海南中部典型槟榔-象草复合系统中根系空间分布及地下竞争
2023-07-11周华熊嘉乾吴金群郑晓敏黄海陈海辉余雪标
周华 熊嘉乾 吴金群 郑晓敏 黄海 陈海辉 余雪标

关键词:农林复合系统;根系;空间分布;种间竞争
中图分类号:S344.2 文献标识码:A
农林复合经营是一种高效的农业生产方式,可使单位面积耕地产量和经济效益最大化,在实际生产中占据重要的地位。国内外众多学者对农林复合系统地下环境的竞争研究表明,部分林木和作物之间的地下竞争要比地上更为激烈[1-3]。根系作为植物的重要器官,承担着植物吸收水分和矿质元素的任务,直接关系到植物对水分及养分的吸收利用,而农林复合系统中林木和作物根系的空间分布决定了其对地下资源的竞争能力[4]。有研究表明,在农林复合系统中,通过合理的树种选择及空间配置,可以使林木和作物根系在空间利用上达到互补,避开强烈的种间竞争,从而提高物种对于资源的利用率,提高复合系统的生产力[5]。所以研究复合系统中林木与作物根系在土壤中的空间分布及其生长发育状况,对研究系统内各物种的关系具有十分重要的意义[6-7]。
槟榔(Areca catechu L.)属多年生常绿乔木植物,原产于马来西亚,是我国热带地区重要的经济作物[8],主产区为海南省与台湾省,在广西和广东等地也有少量的栽培,其种植规模逐步由单一零散栽培发展为大规模生产[9-10]。槟榔种植株行距一般为2.5 m×2.5 m,林下空间充足,合理的间作设置可以充分利用槟榔林内的资源,改善槟榔的生长环境[11]。有研究表明,槟榔林下栽培香草兰能够有效改善土壤养分状况和微生物环境条件,促进槟榔和香草兰的生长,提高槟榔林的整体经济效益[12-13]。槟榔复合栽培香露兜后,可以提高土壤酶活性,进而促进槟榔根系生长[14]。在槟榔复合栽培胡椒的模式中,胡椒根系总根长相比于胡椒单一栽培时有所减小,但是根系表面积增大,相比之下,槟榔-胡椒复合栽培模式具有明显的产量优势[15]。
象草(Pennisetum purpureum Schum.),又称紫狼尾草,原产于非洲,是适口性好、适应能力强的多年生丛生大型草本植物[16]。由于其产量高、适应性广、管理粗放、易于栽培、病虫害少,得以在热带亚热带地区广泛推广应用,是我国南方重要牧草品种[17-18]。
目前,关于槟榔–象草复合系统中,二者的根系分布和地下竞争状况尚不明确。本研究拟通过对槟榔-象草复合系统中的根系进行调查研究,比较二者根系的空间分布特征,探讨二者的种间地下竞争关系,从而为进一步发展和扩大槟榔-象草高效复合栽培模式提供理论依据。
1 材料与方法
1.1 试验地概况
研究区位于海南省屯昌县坡心镇洪涛坡农场(19°17′10″N,110°10′52″E,海拔105 m),地处海南省中部偏北,五指山北麓,南渡江南岸,属于典型热带海洋季风区。春季常有干旱,夏季高温多雨,夏秋多台风,冬季凉爽多雨,年平均降水量1960~2400 mm,以5—11 月为主。年平均气温23.5 ℃,最高年平均气温28.3 ℃,最低年平均气温20.3 ℃。
