CYP74A2在白粉菌诱导月季对甜菜夜蛾产生抗性中的功能及其生物信息学分析
2023-07-10杨琦刘思琪李云仙杨发忠
杨琦 刘思琪 李云仙 杨发忠
摘要:为明确CYP74A2基因在白粉菌诱导月季产生对甜菜夜蛾的抗性过程中的功能,并对CYP74A2作生物信息学分析,利用转录组测序获得的差异表达基因进行功能富集分析。结果表明,α-亚麻酸代谢产生的茉莉酸是白粉菌诱导月季产生对甜菜夜蛾抗性的主要挥发性成分之一,发现该通路中共有5个CYP74A2差异表达基因,分别是LOC112173678、LOC112172343、LOC112172188、LOC112173396和LOC112169957。采用qRT-PCR验证CYP74A2基因的功能,利用NCBI、KEGG等数据库找到CYP74A2同源蛋白并结合ExPASy-ProtParam、TMHMM、SignalP 6.1、MEME、SOPMA、SWISS-MODEL、MEGA 11等在线工具或软件对CYP74A2进行生物信息学分析。通过ExPASy-Protparam分析发现,CYP74A2蛋白分子量都在54.3 ku左右,是一种稳定的亲水性蛋白。该类蛋白无跨膜结构和信号肽,亚细胞定位于细胞质和叶绿体中,少量分布于质膜和细胞核内。SOPMA预测结果表明该类蛋白以α-螺旋为主,SWISS-MODEL对CYP74A2蛋白三维建模的结果与SOPMA预测结果一致。系统发育分析结果表明,月季CYP74A2蛋白与草莓CYP74A2蛋白亲缘性最近。MEME对蛋白的保守基序分析结果表明,CYP74A2蛋白具有底物识别位点SRS-1、P450s的保守序列、血红素结合域以及血红素结合域的Ⅰ型螺旋。研究结果为探明CYP74A2基因的生物学功能及白粉菌诱导月季产生对甜菜夜蛾抗性的生物分子学机制提供了依据。
关键词:月季;白粉菌;甜菜夜蛾;CYP74A2;茉莉酸;生物信息学
中图分类号:S436.8+1文献标志码:A文章编号:1002-1302(2023)11-0119-09
月季(Rosa chinensis Jacq.)别称玫瑰、月季花、中国月季,在云南省的鲜切花生产中,切花月季占据了鲜切花产量的40%以上[1-2]。甜菜夜蛾[Spodoptera exigua (Hübner)]和月季白粉菌[Podosphaera pannosa (Wallr. ∶Fr.)]是云南省月季种植过程中主要的有害生物[3-4]。甜菜夜蛾是鳞翅目夜蛾科的一种杂食性害虫,其寄主植物达170多种。目前,还没有对甜菜夜蛾有效的生物防治,而长期使用化学试剂会导致土壤污染以及甜菜夜蛾耐药性的增强,据统计,甜菜夜蛾已对38种有效化学成分产生抗药性[5]。
Karban等研究发现,真菌病原体大丽轮枝菌(Verticillium dahliae)感染棉花幼苗后,二斑叶螨(Tetranychus urticae)的种群增长速度降低[6]。这一结果表明,2种高度无关的生物如果共享同一寄主,它们之间则可能发生强烈的相互作用,这揭开了由寄主植物介导的病虫互作关系的序章。该领域也成为国内外研究的热点[7-9],但是截至目前,该领域的研究主要是从现象上阐明了共享寄主的病虫间存在着寄主植物介导的间接互作关系,而对于互作的生物分子学机制研究仍不清楚,特别是关于白粉菌诱导月季对甜菜夜蛾产生抗性的分子生物学的研究仍然是未知的。研究表明,白粉菌侵染可诱导月季产生对甜菜夜蛾的抗性[4,10-12],虽然白粉菌诱导月季对甜菜夜蛾产生抗性的化学机制已被探明,但其生物分子学机制仍是未知的。
