马铃薯套种苦荞对土壤养分、酶活性和微生物的影响
2023-06-17刘辉卢扬冯月玲叶夕苗张叶李飞邓仁菊张廷刚王天顺宋莉
刘辉 卢扬 冯月玲 叶夕苗 张叶 李飞 邓仁菊 张廷刚 王天顺 宋莉

摘要:为分析马铃薯套种苦荞下不同时期土壤中的养分、土壤酶活性和微生物生态在苦荞不同生长时期的变化。结果表明,与马铃薯净作区相比,马铃薯套种苦荞区的土壤pH值在苦荞各个生长时期整体均显著上升。在马铃薯净作区,与2020年相比,2021年不同生长时期各土壤营养指标、土壤酶活性具有显著差异,但是对于马铃薯套作苦荞区,不同生长时期的土壤营养指标和土壤酶活性无显著差异,表明套作苦荞可显著缓解马铃薯连作对土壤的影响。在2020年,与净作区相比,马铃薯套作苦荞区在幼苗期的土壤全碳、速效磷含量显著下降,有机质含量显著上升。在苦荞开花期,土壤全碳、全氮、全磷、碱解氮磷含量显著降低,全钾和有机质含量则显著上升;在苦荞成熟期,土壤全碳、全氮、全磷、碱解氮含量显著下降,全钾、速效钾、速效磷含量则无显著变化,有机质含量显著上升;在苦荞幼苗期、开花期与成熟期,土壤脱氢酶活性均显著上升,其他土壤酶活性的变化均不显著;2021年各指标的变化趋势与2020年一致。由结果可知,幼苗期的苦荞需要从土壤中吸收部分碳肥、磷肥,开花期的苦荞需要从土壤中吸收部分碳肥、氮肥、磷肥,成熟期的苦荞需要从土壤中吸收部分碳肥、氮肥。但在上述3个时期,土壤脱氢酶活性的提升可以促进有机质代谢,从而提升有机质含量。另一方面,马铃薯套作苦荞可以降低真菌总数、提升细菌总数,尤其是部分具有生防作用的放线菌总数。综上,苦荞套作模式可稳定土壤pH值,促进苦荞合理利用土壤中的营养成分,并提升有机质、钾肥含量,同时改善土壤微生物生态系统,降低病害发生率,以缓解马铃薯连作对土壤的影响,因而对马铃薯、苦荞而言均是有益的选择。
关键词:马铃薯;套作;苦荞;土壤养分;土壤酶;土壤微生物
中图分类号:S532.04;S532.06文献标志码:A
文章编号:1002-1302(2023)10-0219-08
马铃薯是世界第四大粮食作物,也是重要的粮菜兼用作物和工业原料[1]。自2000年以来,我国马铃薯种植面积逐年增加,已经成为世界上马铃薯第一大生产国,播种面积和产量均居世界首位[2]。大面积单作马铃薯的种植模式改变了传统的轮作制度,使得马铃薯与其他作物轮作倒茬困难。此外,马铃薯重茬、迎茬种植、轮作倒茬年限不够等的问题也愈加突出[2]。套作利用作物对养分吸收利用的不同生物学特征而形成的不同养分吸收规律,使同一地块上的作物形成时间、空间上的互补性,可从根本上解决作物养分吸收的不平衡,改善群体微生态环境,更有利于农作物的生长发育,从而达到农作物增产增效的目的[3]。因此,合理发展套作技术对解决马铃薯连作障碍、提升产量和土地利用率具有重要作用。
贵州省毕节市威宁县地处乌蒙山区,属亚热带季风性湿润气候,无霜期长,雨热同期,是中晚熟马铃薯的主产区。马铃薯生长后期病虫害高发,地上植株非正常死亡后,多生长杂草,降低了土地利用率[4]。为了解决这一难题,应该合理应用马铃薯的间作套种技术[5]。王海燕等研究发现,马铃薯套种蚕豆虽然会造成单位面积土地上的作物品种增加、密度上升,但其形成的复合生态体系可显著改善蚕豆条带内的透光率,同时提高马铃薯、蚕豆对光能的利用率,从而达到作物增产、増收、增效的目的[6]。