所选的槟榔-象草复合经营地块,槟榔林龄12 年,株行距2.0 m×3.0 m,胸径(8.67±0.68)cm,树高(4.64±0.81)m,東西冠幅(1.83±0.35)m,南北冠幅(1.78±0.38)m。象草为2016 年扦插种植,种植时距离槟榔树行30 cm,株行距0.4 m×0.5 m,调查时高度(1.54±0.42)m。
1.2 方法
1.2.1 槟榔与象草根系的辨别 槟榔、象草根系的形态和颜色差别较大,根据二者根系的形态和颜色,能够很好地将象草和槟榔根系区分开来。
颜色上,槟榔根系呈褐色且颜色偏暗,象草根系呈淡棕色;形态上,槟榔根系较粗且比较零散,而象草根系须根多且比较聚集。
1.2.2 根系分布特征测定 采用带状分层挖掘法,于2021 年7 月下旬对槟榔-象草复合模式的根系进行取样研究。在选定的样地中,对槟榔进行每木检尺,选择4 株树高、冠幅和胸径相近且生长状况良好的槟榔树作为标准木,在垂直于槟榔树行方向,以槟榔树干基部外20 cm 为起点,挖一条长140 cm、宽20 cm、深40 cm 的样带,水平方向上每20 cm 为一个取样点,即距树干基部20~40、40~60、60~80、80~100、100~120、120~140、140~160 cm,分别记为S1、S2、S3、S4、S5、S6、S7,垂直方向上每20 cm 为一层进行取样。最终每个样方取0.2 m×0.2 m×0.2 m 共计0.008 m3的土壤,将含有槟榔和象草根系的整个土壤样品进行称重,随后将根系从中挑出;从挑出根系后的剩余土壤中取1 kg 左右的样品称重后装入密封袋中并编号,带回实验室冲洗并收集剩余根系,用于降低实验误差。返回实验室后,将根系样品用清水浸泡1 h 以上,搅拌均匀,倾倒在孔径为0.2 mm 的土筛中,将土筛悬浮在水盆中不断摇动冲洗,洗去土壤。冲洗后分别拣出槟榔和象草根系。
用游标卡尺测量槟榔根系直径,将其分为细根(0~2 mm)、中根(2~5 mm)和粗根(>5 mm),象草根系较细,不再进行分级。从已分类的槟榔根系中挑选出具有代表性的根系,测量其长度(精确至0.1 cm)和重量(精确到0.001 g),用于计算根系总长度。然后将根系放入烘箱中,烘干至恒重,测量其生物量(精确至0.01 g)。
1.2.3 地下种间竞争强度的度量 Levins 生态位重叠表示2 种物种利用同一资源而相互重叠的情况,可以从某种意义上反映出由于生态位重叠所造成的竞争[19]。利用Levins 生态位重叠公式计测槟榔和象草间的地下竞争指数,竞争指数越大代表竞争能力越强[20],数学表达式为:
1.3 数据处理