CYP450s是一种单加氧酶,能催化植物的多种初级和次级代谢反应,其催化作用的共同点是在底物分子中加入一个氧原子。CYP450s是一个庞大的家族,包括1 000多个家族和2 500个亚家族,在植物次生代謝方面有着非常广泛和复杂的功能,该超家族的特点是结构中均存在“FxxGxRxCxG”的保守血红素结合域。CYP450s超家族在植物的脂肪酸代谢、次生代谢产物的合成、植物激素的生物合成与降解等代谢通路中以及植物抵抗病虫害方面发挥了重要作用[13]。而P450的CYP74家族的酶无需氧就可催化脂肪酸过氧化物形成挥发性物质或丙二烯氧化物,该类家族包括2种脱氢酶[丙二烯氧化合酶(AOS)和二乙烯基醚合成酶(DES)]以及2种异构酶[氢过氧化物裂解酶(HPL)和环氧醇合酶(EAS)][14]。丙二烯氧化合酶是参与α-亚麻酸代谢的主要脱氢酶[15]。亚麻酸属于脂肪酸,其分解产物之一的茉莉酸(JA)不仅是重要的植物生长调节物[16],还是白粉菌诱导月季对甜菜夜蛾产生抗性的挥发性成分[17]。
因此,本研究利用月季转录组中的CYP74A2差异表达基因,通过生物信息学分析方法分析CYP74A2基因,以确定月季抗甜菜夜蛾关键候选基因。可为病虫互作机制的系统深入研究提供科学依据,为月季鲜切花生产过程中的病虫害防治提供新思路,为绿色环保、可持续发展的现代植保理念助力,为高原特色现代化农业的健康发展提供动力。
1材料与方法
1.1试验材料
试验材料为中国月季品种艳粉,采自云南省昆明市呈贡区斗南花卉种植基地(102.78°E,24.90°N)。
1.2转录组测序
将感染白粉菌前后的月季叶片用去离子水冲洗揉搓后,分开用液氮冷冻保存。将样本送至上海美吉生物医药科技有限公司测序,样本用Trizol试剂法提取总RNA并去除DNA污染。用NanoDrop 1000检测RNA纯度及浓度后,对2组样本(每个样本3次重复)共6份待测样本分别富集mRNA,然后将mRNA进行随机打断。以mRNA作为模板,合成第1条cDNA链,随后合成第2条cDNA链。将双链的cDNA补成平末端后在3′末端加上“A”碱基,最后采用Illumina Novaseq 6000对修饰后的cDNA进行高通量测序。
测序完成获得该物种的Unigene库后,利用R语言编写脚本将Unigene序列与Swiss-Prot(http://web.expasy.org/docs/swiss-prot_guideline.html)、GO(http://www.geneontology.org)、KEGG(http://www.genome.jp/kegg/)、NCBI(ftp://ftp.ncbi.nlm.nih.gov/blast/db/)、Pfam(http://pfam.xfam.org/)等数据库对比以获取注释信息。
参考基因组:Rosa chinensis;参考基因组版本:GCF_002994745.1;参考基因组来源网站:https://www.ncbi.nlm.nih.gov/genome/11715?genome_assembly_id=366730。
1.3差异表达基因的筛选
获取注释信息后对基因进行差异表达分析,从而鉴定出样本间的差异表达基因。使用DESeq2软件(版本1.38.0,http://bioconductor.org/packages/stats/bioc/DESeq2/)对源数据进行标准化处理,随后筛选出组间的P450差异表达基因,筛选条件为 P-adjust<0.05且|log2FC| ≥ 1,采用BH(Benjamini & Hochberg)法校正,上下调差异倍数2.0倍。