苦荞含有生物活性物质如黄酮类物质、γ-氨基丁酸、D-手性肌醇等,具有较强的抗氧化活性,同时含有超过60%的能量物质——淀粉,因而是一种重要的药食两用作物,在西南地区的四川省、云南省、贵州省等地,大面积种植了苦荞,而苦荞在饥荒年代也是一种重要的互补性粮食作物[7-8]。目前,在实践和理论方面已有苦荞套作的报道。如龙梦千等发现,苦荞‖马铃薯间作能够促进苦荞生长、延缓苦荞衰老、提高光合速率、增加籽粒质量,达到增产的效果[9]。毛春等将苦荞套作于马铃薯地块中发现,套作对马铃薯产量无明显影响,还能额外增加苦荞产量和收益[10]。汤睿等在春、秋2季开展油茶/苦荞、茶树/苦荞、猕猴桃/苦荞3种套种模式,其中猕猴桃/苦荞套种模式的经济效益最高[11]。马铃薯套作苦荞是贵州省2季作区采用的栽培模式,但是目前关于这种栽培模式对土壤理化性质变化的影响鲜见报道。本研究设置马铃薯净作、马铃薯套作2种栽培方式,分别于苦荞幼苗期、开花期、成熟期测定农田土壤的养分含量、pH值和土壤酶活性。本研究拟解决的关键问题在于通过从土肥领域探索马铃薯套作苦荞的深层次机制,旨在为马铃薯套作苦荞的应用推广提供理论指导。
1材料与方法
1.1试验区概况
试验地址在贵州省毕节市威宁县双龙镇江林村,海拔2224m,地理坐标为104.28°E、26.87°N,属于亚热带季风性湿润气候区,具有低纬度的高原季风气候和山地气候特征。年均气温为10~13℃,年均降水量为925mm,无霜期约250d。试验地块无坡度,土壤类型为黄棕壤。
1.2试验设计
田间种植试验于2020年3月至2021年11月进行。试验采用单因素随机区组设计,设2个处理,分别为马铃薯净作(CK)和马铃薯套作苦荞,每个处理的面积为672m2(24m×28m)。马铃薯播前牛耕1次,人工精细整地。马铃薯品种为黔薯7号,苦荞品种为黔苦7号(均由贵州省生物技术研究所提供)。2个处理马铃薯的播种时间相同,于2020年3月种植,播种量为2250kg/hm2,按常规种植,行距7.0cm,株距3.0cm。马铃薯施用375kg/hm2磷肥、225kg/hm2复合肥、22.5t/hm2农家肥,混合拌匀后用作基肥。现蕾期结合第1次中耕培土追施225kg/hm2氮肥;苦荞的播种期根据马铃薯的田间长势确定,原则上在马铃薯成熟期前,基部叶片变黄时,在用耙子除去行间杂草的同时松土,再将苦荞种子均匀撒播在空隙地,等苦荞收获后再收马铃薯。苦荞于2020年7月中旬种植,播种量为67.5kg/hm2,采用人工撒播方式播种。2021年重复上述試验1次,马铃薯和苦荞的种植、收获时间与2020年一致。
1.3土壤样品的采集
分别于苦荞幼苗期、开花期、成熟期,在每个处理内按“Z”字形选取采样点,取5~30cm土层土壤作为样品。每个处理取5个点的样品均匀混合后作为1个土样,剔除样品中的断根、石块等杂质,自然风干后过1mm土筛。土壤样品分为2份,1份用于土壤速效养分含量、pH值及酶活性的测定,另1份保存于4℃冰箱中备用。设T1处理为幼苗期马铃薯净作的土样,T2处理为幼苗期马铃薯套作苦荞的土样,T3处理为开花期马铃薯净作的土样,T4处理为开花期马铃薯套作苦荞的土样,T5处理为成熟期马铃薯净作的土样,T6处理为成熟期马铃薯套作苦荞的土样。
1.4测定项目及方法
1.4.1土壤pH值和养分的测定土壤pH值的测定参照王玉琴等的方法[12],以无CO2的超纯水作为浸提液,按土液比1g∶1.5mL均匀搅拌1min,于4000r/min离心2min后取上清液,用pH计测定酸碱度。