用Microsoft Excel 2019 和SPSS 22.0 软件对数据进行统计整理和单因素方差分析(one-wayANOVA),检验种内根系生物量密度、根长密度水平分布差异的显著性,并采用Tukey 进行多重比较。对槟榔和象草的竞争强度指数、根系生物量密度、根长密度垂直分布的差异显著性采用Paired-samples T test 检验法进行检验; 用GraphPad Prism 8.0.2 软件制图。
2 结果与分析
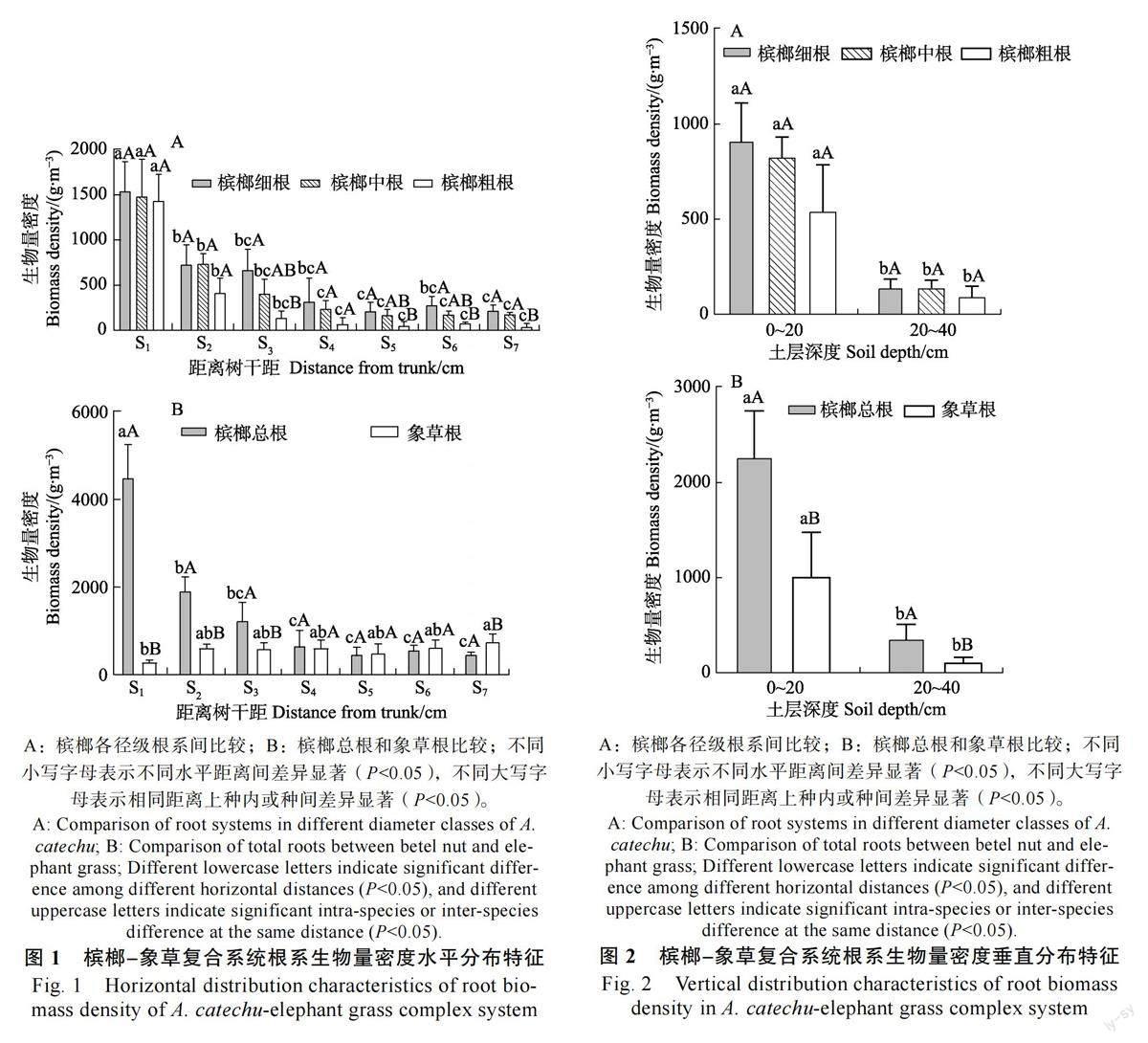
2.1 槟榔-象草复合系统根系生物量密度分布特征
取得的56 个样品中,土壤总体积为0.448 m3,其中槟榔细根、中根、粗根以及总根的生物量分别为244.56、216.02、141.48、602.06 g;象草根系的生物量为250.22 g。槟榔和象草根系的生物量密度分别为1343.88、558.53 g/m3,槟榔根系生物量是象草的2.41 倍。
2.1.1 生物量密度水平分布特征 水平方向上,檳榔细根、中根、粗根以及总根生物量密度整体上随着距树干基部距离的增加而下降,象草根系生物量密度分布则较为均匀(图1)。由S1 至S7槟榔细根生物量密度从(1541.15±327.71)g/m3 下降到(216.23±64.43)g/m3, 中根生物量密度从(1482.25±415.24)g/m3下降到(175.31±23.56)g/m3,粗根生物量密度从(1431.93±297.84)g/m3下降到(36.79±40.89)g/m3,总根生物量密度从(4455.32±795.73)g/m3下降到(428.33±87.81)g/m3,差异均极显著(P<0.01);象草根系生物量密度整体呈增加趋势, 从(281.49±57.70)g/m3增加到(742.45±186.35)g/m3,差异显著(P<0.05)。
水平距离上,槟榔根系生物量的组成均表现为细根>中根>粗根,说明细根是决定槟榔根系分布的主要因素。随着距槟榔树干水平距离的增加,各距离上粗根生物量密度所占的比例逐步减小。各水平距离上槟榔和象草相比,槟榔根系的生物量密度在S1、S2、S3及S4 上大于象草,且在S1、S2和S3上差异显著(P<0.05);在S5、S6及S7上槟榔根系的生物量密度均小于象草,且在S7 上差异显著(P<0.05)。综上,在水平方向上,不同区域内槟榔和象草根系生物量密度的分布各有优势:在距槟榔树干80 cm 内时,槟榔根系生物量密度显著大于象草根系;120~140 cm 处象草根系生物量密度显著大于槟榔;其余水平距离内二者之间的差异不显著。
