竹林扩张进入杉木人工林对土壤磷素的影响
2023-07-05田书荣廖德志肖静晶刘思思黄莞莞
吴 慧,田书荣,廖德志,彭 超,涂 佳,肖静晶,刘思思,黄莞莞,周 诚
(1. 湖南省森林草原防火监测调度评估中心,湖南 长沙 410004;2. 湖南省林业科学院,湖南 长沙 410004;3. 长沙市林业技术推广站,湖南 长沙 410205)
竹类Bambusoidea是我国重要的非木材森林资源[1],主要分布于热带至温带平原、丘陵及高山地区。根据竹林的生理特性,不论是人工竹林还是天然竹林,均有不同程度向四周非竹林扩张的特点。相对被入侵的树种,竹类根系相对生长速率更高[2],渗透能力随植株密度和根系的增加而增加[3]。叶耿平等[4]的研究表明竹林扩张过程使土壤表层水解氮明显提高,有效磷含量显著降低;竹林入侵阔叶林所引起的土壤微生物群落结构改变[5]、土壤纤维素酶和木聚糖酶活力降低[6]等,限制了被入侵树种的正常生长。
红壤是适宜竹类生长的区域地带性土壤,其有机磷含量占全磷的26%~49%[7]。Fink 等[8]研究认为被植物根系直接吸收的有机磷只是一小部分,大部分必须进一步矿化为无机磷才能被植物吸收利用。尤其是在无机磷含量较低的土壤中,有机磷的转化是植物吸收磷素的重要来源[9]。然而,土壤中有机磷的化学组成复杂,与无机磷相比,有机磷的可利用性、动态变化以及转化过程缺乏足够的认识[10-12]。有关有机磷的研究[13]表明,林地立地改变会改变磷形态而显著影响植物对磷的吸收。Chen 等[14]的研究表明植物种类的转变引起植物产生的生物量和养分循环变化,这对土壤性质和养分的有效性产生显著积极或负面的影响。这种转变引起有机磷形态的改变,并因此对磷的有效性和稳定性产生潜在的影响[15]。
杉木Cunninghamia lanceolata是湖南省的主要用材树种之一,是我国民经济发展和森林多种效益的支柱资源。本研究选择一个竹杉混交界面,研究土壤磷素特征,包括有机磷、无机磷和不同形态的磷,比较不同入侵程度土壤磷素有效性分级特征,揭示毛竹林扩张进入周边杉木林引起土壤磷素的动态变化,为杉木林的管理提出合理建议。
1 材料与方法
1.1 研究区概况
试验区位于湖南省浏阳市张坊镇万年林场(114°8′3.597″N,28°20′48.182″E)。该地区为典型的中亚热带季风气候,年平均气温12 ℃,年平均降水量1 700 ~1 800 mm,无霜期243 d。选择的林地为7年以上未受人为干扰的杉木林。该区域母岩由花岗岩和板页岩组成。土壤为黄壤和黄棕壤,土层深度范围为45 cm 以上。
1.2 试验方法
该研究于2019年9月—10月进行。在竹林入侵前各点建立20 ~60 m 的样带,划分为3 个20 m×20 m 的样方,每个样方具有不同的森林类型,分别为纯竹林、竹杉混交林和纯杉木林。为了确定入侵时间,在每个样方中分别建立了毛竹林(A)、70%竹林与30%杉木混交林(B)、50%竹林与50%杉木混交林(C)、30%竹林与70%杉木混交林(D)、纯杉木林(E)5 个采样样地(5 m×20 m)。
1.3 Hedley 磷分级
1982年Hedley[16]提出的磷素分级法,同时考虑了无机和有机磷组分的动态变化,较为合理地反映了土壤磷素分级。
Hedley 分级法利用不同溶剂提取不同形态的磷,根据溶剂不同,将土壤磷分为7 类:
1)树脂交换态磷(Resin-P):利用阴离子交换树脂取代出的土壤磷能被土壤中水或溶剂迅速溶解和补充,这部分磷是土壤可被植物利用的最活跃部分。
2)NaHCO3-P 态磷:包括NaHCO3-Pi(无机磷)和NaHCO3-Po(有机磷)。该部分磷是可溶性有效磷,易于矿化。
3)土壤微生物磷(Microbial-P):这部分磷是微生物死后能被植物利用的磷。提取方法:取两份树脂交换磷,一份氯仿熏蒸后用NaHCO3提取,另一份直接用NaHCO3提取,两者差值即土壤微生物磷。
4)NaOH 溶性磷(NaOH-P):交换出土壤Fe和Al 结合的磷。
5)土壤团聚体内磷:结合在土壤团聚体内表面上的无机和有机磷。
6)HCl 磷:高度风化土壤中(如红壤)提取的部分闭蓄态磷。
7)残留磷(Residual-P):用上述方法不能提取的比较稳定态的磷。
1.4 土壤磷酸酶的测定
采用Tabatabai[17]方法测定磷酸酶活性。土壤营养液的配制方法如下:称取1 g 新鲜的土壤,加上4 mL 的马来酸缓冲对硝基苯磷酸(pH 值6.5)和1 mL 4-硝基酚作为衬底,37 ℃震动孵化1 h。使用分光光度计测定吸光度。磷酸酶活性单位(U)为:1 U 为每g 土壤每1 h 产生对硝基酚的量。
1.5 DNA 提取和实时PCR
按照Power soil DNA Isolation Kit(MoBio Laboratories, Carlsbad, CA)标识的方法提取土壤微生物基因组总DNA[18]。检测土壤总基因组DNA 的纯度和质量。采用PCR 法扩增PhoD基因(参数为:94 ℃5 min,94 ℃30 s,55 ℃30 s,72 ℃30 s,72 ℃10 min)。PhoC和PhoD基因扩增引物分别为PhoC-a-f1(5-CGGCTCCTATCCGTCCGG-3)/PhoCa-r1(5-caacatcgctttgcca. GTG-3)和PhoD-f733(5-tgggaygatcaygargt-3)/PhoDR1083(5-ctgsgcsak sacrttcca-3)。用ABI 7500 型实时荧光定量PCR 仪(美国)进行qPCR 反应。反应体系20 μL 包括10 μL 2×SYBR Premix Ex Taq(Takara,中国),0.4 μL 10 μmol/L 正/反向引物,2 μL cDNA,7.2 μL ddH2O。每个反应3 个复孔,3 次生物学重复。PCR 程序:95 ℃,30 s;95 ℃,5 s; 60 ℃, 30 s,40 个循环。反应结束绘制熔解曲线。根据生成的循环阈值(cycle threshold, Ct),用2-ΔΔCt方法计算相对表达量。
1.6 统计分析
利用IBM SPSS Statistics(Version 19.0)软件(IBM,Armonk,NY,USA)对土壤无机磷、土壤无机磷、土壤酶活性及PhoC和phoD基因丰度数据进行分析。数据采用单因素方差分析,平均值采用Duncan 检验(P<0.05)。结构方程建模使用软件包AMOS。
2 结果与分析
2.1 土壤总无机磷和有机磷的含量
以不同程度竹林扩张进入杉木人工林为研究对象,比较不同扩张程度杉木人工林土壤磷的分布特征。由表1 可知,毛竹林扩张进入杉木林各形态磷总体分布特征表现为:残余磷>HCl 无机磷>NaOH 有机磷>树脂无机磷>团聚体有机磷>NaHCO3无机磷>NaHCO3有机磷>NaOH无机磷>团聚体无机磷>微生物有机磷>微生物无机磷。