1.4P450基因的生物信息学分析
利用ExPASy-ProtParam(https://web.expasy.org/protparam/)对蛋白的等电点、不稳定系数和分子量等理化性质进行分析;利用ExPASy-ProtScale(https://web.expasy.org/protscale/)预测蛋白的亲疏水性;利用TMHMM(https://dtu.biolib.com/DeepTMHMM)预测蛋白质的跨膜区域;利用在线工具SignalP 6.1(https://services.healthtech.dtu.dk/service.php?SignalP)对信号肽进行预测;利用WolfPsort(https://wolfpsort.hgc.jp)对P450蛋白进行亚细胞定位;利用在线预测工具MEME(https://meme-suite.org/meme/index.html)对蛋白结构进行分析;利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测蛋白二级结构;利用在线工具SWISS-MODEL(https://swissmodel.expasy.org/)预测蛋白质三维结构。通过NCBI序列比对,获取CYP74A2的同源蛋白后,利用MEGA 11对该类蛋白构建系统进化树(NJ法,Bootstrap值为1 000)。
1.5差异表达基因的qRT-PCR验证
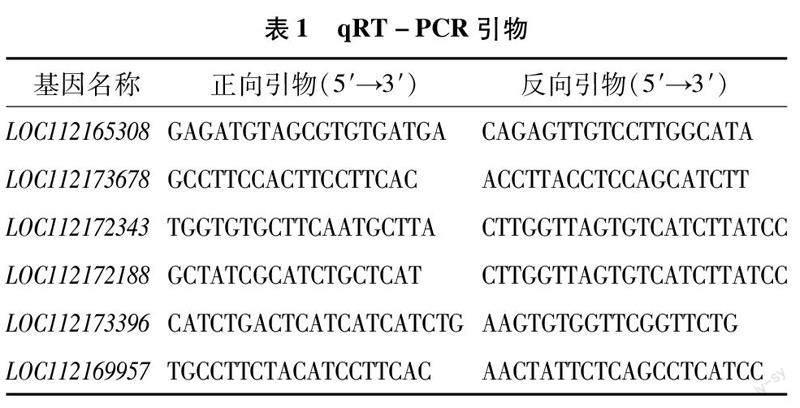
利用提取的RNA进行逆转录合成cDNA,每个样本设置3次生物学重复。以月季染白粉菌前后稳定表达的基因LOC112165308为内参基因,选择白粉菌诱导月季产生茉莉酸的5个CYP74A2差异表达基因,利用Primer Premier 6.0软件设计qRT-PCR引物(表1), 将月季叶片样本与引物信息一并送至北京擎科生物科技有限公司昆明分公司进行qRT-PCR荧光定量验证。qRT-PCR的扩增流程为95 ℃预变性120 s;95 ℃变性10 s,60 ℃退火 30 s,72 ℃延伸120 s,进行30个循环。
2结果与分析
2.1转录组测序及序列比对结果
以未感病月季叶片样本为对照组(CK组),感病月季叶片样本为实验组(T组)转录组测序共统计到表达基因37 309个,其中已知基因32 950个,新基因4 359个。各样品质控后的数据总量(clean data)均达到6.88 Gb以上,Q20、Q30碱基比例分别在98.20%、94.39%以上,测序碱基平均错误率在0.025%以下(表2),结果表明测序质量较好。
与参考基因组相比,定位到基因组上的clean reads占总clean reads数的86.50%左右。其中,有唯一比对位置的clean reads数量占比最大,在83.60%~84.45%之间;有多个比对位置的clean reads占比最小,在2.50%左右。reads区域分布方面,比对到蛋白质编码区(CDS)的clean reads占比最大,最高达到87.29%;比对到基因间区的最小,在0.70%~0.78%之间;比对到内含子区域的也较低,为1.68%~2.04%;而非编码区的则在 10.29%~11.34%之间(表3)。总体来看,有超过83%的clean reads能定位到基因组中且有唯一比对位置,低于3%的clean reads有多个比对位置。Reads区域分布中,超过85%的reads比对到编码区,比对到非编码区的低于12%。
2.2差异表达基因功能富集分析