2.1.2 生物量密度垂直分布特征 垂直方向上(图2),槟榔细根、中根、粗根、总根以及象草根系生物量密度在0~20 cm 土层中均显著大于20~40 cm 土层。由0~20 cm 土层到20~40cm 土层:槟榔细根生物量密度由(907.25±205.01)g/m3降到(134.16±51.22)g/m3, 中根生物量密度由(826.21±107.51)g/m3降到(138.18±42.61)g/m3,差异极显著( P<0.01 ); 粗根生物量密度由(540.83±246.41)g/m3降到(90.79±57.20)g/m3,差异显著( P<0.05 ); 槟榔总根生物量密度由(2274.28±480.44)g/m3降到(363.13±144.89)g/m3,差异极显著(P<0.01);象草根系生物量密度由(1009.76±467.67)g/m3降到(107.30±57.38)g/m3,差异显著(P<0.05)。
槟榔各径级根系间比较,各土层根系生物量的组成均表现为细根>中根>粗根,但三者之间差异不显著(P>0.05)。槟榔和象草相比,各土层槟榔根系的生物量密度均大于象草,在0~20 cm 土层差异极显著(P<0.01),在20~40 cm 土层差异显著(P<0.05)。综上,在垂直方向上,2 个土层中槟榔根系生物量密度均显著大于象草,且二者根系均主要集中于0~20 cm 土层;2 个土层中的槟榔根系均以细根和中根为主,粗根所占比例最小。
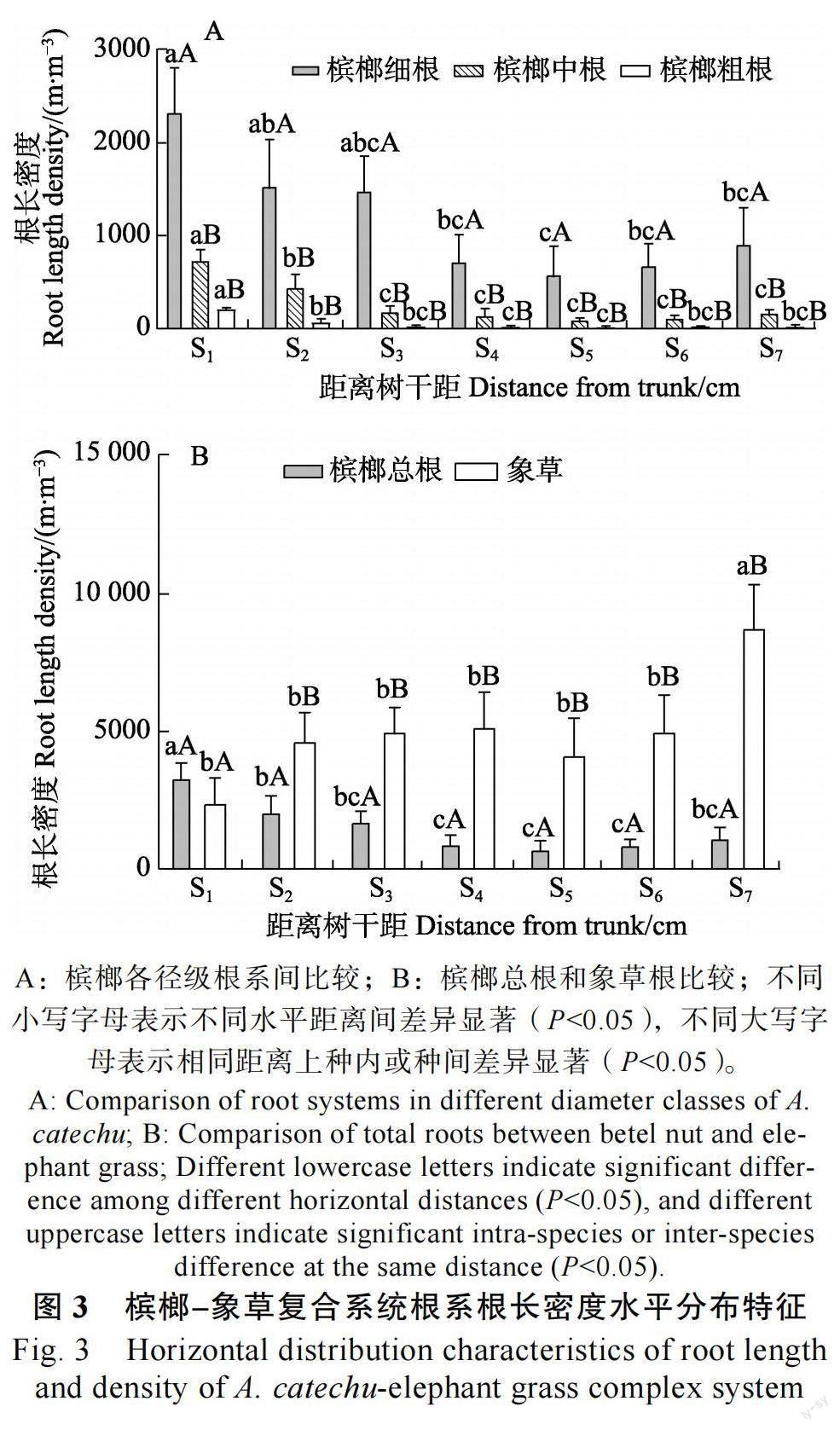
2.2 槟榔-象草复合系统根系根长密度分布特征
2.2.1 根长密度水平分布特征 水平方向上(图3),随着距槟榔树干基部距离的增加,槟榔细根、中根、粗根、总根的根长密度整体呈下降趋势,象草根长密度则呈增加趋势。从S1到S7,槟榔细根根长密度从(2036.11±506.28)m/m3 下降到(882.78±420.64)m/m3,中根根长密度从(725.63±126.09)m/m3下降到(158.58±44.79)m/m3,差异均极显著(P<0.01);槟榔粗根根长密度从(206.01±17.06)m/m3下降到(17.36±27.37)m/m3,差异显著( P<0.05 ); 槟榔总根根长密度从(3237.75±621.02)m/m3下降到(1058.72± 462.25)m/m3,差异极显著(P<0.01);象草根系根长密度从(2369.25±945.49)m/m3增加到(8725.75±1615.88)g/m3,差异极显著(P<0.01)。
各水平距离上槟榔各径级根系间比较,根长密度的组成均表现为细根>中根>粗根,且细根显著大于中根和粗根,中根和粗根之间差异不显著,说明细根是槟榔根长的主要组成部分。各水平距离上槟榔和象草相比,槟榔根长密度只在S1 处大于象草(P>0.05),在S2至S7上均显著小于象草(P<0.05)。综上,在水平方向上,象草根长密度的分布比槟榔更有优势,在距槟榔树干20 cm 内槟榔总根长密度大于象草,20 cm 外象草根长密度显著大于槟榔。