竹林扩张进入杉木林的过程显著改变林地土壤无机磷、有机磷和残余磷的组成。30%竹林与70%杉木混交林总无机磷、总有机磷和总磷含量均显著高于其他立地林;纯杉木林残余磷含量显著高于其他立地林。
30%竹林与70%杉木混交林总无机磷含量分别较纯竹林、70%竹林与30%杉木混交林、50%竹林与50%杉木混交林和纯杉木林高30.09%、14.55%、8.25%、7.23%。其中NaHCO3无机磷、微生物无机磷、NaOH 无机磷、团聚体无机磷含量随着竹林扩张入杉木进程逐渐增加,到30%竹林与70%杉木混交时达到顶峰,最终在纯杉木林时较30%竹林与70%杉木混交时显著下降。纯竹林HCl 无机磷含量与纯杉木林70%竹林与30%杉木混交林、50%竹林与50%杉木混交林、30%竹林与70%杉木混交林差异显著,而70%竹林与30%杉木混交林、50%竹林与50%杉木混交林、30%竹林与70%杉木混交林、纯杉木林混交林之间差异不显著。
30%竹林与70%杉木混交林总有机磷含量分别较纯竹林、70%竹林与30%杉木混交林、50%竹林与50%杉木混交林和纯杉木林高30.09%、14.55%、8.25%、7.23%。其中NaHCO3无机磷和NaOH 有机磷含量随着竹林扩张入杉木进程逐渐增加,到30%竹林与70%杉木混交时达到顶峰,最终在纯杉木林时较30%竹林与70%杉木混交时显著下降。微生物有机磷含量随着竹林扩张入杉木林显著增加,到纯杉木林时含量达到峰值。团聚体有机磷在70%竹林与30%杉木混交林、50%竹林与50%杉木混交林和纯杉木林间差异不显著。
2.2 磷酸酶活性
由图1 可知,竹林扩张进入杉木林的过程显著改变了林地土壤磷酸酶含量(P<0.05)。其中,30%竹林与70%杉木混交林土壤磷酸酶含量显著高于其他立地人工林,且随着竹林扩张进入杉木林的进程,土壤磷酸酶含量逐渐增加,到30%竹林与70%杉木混交林时达到顶峰,纯杉木林时下降。
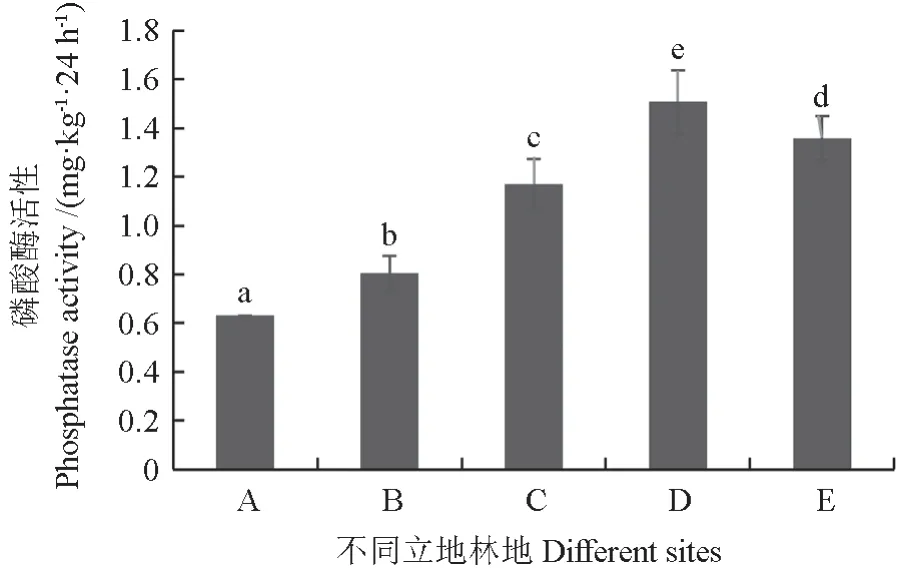
图1 磷酸酶活性Fig. 1 Phosphatase activity

图2 PhoC 和PhoD 基因丰度Fig. 2 The gene abundance of PhoC and PhoD
2.3 PhoC 和PhoD 基因丰度
竹林扩张进入杉木林的过程中,各个阶段土壤样本提取的DNA,PhoC基因丰度均显著高于PhoD基因丰度。其中,30%竹林与70%杉木混交林土壤PhoC基因和PhoD基因丰度显著高于其他立地类型,且随着竹林扩张进入杉木林的过程进展,PhoC基因和PhoD基因丰度显著增加,在30%竹林与70%杉木混交林时PhoC基因和PhoD基因丰度达到峰值,分别为(5.25±0.28)copies·g-1和(3.71±0.13)copies·g-1,随后在纯杉木林时显著下降。
2.4 相关性分析
竹林扩张对磷酸酶、有机磷和无机磷均产生了负面影响。根据方差分析和结构方程模型分析,竹林扩张进程直接和间接驱动土壤磷酸酶、有机磷和无机磷的变化,主要是通过影响土壤磷酸酶进而影响土壤有机磷,而土壤有机磷又驱动土壤无机磷的变化(图3)。卡方检验MIN/DF=3.759<5,各标准化路径系数均显著(P<0.05)。磷酸酶因子中,酸性磷酸酶活性负载最大(0.95),其次是PhoD和PhoC基因,分别为0.75 和0.67。无机磷因子中,树脂无机磷因子负荷最大(0.94),其次是微生物无机磷、NaHCO3无机磷、NaOH 无机磷、HCl无机磷、团聚体无机磷、总无机磷,分别为0.92、0.91、0.89、0.88、0.86 和0.66。有机磷因子中,微生物有机磷负载最大(1.11),其次是残余磷、总有机磷、NaHCO3、NaOH 有机磷,分别为0.85、0.83、0.77、0.74、0.71。