转录组测序共统计到1 646个差异表达基因,其中上调基因1 251个,下调基因395个。KEGG富集分析结果表明, 富集到的KEGG通路共81条,有56条通路参与新陈代谢,主要包括酪氨酸、丙氨酸、天冬氨酸和谷氨酸代谢,萜类化合物和聚酮化合物的代谢、次生代谢产物的生物合成、脂质代谢、碳水化合物代谢等;有12条通路参与了遗传信息处理,主要包括内质网中的蛋白质加工、DNA复制、不匹配修复、同源重组及泛素介导的蛋白水解;有12條通路参与环境信息处理,主要包括磷脂酰肌醇信号系统、MAPK信号通路、植物激素信号转导;有4条通路参与细胞过程,主要包括细胞衰老、内吞、坏死;有5条通路参与生物体系统,主要包括植物-病原体相互作用、胆固醇代谢、昼夜节律等。
差异表达基因的GO功能富集分析结果表明,共250条GO term被显著富集。其中,参与分子功能(MF)的GO term共156条,主要包括酶活性、酶抑制剂活性、脂质结合、有机酸结合、单氧酶活性、氧化还原酶活性等;参与生物过程(BP)的共85条,包括信号传导途径、防御反应、糖类的分解代谢过程、黄酮类的生物合成及代谢过程、脂质分解代谢过程等;参与细胞组成(CC)的共9条,主要包括细胞外区域、叶绿体、膜的整体成分等。图1为富集最显著的前20条KEGG通路、GO term。
2.3α-亚麻酸代谢中的CYP74A2差异表达基因
富集程度最显著的KEGG代谢通路中的α-亚麻酸代谢(图2)产生的茉莉酸是甜菜夜蛾的天然抗
性化合物,其对甜菜夜蛾的驱避率达到71%。茉莉酸的生物合成方式为细胞膜上释放的α-亚麻酸在脂氧合酶(LOX2S,EC 1.13.11.12)的催化下合成13(S)-氢过氧-亚麻酸[13(S)-HpOTrE],随后在丙二烯氧化合酶(AOS,EC 4.2.1.92)和丙二烯氧化环化酶(AOC,EC 5.3.99.6)的作用下生成12-氧-植物二烯酸(OPDA),OPDA经12-氧-植物二烯酸还原酶(OPR,EC 1.3.1.42)以及3次β氧化后形成茉莉酸,最后在茉莉酸羧基甲基转移酶(JMT,EC 2.1.1.141)的作用下以茉莉酸为底物生成茉莉酸甲酯。
α-亚麻酸代谢共8个差异表达基因被富集到,包括脂氧合酶基因LOC112191508和丙二烯氧化合成酶基因LOC112173678、LOC112172343、LOC112172188、LOC112173396、LOC112169957,该类基因属于CYP450的74亚家族A2(CYP74A2);12-氧-植物二烯酸还原酶基因LOC112178863以及茉莉酸羧基甲基转移酶基因LOC112174859。
2.4CYP74A2的生物信息学分析
ProtParam理化性质分析结果(表4)表明,5个P450差异表达基因蛋白的分子质量很接近,都在54.3 ku左右。理论等电点预测上,除LOC112173396等电点为6.81外,其他都大于7。5个差异表达的CYP74A2蛋白不稳定性系数都小于40,表明此类蛋白为稳定蛋白。平均亲水系数都小于0,表明该类蛋白为亲水性蛋白。
ProtScale分析結果(图3)也表明,CYP74A2是一类亲水性的蛋白质。跨膜预测和信号肽分析结果表明,该类蛋白无跨膜结构和信号肽。亚细胞定位结果(表4)显示,CYP74A2蛋白主要定位于细胞质和叶绿体中,少量分布于质膜和细胞核内。
SOPMA对CYP74A2蛋白预测的结果(图4)显示,该类蛋白以α-螺旋和无规则卷曲为主,有少量的β-折叠和延伸链。利用SWISS-MODEL对CYP74A2蛋白质进行三维建模,结果(图 5)也表明CYP74A2蛋白主要由α-螺旋和和无规则卷曲构成,这与SOPMA的二级结构预测结果一致。