2.2.2 根长密度垂直分布特征 垂直方向上(图4),槟榔细根、中根、粗根、总根以及象草根长密度在0~20 cm 土层中均显著大于20~40 cm 土层。由0~20 cm 土层到20~40 cm 土层,槟榔细根根长密度由(2204.8±690.07)m/m3下降到(241.69±165.00)m/m3, 中根根长密度由(449.52±91.03)m/m3下降到(66.78±15.18)m/m3,差异均极显著(P<0.01);槟榔粗根根长密度由(79.64±33.77)m/m3下降到(16.33±7.00)m/m3,差异显著(P<0.05 ); 槟榔总根长密度由(2733.96±780.02)m/m3降到(324.81±180.54)m/m3,差异极显著( P<0.01 ); 象草根长密度由(9170.40±1476.02)m/m3降到(970.57±211.27)m/m3,差异极显著(P<0.01)。
垂直方向上槟榔各径级根系间比较,各土层根长密度的组成均表现为细根>中根>粗根。各土层上槟榔和象草相比,槟榔根长密度均小于象草,且在2 个土层上均有显著差异(P<0.05),二者根系均主要集中于0~20 cm 土层。
2.3 槟榔-象草复合系统地下竞争指数
水平方向上(图5A),在S1和S2处槟榔根系的竞争指数(5.09±1.28;1.25±0.27)均大于象草根系(0.19±0.08;0.81±0.14),且在S1 處差异极显著(P<0.01)。其他距离处,象草根系的竞争指数均大于槟榔根系,且在S4至S7处差异显著(P<0.05)。因此,随着距槟榔树干距离增加,槟榔根系的竞争能力逐渐降低,象草根系的竞争能力逐渐增强。二者相比,距离槟榔树干60 cm 以内槟榔根系的竞争能力占据优势,60 cm 以外则是象草根系的竞争能力更强。垂直方向上(图5B),0~20 cm 土层和20~40 cm 土层槟榔的竞争指数(0.74±0.12;0.82±0.28)均大于象草(0.45±0.11;0.26±0.07),且差异显著(P<0.05)。综上,在2个土层中,槟榔根系的竞争能力均显著大于象草根系。
3 讨论
3.1 槟榔与象草根系空间分布特征
本研究发现,在槟榔-象草复合系统土层深度0~20 cm、距树干基部20~160 cm 的地下空间中,水平方向上槟榔各径级根系的生物量密度随着距树干距离的增加而下降,象草根系的生物量密度和根长密度分布则比较均匀,主要集中在距槟榔树干40~160 cm 内,且整体呈增加趋势。在距槟榔树干20~40 cm 处是槟榔根系最集中的区域,同时也是象草根系分布最少的区域,该区域内槟榔根系对于象草的生长影响较大,象草根系生物量密度的增加能够提高其对土壤水分和养分的竞争吸收优势,同时也是对槟榔根系竞争力减弱的积极回应[21-22]。云雷[22]、李洁[23]通过对晋西黄土区苹果农作物间作系统的研究发现,垂直方向上苹果细根在20~40 cm 土层深度分布最多。