0.689,不稳定系数是42.19,属亲水性不稳定蛋白。此蛋白中精氨酸(Arg)亲水性最强,亲水指数为-4.500,异亮氨酸(Ile)疏水性最强,亲水指数为+4.500 0。
2.4.2 基因编码蛋白的保守结构域
研究表明,此基因编码蛋白匹配所属的超家族是PLN03091,该家族被归类为可能包括多个结构域的模型。该基因属于MYB基因家族,利用SMART分析GhMYB4在13-63、66-114氨基酸处存在两个SANT结构域,GhMYB4基因可能属于R2R3-MYB亚族。该基因与其他物种的MYB4氨基酸序列同源性较高,物种间MYB蛋白氨基酸长度基本一致,同源性达71.19%,2个SANT结构域所在的氨基酸几乎完全一致。图3

图3 GhMYB4与其他物种MYB家族同源蛋白的多序列比对
2.4.3 基因编码蛋白的二级结构预测
研究表明,在NPS@:SOPMA secondary structure prediction网站预测该蛋白二级结构,其中占比最大的无规则卷曲占58.37%,包含150个氨基酸,其次是α-螺旋占比为32.68%,包含84个氨基酸;此蛋白还含有少部分延伸链(Extended strand)和β-转角(Beta turn),其中β-转角占比为4.67%,有12个氨基酸,延伸链有11个氨基酸,占比为4.28%。此蛋白的二级结构由α-螺旋和无规则卷曲占据主体地位所组成,各结构在各个区间分布较为均匀,由于无规则卷曲占比较多,预测其结构比较复杂。
2.4.4 基因编码蛋白的三级结构预测
研究表明,QMEAN值为0.79,GMQE值为0.33,预测其含有转录因子WER,与R2R3-MYB转录因子识别靶基因的结构类似。蛋白的三级结构与二级结构分析结果一致,均以α螺旋和无规则卷曲为主体。图4
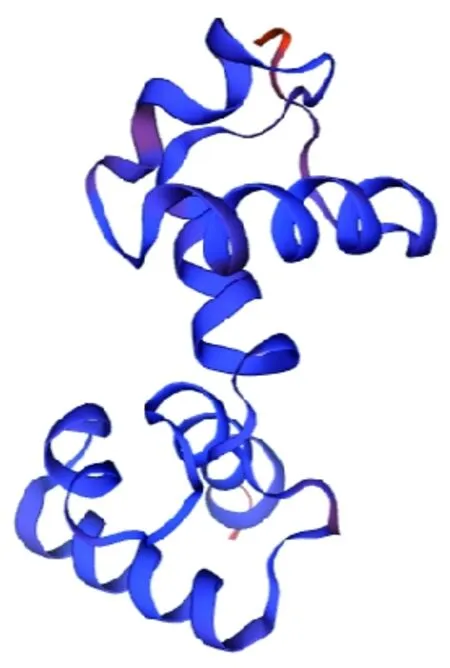
图4 GhMYB4蛋白的三级结构预测
2.4.5 基因编码蛋白跨膜结构与信号肽
研究表明,该基因所编码蛋白不含有跨膜结构域,属于非跨膜蛋白,并未形成跨膜螺旋区。此基因编码蛋白有信号肽的概率是0.001 1,其他的可能性高达0.998 9。257位氨基酸中并未出现典型的信号肽趋势,即该蛋白不存在信号肽。
2.4.6 基因编码蛋白的亚细胞定位
基因编码蛋白的亚细胞研究表明,预测定位在细胞核(Nucleus)。该蛋白是一个核蛋白,直接位于细胞核发挥作用。
2.4.7 基因蛋白互作网络预测
研究表明,获得的基因编码蛋白能够预测到蛋白质的相互作用网络,预测该基因可能是通过多个蛋白互作调控。
2.4.8 N-糖基化与磷酸化位点
研究表明,N-糖基化对蛋白稳定及功用均会产生及其重要的影响作用。获得的基因编码序列位于该蛋白质69~72、233~236位点处存在2个潜在的N-糖基化位点。磷酸化位点与蛋白的功能、结构都有很大的关系,此蛋白中占比最大的是丝氨酸和苏氨酸磷酸化位点,而酪氨酸占比较少。
2.4.9 基因编码蛋白序列系统进化树
研究表明,在NCBI数据库利用BLASTp检索氨基酸序列的同源序列,下载了另外13条不同物种的蛋白序列,木槿(Hibiscussyriacus)、可可树(Theobromacacao)、榴莲(Duriozibethinues)、桃树(Prunuspersica)、苹果树(Malusdomestica)、月季(Rosachinensis)、拟南芥(Arabidopsisthaliana)、冬瓜(Benincasahispida)、黄瓜(Cucumissativus)、枇杷(Rhaphiolepisbibas)、梅花(Prunusmume)、芝麻(Sesamumindicum)、核桃(Juglansregia)。棉花蛋白序列与木槿、可可树、榴莲的亲缘关系最近,与其余物种的亲缘关系较远,不在一个大支上。图5