土壤养分指标包括土壤干湿比、全碳含量、全氮含量、全磷含量、全钾含量、速效磷含量、速效钾含量、碱解氮含量、有机质含量。全碳含量的测定参照HJ695—2014《土壤有机碳的测定燃烧氧化-非分散红外法》,全氮含量的测定参照LY/T1228—2015《森林土壤氮的测定》,全磷含量的测定参照GB/T9837—1988《土壤全磷测定法》,全钾含量的测定参照NY/T87—1988《土壤全钾测定法》,碱解氮含量的测定参照LY/T1229—1999《森林土壤水解性氮的测定》,速效磷含量的测定参照NY/T1121.7—2014《土壤检测第7部分:土壤有效磷的测定》,速效钾含量的测定参照NY/T889—2004《土壤速效钾和缓效钾含量的测定》,土壤有机质含量采用重铬酸钾容量法进行测定。
1.4.2土壤酶活性的测定土壤酶包括土壤脲酶(soilsurease,Ure)、土壤蔗糖酶(soilsaccharase,Sac)、土壤酸性磷酸酶(soilacidphosphatase,ACP)、土壤硝酸还原酶(soilnitratereductase,NR)、土壤脱氢酶(soildehydrogenase,DHA)活性,分别采用苯酚钠-次氯酸钠比色法、3,5-二硝基水杨酸(3,5-dinitrosalicylicacid,DNS)比色法、磷酸苯二钠比色法、2,3,5-氯化三苯基四唑(triphenyltetrazolechloride,TTC)比色法、S-NR活体法测定其活性[13]。
1.5土壤微生物类群的测定
用平板稀释法对细菌、真菌和放射菌进行计数,具体采用稀释涂抹平板法。对土壤溶液设置3个稀释梯度(10-3、10-5、10-6),分别吸取200μL土壤悬液涂在细菌固体培养基、真菌固体培养基和放线菌固体培养基上,置于30℃恒温培养箱中培养3d后计数。具体参照周华兰等的方法[1]。
1.6数据处理
用Origin2018进行数据整理和绘图,用Genstat19.0进行显著性分析,用最小显著性差异法(LSD)进行处理间的差异显著性多重比较(α=0.05),用DPS7.0进行指标间的相关性分析。
2结果与分析
2.1不同时期的土壤pH值和养分含量
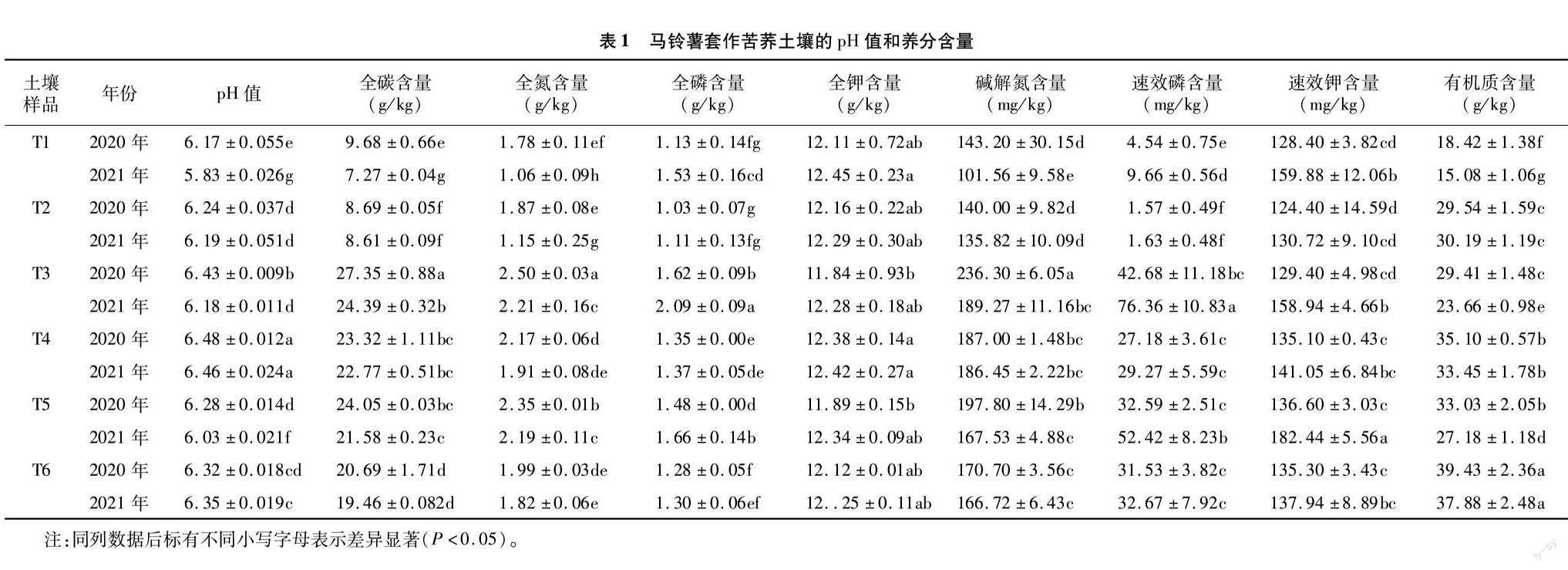
由表1可知,在马铃薯净作的土地上,2021年的土壤pH值相较2020年均显著下降,在苦荞幼苗期、开花期、成熟期分别降低了5.51%、3.89%、3.98%。但在套作区,套作苦荞后的土壤pH值在同一年相比净作区整体显著升高,2020年与2021年相比无显著差异。
从各种营养元素的变化情况来看,在同一年份,苦荞幼苗期、开花期和成熟期的土壤全碳、全氮、全磷、全钾和碱解氮、速效磷含量整体均呈现先升高后降低的趋势,排序为开花期>成熟期>幼苗期;速效钾含量均呈现持续增加的趋势。在2020年,马铃薯套作苦荞与马铃薯净作相比,全碳含量在幼苗期、开花期、成熟期分别降低了10.23%、14.73%、13.97%,且差异均达到显著水平;全氮含量在幼苗期无显著差异,而在开花期、成熟期则分别显著下降了13.20%、15.32%;碱解氮含量在幼苗期无显著变化,开花期、成熟期则分别显著下降了20.86%、13.70%;全磷含量在幼苗期无显著差异,但呈下降趋势,在开花期、成熟期则分别显著下降了16.67%、13.51%;速效磷含量在幼苗期显著降低了65.42%,而在开花期、成熟期无显著差异;全钾、速效钾含量在幼苗期、开花期和成熟期均无显著差异;有机质含量在幼苗期、开花期、成熟期分别显著上升了60.37%、19.35%、19.38%。