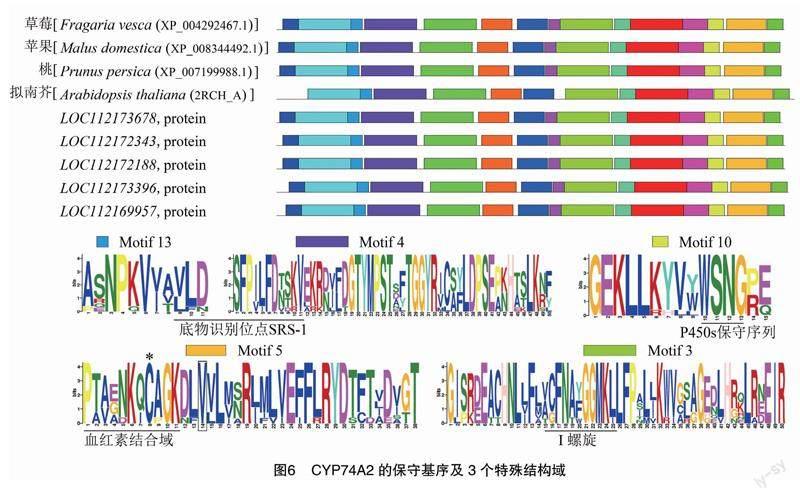
通过NCBI Protein Blast找到月季CYP74A2同源性最高的蛋白,利用MEME对这些蛋白进行保守基序分析,结果(图6)显示,CYP74A2蛋白与苹果、草莓、桃、拟南芥的CYP74A2蛋白有极高的同源性。在基序4和基序13中存在底物识别位点 SRS-1,基序10中“WSNG”的序列是P450s的保守序列。基序5为血红素结合域,用于血红素配体结合的保守半胱氨酸残基用星号表示,该结构域在P450蛋白中普遍存在,其氨基酸序列特征为“PXAXNKQCAG”。基序3中则存在血红素结合域的I型螺旋,氨基酸序列特征为“TCFNAXXGXXXF/L”。
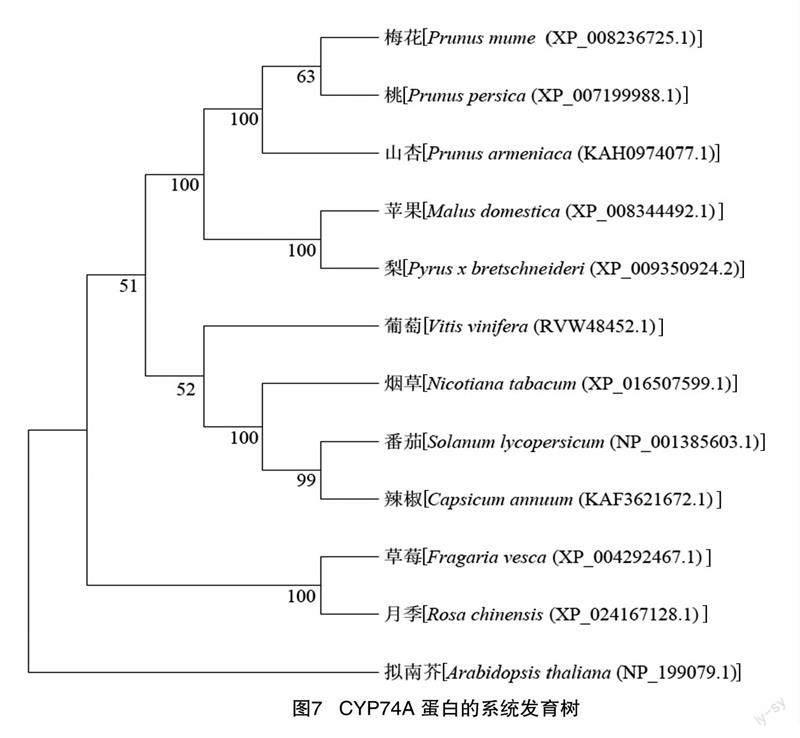
将NCBI Protein Blast得到的同源性最高的11个物种的CYP74A蛋白与测序获得的月季CYP74A2蛋白一起构建系统发育树,结果(图7)表明,月季CYP74A2蛋白与同为蔷薇科的草莓CYP74A蛋白亲缘性最近,与拟南芥CYP74A蛋白亲缘性最远。
2.5CYP74A2差异表达基因的qRT-PC验证
以LOC112165308基因为内参基因,对5个CYP74A2基因进行qRT-PCR验证,结果表明,LOC112173678、LOC112172343、LOC112172188、LOC112169957这4个基因与转录组测序结果一致,虽然LOC112173396表达水平略低于转录组,但其上调的趋势仍然一致(图8)。
3讨论与结论
以中国月季为试验材料,对白粉菌诱导前后的月季进行转录组测序, 随后将鉴定的差异表达基因进行功能富集分析,最后通过生物信息学分析和qRT-PCR验证,明确CYP74A2在分泌月季挥发物中抗甜菜夜蛾的生物分子学机制。
虽然在很多植物尤其是苔藓中CYP74家族有很多报道,且月季的全基因组也已公布[18],但关于月季作为寄主植物介导的病虫害互作关系的生物分子学机制还未被探明,尤其是关键基因在该互作关系下的功能及机制。茉莉酸作为参与应激反应和发育的重要信号分子,同时也参与了对甜菜夜蛾
的驱避。其通路中的丙二烯氧化物合酶基因属于CYP74A家族,该类酶在催化脂肪酸形成挥发性物质过程中不需要氧。CYP74A2可以将氧脂合酶催化的13(S)-氢过氧-亚麻酸转化成不稳定的 12-氧-植物二烯酸,最终经过酶催化转化为茉莉酸,该代谢途径是植物采用的主要防御机制之一。
CYP74A2具有CYP450共有的血红素结合域以及该家族所特有的底物识别位点SRS-1结构域[19-20],有研究表明,CYP74亚家族在氨基酸的 N-末端具有类似质体或线粒体的运送肽[21]。另外,血红素结合域通常位于蛋白质一级结构中的第330~370位氨基酸,有特征性半胱氨酸并包含标志性序列“WSNG”是血红素结合环的一部分[22]。在血红素结合域后的第4个残基上,其他P450s中强烈保守为丙氨酸或甘氨酸,但在CYP74中却是缬氨酸或异亮氨酸,该位置缬氨酸的功能可能是使CYP74的血红素倾斜[23]。
茉莉酸类化合物在植物防御方面发挥了重要的作用,该类化合物的生物合成需要脂氧合酶、丙二烯氧化合酶和丙二烯氧化环化酶的连续作用[16,24]。植物界广泛存在参与JA生物合成的酶,这也反映了植物对具有挑战性的昆虫和病原体的防御反应的高度保守机制。在植物对昆虫和病原体的防御机制中,茉莉酸类化合物被认为是一个关键的组成部分,也有文献表明,氧代植物二烯酸也是激活防御机制的必要条件。因此,植物体内可能已经进化出协调与协同的防御机制[25],JA和OPDA可能具有协同作用[26]。