图3 竹林扩张进入杉木林土壤磷素组成的结构方程模型Fig. 3 The structural equation model of soil phosphorus composition during the expansion of bamboo forest intoCunninghamia lanceolata forest
3 讨 论
毛竹林扩张进入杉木林各形态磷总体分布特征表现为:残余磷>HCl 无机磷>NaOH 有机磷>树脂无机磷>团聚体有机磷>NaHCO3无机磷>NaHCO3有机磷>NaOH 无机磷>团聚体无机磷>微生物有机磷>微生物无机磷。因此,具有高有效性的磷,随着竹林扩张进展逐渐降低,而有机磷与具有功能潜力的磷分的动态有关,并随竹林扩张进展呈现下降趋势。值得注意的是,考虑到磷对植物的有效性,土壤样品中无机磷随着竹林扩张的进展逐渐降低,竹林扩张程度高无机磷含量的降低可能与植物对土壤养分的竞争有关[19],因为有研究表明,竹林扩张程度高一般对有效土壤资源具有较高的种内竞争[20]。再加上溶解的微生物细胞是磷释放的来源,可能是纯杉木林无机磷浓度高的原因。此外,磷的有效性受到植物根系的影响,因为土壤中有机磷的主要形式是通过酯键与有机质结合[21-23]。在本研究中,无机磷的富集可能与竹林扩展进程低的根际效应增加有关,因为竹林扩展进程低通常会降低森林生态系统的透光性和光合效率[24],这导致根系分泌物和地下光合产物分配的下降[25]。
通过将土壤磷分离成以其提取方式为特征的组分,并使用温和的试剂去除较容易提取的组分,就有可能识别出竹林扩张进程而改变的土壤磷组分。随着人工林的发展,磷酸酶活性和PhoC、PhoD基因丰度动态明显,其变化趋势由竹林扩张进度进展逐渐转变。这充分说明磷酸酶活性和PhoC、PhoD是土壤有机磷转变为无机磷的敏感指标,与竹林扩张进入杉木林程度有关。土壤磷酸酶活性主要由微生物基因主导[26],因为在杉木的人工林中,磷酸酶活性与PhoC和PhoD基因丰度显著相关。微生物促进的磷酸酶的产生通常会导致有机磷的耗用和矿化[27],本研究中磷酸酶活性与无机磷和有机磷呈显著负相关,磷酸酶活性会随着土壤有效磷的耗竭而增加,正如Garg 等[28]研究表明低磷酸酶活性与低速效磷含量有关。这进一步证明PhoC和PhoD基因丰度的降低,降低了磷酸酶的产量[29]。本研究结果表明,有机磷在纯杉木人工林中是稳定的,显示竹林扩张进入杉木林需要干预,在竹林有趋势进入杉木林时就需要干预。
土壤活性磷(树脂无机磷、NaHCO3无机磷、NaHCO3有机磷)和中度活性磷(HCl 无机磷、NaOH 无机磷、NaOH 有机磷)是土壤供磷能力和生物有效性的主要指标[30],而残磷被认为是非活性磷,对植物无效。在本研究中,树脂无机磷、NaHCO3无机磷、NaHCO3有机磷含量在竹林扩张前期低,说明磷的生物有效性随着扩张的进行而降低。NaHCO3有机磷和NaOH 有机磷会随着扩张进程而下降,因为它们是植物有效磷的主要来源。造成这一结果的主要原因有两个:1)酸性条件下,有机磷的主要形式是由土壤有机质与Fe、Al、Ca、Mg 降解产生的有机复合体键吸附而成;2)微生物将有机磷转化为无机磷在磷循环中起着重要作用。我们的推论得到了一些早期研究的支持,这表明矿化和分解的有机磷被认为是由有机质和土壤中微生物的活动所控制。磷供应能力由NaHCO3有机磷和NaOH 有机磷分数随着竹林扩张的进行而降低了磷的生物有效性,是因为纯杉木林较竹林扩张进入杉木林林地密度低,纯杉木林微生物的转化能力高于扩张阶段的混合林。