2021年的变化趋势与2020年的一致,但在净作区,2021年的土壤全碳、全氮、碱解氮含量相对2020年显著降低,而全磷、速效磷、速效钾含量显著上升,全钾含量差异不显著。在套作区套作苦荞后,除全氮含量幼苗期在2021年显著低于2020年外,所有土壤的营养指标2021年与2020年相比无显著差异。
土壤养分与酶活性之间的相关性分析结果表明,在连坐土壤中,蔗糖酶活性与全碳、速效钾含量极显著正相关,而与速效磷极显著负相关;磷酸酶活性与pH值、速效磷、速效钾含量极显著正相关;脲酶活性与速效磷含量极显著负相关,而与速效钾含量显著正相关;硝酸还原酶活性与碱解氮含量、有机质含量、细菌数量显著或极显著正相关,而与速效磷和真菌数量显著或极显著负相关;脱氢酶活性与碱解氮、速效钾、有机质、细菌数量显著或极显著正相关,而与速效磷含量、真菌数量显著或极显著负相关(表2)。
在套作土壤中,蔗糖酶活性与全碳含量、碱解氮含量、速效钾含量、有机质含量、细菌数量、放线菌数量显著或极显著正相关,而与真菌数量极显著负相关;磷酸酶活性与速效磷、有机质含量显著或极显著正相关;脲酶活性与碱解氮含量、有机质含量、放线菌数量显著或极显著正相关;硝酸还原酶活性与pH值、碱解氮含量、有机质含量、细菌数量、放线菌数量显著或极显著正相关;脱氢酶活性与pH值、碱解氮含量、速效钾含量、有机质含量、细菌数量、放線菌数量显著或极显著正相关,而与真菌数量极显著负相关(表2)。
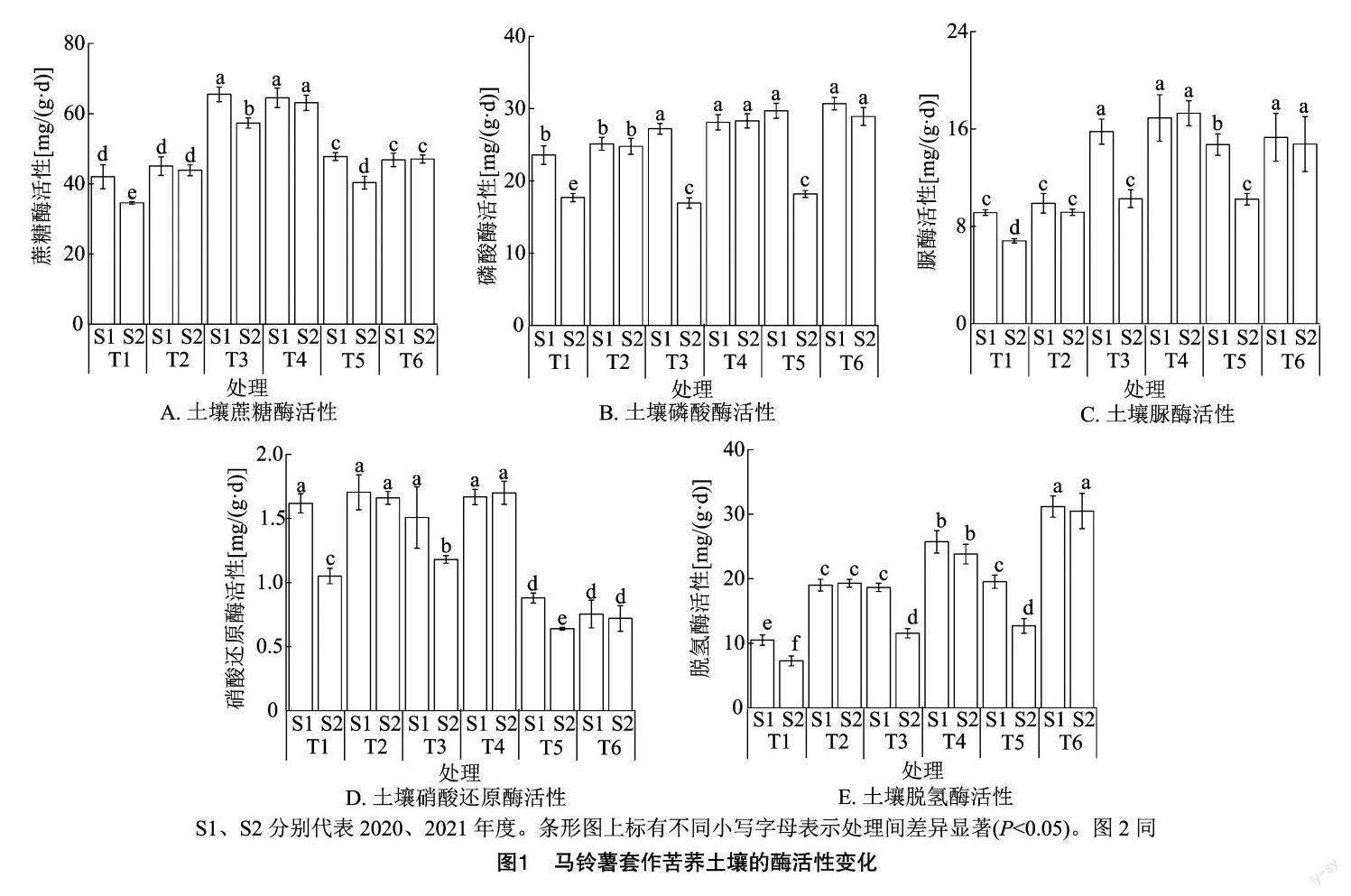
糖酶活性相对净作区显著上升,但在各个生长时期,2021年马铃薯净作区的土壤蔗糖酶活性要显著低于2020年。套作苦荞后,土壤蔗糖酶活性相对净作显著升高,恢复至连作之前的水平,重复套作的第2年(2021年)的酶活性与套作区的2020年数据差异不显著(图1-A)。除此以外,土壤磷酸酶、脲酶、硝酸还原酶活性的变化趋势与土壤蔗糖酶活性一致(图1-B、图1-C、图1-D)。而对于土壤脱氢酶活性而言,在2020年的马铃薯幼苗期,马铃薯套作苦荞的土壤脱氢酶活性比马铃薯净作处理显著提高了81.22%,在开花期、成熟期则分别显著提高了37.83%、59.70%。在2021年的马铃薯幼苗期、开花期和成熟期,马铃薯套作苦荞处理的土壤脱氢酶活性比马铃薯净作处理分别显著提升165.84%、106.33%、140.19%;在各个生长时期,2021年马铃薯净作区的土壤脱氢酶活性显著低于2020年,但套作苦荞后,土壤脱氢酶活性显著提高,与2020年套作苦荞区各时期相比无显著差异(图1-E)。
2.3不同时期土壤中的微生物含量