白粉菌诱导月季合成的茉莉酸不仅调控了植物的新陈代谢,还调控了甜菜夜蛾的寄主选择行为,增强了月季的抗虫性[27]。KEGG功能富集分析和CYP74A2的生物信息学分析结果表明了月季抗虫性产生的分子生物学机制。CYP74A2在白粉菌诱导月季产生对甜菜夜蛾的抗性方面发挥了重要作用。本研究结果可为筛选月季抗甜菜夜蛾的关键基因提供重要理论支撑。但白粉菌诱导月季产生对甜菜夜蛾的影响受多个网络调控,具体调控途径仍需深入研究。
参考文献:
[1]云南省统计局,国家统计局云南调查总队. 云南省2021年国民经济和社会发展统计公报:2022年3月[N]. 云南日报,2022-03-28(5).
[2]云南省农业农村厅,云南省人民政府新闻办公室.“COP15春城之邀”云南生物多样性保护系列新闻发布会暨云南打造“世界花园”专题新闻发布[EB/OL]. (2021-09-14)[2022-07-22]. https://nync.yn.gov.cn/html/2021/hudongjiaoliu-xinwenfabu_0914/380335.html?cid=3035.
[3]Yang F Z,Dong W X,Zhang X G,et al. Volatile-organic compound changes in rose twigs consequent to infection with rose powdery mildew[J]. Chilean Journal of Agricultural Research,2019,79(4):596-608.
[4]楊发忠,杨德强,杨斌,等. 中国月季感染白粉菌对甜菜夜蛾取食行为的影响及原因初探[J]. 广东农业科学,2015,42(17):67-71.
[5]Hu B,Huang H,Hu S Z,et al. Changes in both trans-and cis-regulatory elements mediate insecticide resistance in a lepidopteron pest,Spodoptera exigua[J]. PLoS Genetics,2021,17(3):e1009403.
[6]Karban R,Adamchak R,Schnathorst W C. Induced resistance and interspecific competition between spider mites and a vascular wilt fungus[J]. Science,1987,235(4789):678-680.
[7]Franco F P,Moura D S,Vivanco J M,et al. Plant-insect-pathogen interactions:a naturally complex ménage à trois[J]. Current Opinion in Microbiology,2017,37:54-60.
[8]Grunseich J M,Thompson M N,Aguirre N M,et al. The role of plant-associated microbes in mediating host-plant selection by insect herbivores[J]. Plants,2019,9(1):6.