图5 GhMYB4基因系统进化树
2.5 半定量RT-PCR
研究表明,GhMYB4基因高表达于根,微量表达于花,中量表达于茎和叶。图6

注:M:Marker 2000
2.6 基因相对表达量
研究表明,棉花磷高效基因在根、茎、叶和花4个组织中都有一定量的表达;但GhMYB4基因在新陆早19号植株的不同组织中的表达情况各不相同,其中在新陆早19号的根系中的相对表达量最高,其次是叶,而在花组织的表达相对较少,茎中的表达量略高于花,根中的表达量约为花中表达量的2倍。图7

图7 GhMYB4在棉花不同组织中的相对表达
3 讨 论
磷影响植物的生物调节(如能量代谢、信号转导和酶调节)和生理过程(如花青素积累和向根际释放有机酸)。为了在缺磷条件下维持细胞内磷稳态,植物主要是通过从土壤中获取磷、磷从根到芽的转移以及植物体内部磷的再活化来实现的[22]。尽管许多研究报告了磷相关的转录因子,但在磷胁迫中直接调节磷酸转运蛋白的几种转录因子的功能与分子调控机制仍未在棉花中得到表征[23]。MYB蛋白包括一大家族的植物转录因子,其成员在植物生物过程中执行多种功能。尚未在棉花中对该基因家族进行全基因组鉴定。第一个植物MYB基因是从玉米中分离出来的,其编码参与花青素生物合成的c-MYB样转录因子,随后在许多植物中鉴定出越来越多的植物R2R3-MYB基因成员[24]。研究表明,R2R3-MYB家族基因除了参与各种生理活动外,还在植物对非生物胁迫的反应中起重要作用[25]。
4 结 论
所得基因序列与拟南芥中的MYB家族中的MYB4基因同源性较高,从陆地棉新陆早19号中克隆得到了一个MYB4类似基因,并将其命名为GhMYB4,该基因属于MYB超家族成员。该基因的结构相对较为简单,仅有一个内含子,GhMYB4蛋白相对分子质量(Molecular weight)是28 959.86D,理论等电点(Theoretical pI)为8.87,是碱性蛋白,具备不稳定性与亲水性。此蛋白的二级结构由α-螺旋和无规则卷曲占据主体地位所组成,通过同源建模可知该基因三级结构预测其含有转录因子WER。该基因编码蛋白不存在信号肽和跨膜结构域。该蛋白存在2个潜在的N-糖基化位点和多个磷酸化位点。亚细胞定位结果显示该基因编码蛋白在细胞核中,预测其能够通过蛋白质的相互作用参与调控。该基因在棉花根中的表达量是最大的,在花中表达相对较小,中量表达于茎和叶,GhMYB4基因在根系发挥作用,在棉花的磷高效利用信号调控过程中具有重要作用。