由图2可知,与马铃薯净作区域相比,马铃薯套作苦荞区域的马铃薯幼苗期、开花期、成熟期的细菌总数、放线菌总数显著上升,其中在2020年,细菌总数分别增加了36.88%、91.85%、188.74%,2021年则分别增加了58.68%、173.12%、242.47%;2020年的放线菌总数分别增加了26.01%、75.77%、193.49%,2021年的放线菌总数则分别增加了79.31%、153.25%、250.34%;真菌总数在2020年分别减少了43.98%、28.20%、50.66%,2021年则分别减少了53.78%、51.69%、61.46%;真菌总数/细菌总数在2020年分别下降了58.57%、62.35%、83.01%,在2021年则分别下降了70.07%、82.18%、88.84%。在马铃薯净作区,与2020年相比,2021年马铃薯幼苗期、开花期、成熟期的细菌总数分别显著减少18.22%、28.95%、17.70%,放线菌总数则分别减少32.19%、27.78%、20.56%,真菌总数分别增加了42.51%、46.33%、34.74%,真菌总数/细菌总数分别显著上升了72.86%、104.71%、64.05%;在马铃薯套作苦荞区,2021年和2020年的细菌总数和放线菌总数在不同生长时期均无显著差异;幼苗期的真菌总数在2021年显著高于2020年,开花期和成熟期的真菌总数无显著性差异。
3讨论
有限的土地资源使得马铃薯连年栽培现象普遍发生,但是多年连作使得马铃薯连作障碍现象发生较为严重,导致土壤pH值下降、根际土壤微生物群落失衡,尤其是镰刀菌、枯萎病病原菌、黑痣病病原菌等致病菌增加,同时土壤蔗糖酶、脲酶等代谢酶活性下降,最终导致病虫害发生频繁,土壤营养失衡,严重影响马铃薯的产量和品质[1,14-16]。研究发现,马铃薯套作苦荞,在马铃薯的行间撒播苦荞,利用马铃薯枯死的残留叶片的肥效和土壤肥料残留可以少施无机肥,防止对土壤造成危害,一方面可以持续利用耕田,维护生态环境,避免连作马铃薯造成品种退化和有害物质的大量残留。另一方面,可以保证有足够多的土地播种苦荞,投入少产量高[10]。因此,马铃薯套作苦荞是解决马铃薯连作障碍的一个有效方法,本研究重点探讨其中的机制。
本研究中,在马铃薯净作区,2021年的土壤pH值显著低于2020年,而马铃薯套作苦荞区,2021年与2020年的土壤pH值差异不显著,且在幼苗期、开花期、成熟期,套作苦荞以后的pH值显著高于当年的净作区。结果表明,马铃薯连作可导致土壤酸化,这与周华兰等的研究结果[1]一致。套作苦荞则可稳定土壤酸碱度,缓解马铃薯连作对土壤pH值的影响。
大量研究表明,马铃薯连作会导致土壤酶(蔗糖酶、脲酶、蛋白酶、过氧化氢酶、磷酸酶等)活性降低,最终引起受土壤酶调控的各种营养指标的变化,从而使马铃薯产量和品质降低,连作障碍因此而形成[17-18]。土壤速效养分含量是衡量土壤肥力的重要指标[19]。土壤酶是土壤肥力密切相关的量,参与土壤中的物质转化、养分固定及矿化过程[20]。蔗糖酶是土壤碳营养分解和转化的重要土壤酶,其活性高低可指示土壤的生化活性[12]。土壤磷酸酶参与了土壤中有机磷转化为无机磷过程,是其中的关键酶[13]。脲酶可促使土壤中尿素转化为氮素,供作物生长所需[21]。土壤硝酸还原酶是一种将土壤中硝酸盐还原为亚硝酸盐的氧化还原酶,是土壤硝态氮还原的关键酶,可作为作物生育期氮素代谢的生理指标[22]。脱氢酶是存在于活体微生物中可催化底物去除氢原子的一种土壤酶,是有机质分解过程中所需的第一酶,可以反映土壤体系中微生物的数量及其对有机物的降解能力[23]。作物之间的套作对土壤酶活性和土壤营养成分具有改善作用,如秦伟等研究发现,设施番茄在套种大蒜以后,土壤脲酶、磷酸酶和蔗糖酶等的活性显著增强,同时碱解氮和速效磷含量因此而得到显著提升[16]。但不同套作模式对土壤酶活性的作用结果不尽相同,例如小麦套作大豆可显著增强土壤磷酸酶活性,而玉米间作鹰嘴豆则可显著降低土壤脲酶和磷酸酶活性[24]。