[9]Bertea C M,Casacci L P,Bonelli S,et al. Chemical,physiological and molecular responses of host plants to lepidopteran egg-laying[J]. Frontiers in Plant Science,2020,10:1768.
[10]李艳,杨发忠,杨斌. 感染白粉菌玫瑰对甜菜夜蛾幼虫取食与发育的影响[J]. 西南林学院学报,2010,30(2):44-46,55.
[11]杨发忠,杨德强,杨斌,等. 白粉菌侵染中国月季对甜菜夜蛾幼虫乙酰胆碱酯酶活性的影响[J]. 河南农业科学,2015,44(12):75-78.
[12]杨发忠,董智森,肖春. 中国月季感染白粉菌后矿质元素种类和含量的变化[J]. 江苏农业科学,2016,44(6):304-306.
[13]汪思远,蒋世翠,王康宇,等. 植物细胞色素P450的研究进展[J]. 吉林蔬菜,2014(4):41-45.
[14]Zhao C Q,Tang T,Feng X Y,et al. Cloning and characterisation of NADPH-dependent cytochrome P450 reductase gene in the cotton bollworm,Helicoverpa armigera[J]. Pest Management Science,2014,70(1):130-139.
[15]Wang Y Q,Liu M F,Ge D D,et al. Hydroperoxide lyase modulates defense response and confers lesion-mimic leaf phenotype in soybean [Glycine max (L.) Merr.][J]. The Plant Journal:for Cell and Molecular Biology,2020,104(5):1315-1333.
[16]陈功锡,田向荣,肖佳伟,等. 中国亚麻酸植物资源[M]. 北京:科学技术文献出版社,2016.
[17]Cheng J,Yin L H,Zhou S P,et al. The inhibitory effect of powdery mildew-induced volatiles from rose on host selection behavior of beet armyworm moths (Lepidoptera:Noctuidae)[J]. Journal of Entomological Science,2022,57(1):96-113.
[18]Raymond O,Gouzy J,Just J,et al. The Rosa genome provides new insights into the domestication of modern roses[J]. Nature Genetics,2018,50(6):772-777.
[19]Toporkova Y Y,Smirnova E O,Mukhtarova L S,et al. Catalysis by allene oxide synthases (CYP74A and CYP74C):alterations by the Phe/Leu mutation at the SRS-1 region[J]. Phytochemistry,2020,169:112152.
[20]Toporkova Y Y,Gogolev Y V,Mukhtarova L S,et al. Determinants governing the CYP74 catalysis:conversion of allene oxide synthase into hydroperoxide lyase by site-directed mutagenesis[J]. FEBS Letters,2008,582(23/24):3423-3428.
[21]宋展,高鑫,吳冕,等. 细胞色素P450酶的结构、功能与应用研究进展[J]. 微生物学通报,2020,47(7):2245-2254.
[22]Koeduka T,Ishizaki K,Mwenda C M,et al. Biochemical characterization of allene oxide synthases from the liverwort Marchantia polymorpha and green microalgae Klebsormidium flaccidum provides insight into the evolutionary divergence of the plant CYP74 family[J]. Planta,2015,242(5):1175-1186.
[23]Brash A R. Mechanistic aspects of CYP74 allene oxide synthases and related cytochrome P450 enzymes[J]. Phytochemistry,2009,70(13/14):1522-1531.
[24]Li C Y,Schilmiller A L,Liu G H,et al. Role of β-oxidation in jasmonate biosynthesis and systemic wound signaling in tomato[J]. The Plant Cell,2005,17(3):971-986.
[25]Hughes R K,De Domenico S,Santino A. Plant cytochrome CYP74 family:biochemical features,endocellular localisation,activation mechanism in plant defence and improvements for industrial applications[J]. Chembiochem,2009,10(7):1122-1133.
[26]Stintzi A,Weber H,Reymond P,et al. Plant defense in the absence of jasmonic acid:the role of cyclopentenones[J]. Proceedings of the National Academy of Sciences of the United States of America,2001,98(22):12837-12842.
[27]Yang F Z,Li Y,Yang B. The inhibitory effects of rose powdery mildew infection on the oviposition behaviour and performance of beet armyworms[J]. Entomologia Experimentalis et Applicata,2013,148(1):39-47.