随着竹林扩张的进展磷酸酶活性、PhoC和PhoD基因丰度显著降低,林地密度由低密度逐渐向高密度转变。这充分说明微生物群落和酶是土壤微气候变化的敏感指标,其动态与竹林扩张阶段有关。土壤磷酸酶活性主要由微生物基因主导,在纯杉木林中,磷酸酶活性与PhoC和PhoD基因丰度显著相关。而在竹林中,磷酸酶活性与PhoC基因丰度呈显著正相关,因为pH 值是影响微生物群落的一个主要变量。土壤酸化,微生物群落可能向具有PhoC基因的生物转移。
微生物促进的磷酸酶的产生通常会导致有机磷的消耗和矿化,这与本研究中磷酸酶活性与NaHCO3有机磷和NaOH 有机磷显著负相关一致。尽管之前研究显示,随着土壤速效磷的消耗,预计磷酸酶活性会增加,竹林扩张程度高时酸性磷酸酶活性低与速效磷含量低相关。这进一步表明,PhoC和PhoD基因丰度的降低会降低磷酸酶的产生。此外,磷酸酶活性的降低可能与低浓度的易降解底物有关,如磷酸单酯[27]。研究表明,纯杉木林有机磷非常稳定,基因丰度的增加可能与微生物群落丰富度和多样性的增加有关。根系生物量的增加会刺激根系分泌物进入土壤,促进微生物群落的丰富度和多样性。这可以解释30%竹林与70%杉木混交林土壤中PhoC和PhoD基因丰度显著高于其他立地。如果PhoC和PhoD基因丰度的增加是促进根系生物量增加,而导致微生物富集,可以认为在竹林扩张过程中30%竹林与70%杉木混交林也是因为根系生物量的增加而微生物富集。人工林的PhoC和PhoD基因丰度表明在竹林扩张进度发展过程中,根系分泌物会随着竹林扩张进展的增加而减少。如果PhoC和PhoD基因的丰度及其表达依赖于有效磷的消耗,则在高度扩张环境中可以观察到PhoC和PhoD基因丰度和磷酸酶活性的增强。结果表明,磷酸酶活性及PhoC和PhoD基因丰度与NaHCO3无机磷和NaOH 无机磷呈显著正相关。由于扩张程度对微生物群落的结构、代谢功能、生物量和多样性有显著影响,因此推测,PhoC和PhoD环境下微生物群落的结构可能会受到种植密度的影响。这表明,扩张进程不仅降低了磷的生物有效性,而且还削弱了微生物群落驱动的有机磷向无机磷转化的生化过程,因为微生物通过增加磷酸酶产量来应对磷缺乏。同时,其他影响PhoC和PhoD微生物群落结构的因素也被提出,如氧化应激、pH 值和矿质养分的组合等。
4 结 论
在酸性土壤中,种植密度的降低有望提高土壤磷的生物有效性。虽然预计随着土壤有效磷的耗尽,磷酸酶活性和PhoC和PhoD基因的丰度会增加,但高密度人工林的低磷酸酶活性和PhoC、PhoD基因的丰度与有效磷含量低有关。这意味着缺磷土壤相比不缺磷土壤,微生物对磷循环的抑制作用更强。研究结果表明,为保证土壤肥力和养分循环的有效性,应在合适的时机限制竹林扩张进入杉木林。本研究还观察到磷酸酶活性与PhoC、PhoD基因丰度之间的显著相关性,从而提高了对磷循环中重要生物过程的理解,以及竹林扩张土壤微生物群落如何驱动磷酸酶活性。