本研究表明,2021年与2020年相比,马铃薯净作区的土壤全碳、全氮、碱解氮、有机质含量在各个生长时期整体均显著降低,全磷、速效磷、速效钾含量则显著上升,全钾含量无显著变化;马铃薯套作苦荞以后,不同生长时期的土壤营养指标均无显著性差异,表明套作苦荞可显著缓解马铃薯连作障碍。在2020年,马铃薯套作苦荞区与净作区相比,幼苗期的土壤全氮、全磷、全钾、碱解氮、速效钾含量无显著差异,全碳和速效磷含量显著下降,有机质含量显著上升,同时期的土壤蔗糖酶、磷酸酶、脲酶、硝酸还原酶无显著性差异,脫氢酶活性则显著上升,与相对应的土壤营养指标变化一致。结果表明,幼苗期苦荞需要从土壤中吸收部分碳肥和磷肥,但同时可以提升土壤脱氢酶活性,以促进有机质代谢,从而提升有机质含量;开花期的土壤全碳、全氮、全磷、碱解氮和速效磷含量显著降低,全钾、速效钾和有机质含量则显著上升,与这些指标相对应的土壤蔗糖酶、磷酸酶、脲酶、硝酸还原酶活性无显著性差异,脱氢酶活性则显著上升,表明苦荞在开花期需要从土壤中吸收部分碳肥、氮肥、磷肥,但可有效提升土壤钾肥和有机质含量;在苦荞成熟期,土壤全碳、全氮、全磷、碱解氮含量显著下降,全钾、速效钾、速效磷含量则无显著变化,有机质含量显著上升,相对应的土壤蔗糖酶、磷酸酶、硝酸还原酶无显著性差异,脱氢酶活性则显著上升,表明在成熟期,苦荞需要从土壤中吸收部分碳肥、氮肥和磷肥,对钾肥需求不明显,但可显著提升土壤有机质含量;2021年的数据变化与2020年一致。结果表明随着苦荞的生长,对土壤中的磷肥、氮素、碳素的需求增加,其中幼苗期需要碳肥和磷肥,开花期需要更多的碳肥、氮肥、磷肥,成熟期需要更多的碳素和氮素,这与苦荞不同生长时期对营养的需求一致[25]。马铃薯套作苦荞可提升土壤有机质和钾肥含量,对马铃薯连作土壤有一定的改善作用。
研究发现,合理间、套作能改善连作导致的土壤微生物群落结构改变,并增加土壤微生物功能多样性,可在一定程度上稳定土壤微生态环境[26-29]。如研究发现玉米间作花生可显著提升土壤细菌和真菌的生物量,花生根系分泌物有助于根际土壤中革兰氏阳性菌和丛枝菌根的累积[30]。马铃薯、玉米间作增强了土壤中以羧酸类、氨基酸类、多聚类、芳香类化合物为碳源的土壤微生物代谢活动,对土壤微生态环境和土壤微生物功能多样性具有较大的改进作用[31]。本研究中,马铃薯套作苦荞,可显著提升开花期和成熟期的细菌总数,尤其是放线菌总数显著提升,而真菌总数则显著下降,提升了细菌总数/真菌总数比值。由于马铃薯连作可导致土壤中一些真菌性病害如枯萎病、黑痣病、晚疫病、早疫病等病原菌增加的概率大增,而一些具有促生和抗病作用的生防细菌尤其是放线菌数量下降[1]。马铃薯套作苦荞对土壤细菌和真菌数量的影响结果表明该栽培模式可有效缓解马铃薯连作土壤微生物由细菌型向真菌型转化,这与马铃薯间作蚕豆、荞麦的研究结果[32]一致。