而本研究发现,同样作为经济林树种间作模式,槟榔和象草的根系在垂直方向上具有明显的分层现象,象草根系90.4%分布于0~20 cm 土层,仅有9.6%分布于20~40 cm 土层,0~20 cm 土层中的生物量密度是20~40 cm 土层的9.4倍;槟榔各径级根系在0~20 cm 分布量同样显著高于20~40 cm 土层。由此可知,垂直方向上槟榔和象草的根系分布并未出现错开情况用于避免竞争,这说明二者根系在2 个土层中能够实现较好的共存,正常生长。
3.2 槟榔-象草复合系统地下竞争
在植物生活史、群落结构和动态以及植物形态变化等方面,竞争是主要的动力之一[24]。丁怡飞等[25]利用Levins 生态位重叠公式计算油茶和鼠茅草的地下竞争指数,其实质是分析油茶和鼠茅草对地下空间资源的共同占有率,表示物种间的生态位重叠。在地下生态系统中,植物吸收土壤资源的能力以及物种间的竞争强度由根系的密度和空间分布决定。因此,槟榔–象草复合系统中二者根系的空间分布格局和数量的多寡就决定了槟榔和象草的种间竞争能力。
本研究表明,在距槟榔树干20~60 cm 水平范围和0~40 cm 的土层内,槟榔根系的竞争指数高于象草根系;在距离槟榔树干60~160 cm 水平范围内,象草根系的竞争指数显著高于槟榔根系,平均高0.73,此结果与骆宗诗等[26]研究花椒林中椒草种间地下竞争相似。这可能是因为在槟榔-象草复合系统中,槟榔根系大部分集中在距树干20~60 cm 范围内,此区域内绝大部分空间被槟榔根系占据,象草根系可利用空间较少,因而槟榔竞争能力更强;随着水平距离的增加,槟榔根系减少,象草根系得以更好地扩展,竞争力增强。因此槟榔与象草复合种植时,象草应该距离槟榔树60 cm 以外开始种植最为合适。
丁怡飞等[25]对油茶–鼠茅草间作的种间地下竞争研究表明,在二者间作系统中,油茶平均竞争指数大于鼠茅草,其中距油茶树干60 cm 内油茶的竞争能力要大于鼠茅草,60~90 cm 内鼠茅草竞争能力更强,此结果与本研究一致。但在垂直方向上,由于鼠茅草87.5%的根系分布于0~20 cm土层,为了避开竞争,油茶根系出现了下移现象,更多地集中在20~40 cm 土层,这与本研究有所不同。这可能是因为油茶为深根性树种,花椒为浅根性植物,而槟榔与花椒相同,也属于浅根性植物[27-29]。在空间配置上,槟榔-象草复合模式中二者根系垂直方向未出现避让现象,会产生更直接的竞争,另一方面也可以说明二者根系分布均未对对方造成绝对的影响,都能够保证自身正常生长。
槟榔和象草的根系分布在空间上虽然有重叠,但二者在不同区域内各有优势,均不能对另一方形成绝对竞争优势,从而二者都能够在竞争中健康生长。因此,为了有效降低槟榔和象草之间的竞争,同时充分发挥各自优势,将二者产量效益最大化,应适当增加象草与槟榔树间的种植距离,并适当加强间作区域内0~20 cm 土壤的水肥投入。