前人研究表明,土壤微生物数量、土壤营养成分含量及土壤酶活性紧密相关,如土壤脲酶、蔗糖酶、过氧化氢酶活性与土壤微生物数量、有机质和速效氮含量显著正相关,但与pH值则显著负相关[33]。大棚内茄子套种大蒜,大蒜可通过根系分泌纤维素、糖类等养分,加强根际有益微生物的代谢活动,间接提高土壤脲酶和磷酸酶活性,从而促进土壤氮素的转化和有机磷的矿化,最终为茄子丰产和连作障碍的改善奠定基础[16]。本研究结果表明,连作土壤中蔗糖酶活性与全碳、速效钾含量极显著正相关,与速效磷含量极显著负相关。磷酸酶活性与pH值、速效磷和速效钾含量极显著正相关。脲酶活性与速效钾含量显著正相关,而与速效磷含量极显著负相关。硝酸还原酶活性与碱解氮、有机质含量和细菌总数显著或极显著正相关,而与速效磷含量和真菌总数显著或极显著负相关。脱氢酶活性与碱解氮、速效钾、有机质含量和细菌总数显著或极显著正相关,而与速效磷含量和真菌总数显著或极显著负相关(表2)。由于马铃薯连作障碍的产生,导致土壤中氮营养、碳营养、有机质含量显著降低,而速效磷含量则显著增加,引起pH值下降,同时真菌数量的上升,造成马铃薯感染病害的可能性增加。因此,速效磷是马铃薯连作障碍形成的主要因子,这与芋艿连作障碍的成因[34]类似。
在套作苦荞的土壤中,有机质含量与所有酶活性显著或极显著正相关,真菌数量与蔗糖酶和脱氢酶活性极显著负相关,细菌数量与蔗糖酶、硝酸还原酶和脱氢酶活性极显著正相关,放线菌数量则与除磷酸酶之外的所有酶活性显著或极显著正相关(表2)。结合套作苦荞以后,相对2020年,马铃薯土壤酶活性增强,有机质和速效钾含量增加,细菌和放线菌数量上升,表明有机质对各种土壤酶发挥其活性具有促进作用,并能增强作物根际土壤微生物活动,是马铃薯套作苦荞缓解连作障碍的关键因素。
综上,马铃薯套作苦荞,可通过改善土壤pH值,促进苦荞对磷肥、有机质、氮素、碳素的合理利用,提高土壤有机质和钾肥含量,提升细菌总数/真菌总数比值,改善土壤微生态环境,降低病原菌数量,提升生防菌数量,从而使马铃薯与苦荞套作形成科学合理的种间效应,以缓解马铃薯连作障碍。研究结果可为马铃薯套作苦荞栽培模式的推广提供科学依据。
参考文献:
[1]周华兰,彭亚丽,李婷,等.马铃薯连作对土壤理化性质和生物学特性的影响[J].湖南农业大学学报(自然科学版),2019,45(6):611-616.
[2]胡新喜,冯艳青,雷艳,等.不同施氮水平下秋马铃薯的生长以及StNR和StAT的表达[J].中国农业科学院,2015,42(10):1974-1982.
[3]何雅祺,王鑫鑫,张弛,等.间作、套种模式在中药材栽培中的效应研究进展[J].江苏农业学报,2021,37(4):1077-1083.
[4]江春明,马琨,代晓华,等.间作栽培对连作马铃薯根际土壤微生物区系的影响[J].生态与农村环境学报,2013,29(6):711-716.
[5]魏常慧,刘亚军,冶秀香,等.马铃薯‖玉米间作栽培对土壤和作物的影响[J].浙江大学学报(农业与生命科学版),2017,43(1):54-64.
[6]王海燕,王晓玲.马铃薯间作蚕豆的效益评价与栽培研究[J].内蒙古农业科技,2007,12(3):37-39.
[7]李春花,陈蕤坤,黄金亮,等.苦荞种质资源遗传多样性分析及抗霜霉病种质筛选[J].南方农业学报,2020,51(4):740-747.
[8]周美亮.西藏荞麦的创新利用和发展前景[J].西藏农业科技,2018,40(1):7-10.
[9]龙梦千,冯洁,李振东,等.苦荞‖马铃薯间作对苦荞生长及产量的影响[J].分子植物育种:1-20[2023-01-28].http://kns.cnki.net/kcms/detail/46.1068.S.20210913.1929.009.html.
[10]毛春,蔡飛,程国尧,等.马铃薯套作秋播苦荞栽培试验研究[J].现代农业科技,2012(8):61,63.
[11]汤睿,李基光,王艳兰,等.苦荞与幼龄林油茶、茶树、猕猴桃套种的产量及效益分析[J].湖南农业科学,2021(10):9-10.
[12]王玉琴,尹亚丽,李世雄.不同退化程度高寒草甸土壤理化性质及酶活性分析[J].生态环境学报,2019,28(6):1108-1116.
[13]AdamczykB,KilpelinenP,KitunenV,etal.PotentialactivitiesofenzymesinvolvedinN,C,PandScyclinginborealforestsoilunderdifferenttreespecies[J].Pedobiologia,2014,57(2):97-102.
[14]马玲,马琨,杨桂丽,等.马铃薯连作栽培对土壤微生物多样性的影响[J].中国生态农业学报,2015,23(5):589-596.
[15]李金花,王蒂,柴兆祥,等.甘肃省马铃薯镰刀菌干腐病优势病原的分离鉴定[J].植物病理学报,2011,41(5):456-463.
[16]秦伟,陈昆,赵跃锋.大蒜套作对温室茄子根际土壤微生物数量与土壤酶活性的影响[J].山西农业科学,2018,46(11):1831-1833,1902.
[17]白艳茹,马建华,樊明寿.马铃薯连作对土壤酶活性的影响[J].作物杂志,2010,136(3):34-36.
[18]要凯,赵章平,康益晨,等.沟垄覆膜对连作马铃薯土壤酶活性、理化性状及产量的影响[J].作物学报,2019,45(8):1286-1292.
[19]胡建利,王德建,孫瑞娟,等.长江下游典型地区农田土壤肥力变化——以常熟市为例[J].土壤学报,2008,45(6):1087-1094.
[20]WangYF,QinYZ,ChaiQ,etal.Interspeciesinteractionsinrelationtorootdistributionacrosstherootingprofileinwheat-maizeintercroppingunderdifferentplantdensities[J].FrontiersinPlantScience,2018,9:483.
[21]卢维宏,张乃明,张丽,等.增效肥料对设施栽培小白菜生长及土壤酶活性的影响[J].南方农业学报,2019,50(9):2022-2028.
[22]CardosoNC,PapadopoulosAO,KanaBD.Mycobacteriumsmegmatisdoesnotdisplayfunctionalredundancyinnitratereductaseenzymes[J].PLoSOne,2021,16(1):e0245745.
[23]TanXP,WangZQ,LuGN,etal.KineticsofsoildehydrogenaseinresponsetoexogenousCdtoxicity[J].JournalofHazardousMaterials,2017,329(5):299-309.
[24]赵庆龙,宋宪亮,孙学振,等.蒜棉、麦棉套作对土壤微生物数量及相关酶活性的影响[J].植物营养与肥料学报,2011,17(6):1474-1480.
[25]王佳,陈伟,张强,等.黄土丘陵沟壑区低氮胁迫对苦荞生长及土壤养分的影响[J].西南农业学报,2021,34(11):2412-2421.
[26]焦念元,汪江涛,张均,等.化学调控和施磷对玉米‖花生间作磷吸收利用和间作优势的影响[J].中国生态农业学报,2015,23(9):1093-1101.
[27]崔霎霎,宋全昊,高小丽,等.糜子不同种植方式对土壤酶活性及养分的影响[J].植物营养与肥料学报,2015,21(1):234-240.
[28]JangidK,WilliamsMA,FranzluebbersAJ,etal.Land-usehistoryhasastrongerimpactonsoilmicrobialcommunitycompositionthanabovegroundvegetationandsoilproperties[J].SoilBiologyandBiochemistry,2011,43(10):2184-2193.
[29]酒鹃鹃,李永梅,王梦雪,等.玉米大豆间作对坡耕地红壤团聚体分布及稳定性的影响[J].江苏农业科学,2021,49(22):219-228.
[30]LiQS,WuLK,ChenJ,etal.Biochemicalandmicrobialpropertiesofrhizospheresundermaize‖peanutintercropping[J].JournalofIntegrativeAgriculture,2016,15(1):101-110.
[31]马琨,杨桂丽,马玲,等.间作栽培对连作马铃薯根际土壤微生物群落的影响[J].生态学报,2016,36(10):2987-2995.
[32]刘亚军,马琨,李越,等.马铃薯间作栽培对土壤微生物群落结构与功能的影响[J].核农学报,2018,32(6):1186-1194.
[33]李旭红,王雪玉,李章波,等.长期温室栽培对玉米生长生理和土壤酶活性及养分平衡的影响[J].江苏农业科学,2022,50(6):60-65.
[34]张翰林,郑宪清,王一斐,等.连作芋艿套作对土壤养分及酶活性的影响[J].环境污染与防治,2014,36(11):55-59,66.
