水曲柳幼苗水力结构和光合生理对光强梯度变化的耦合响应
2023-03-30王爱英郝广友宁秋蕊殷笑寒周永姣
孔 鑫,王爱英,郝广友,宁秋蕊,王 淼,殷笑寒,周永姣
(1.中国科学院沈阳应用生态研究所森林生态与管理重点实验室,辽宁 沈阳 110016;2.中国科学院大学,北京 100049;3.辽宁省城市有害生物治理与生态安全重点实验室,沈阳大学生命科学与工程学院,辽宁 沈阳 110044;4.Forest Ecology and Forest Management Group,Wageningen University,Wageningen 476700,Netherlands)
光照是影响植物生长发育及存活的重要生态因子,也是森林演替过程中促进物种更替的主要环境因素[1-2]。森林中的光环境在时间和空间上都处于不断变化之中[3],太阳光穿过林冠受到阻挡与反射,使到达林下的光强只有冠层的1%~5%[4]。树木幼苗在弱光条件下生长缓慢,常需要在林下生长几年至几十年才能到达冠层[5]。为了能够在弱光下生存,树木在权衡生长-存活关系的过程中进化出了不同的生活史策略,通常演替初期的先锋树种生长快速但在弱光下的存活率低,而演替后期的树种则具有较高的存活率和较低的生长速率[6-7]。同时,同一种树木在不同的发育阶段对光的需求以及对光环境变化的响应也存在较大差异,树木在幼龄阶段对光照条件的适应和响应是森林更新和群落演替的重要内在驱动力[6,8]。
植物适应异质性光环境的能力对其生存至关重要,在不同的光环境下,植物通过调节形态结构、生理特征等来提高其生存适合度[2,8-9]。通常,弱光下植物的比叶面积增大,叶片更大、更薄,有利于增强光能捕获力,积累更多的光合产物,提高其耐阴性[5,10]。光照强度的不同还会引起生物量积累和分配的改变,这也是植物适应不同光环境的重要机制,如弱光下植物通过降低根生物量占比、根冠比,增加叶、茎生物量占比来增强光能捕获力,遮阴处理也会增加植物叶绿素含量和表观量子效率(AQY),降低光饱和点(LSP)、光补偿点(LCP)以及暗呼吸速率(Rd)等[11-14]。这些可塑性响应可提高叶片对光能的利用效率并减少叶片及个体水平上呼吸作用的能量消耗,从而维持植物在弱光下的碳平衡[13-14]。

水曲柳(Fraxinusmandshurica)是中国东北“三大硬阔树种”之一,也是构成我国东北地区典型针阔混交林的重要优势树种,该树种在生长过程中表现出高生产力和高竞争力[29]。水曲柳在苗期具有一定的耐阴性,其幼苗在相对高的光强下更新数量多且存活率高,但在弱光下更新较差,因而光强可能是限制水曲柳幼苗更新的主要环境因子[1,30]。本研究以2年生水曲柳幼苗为研究对象,探讨其生长、光合特性以及水力性状对不同光强的响应,以期验证以下假设:①与低光强相比较,高光强下水曲柳幼苗可构建效率更高的水分传输系统,因而具有更高的器官和整株水平的水力导度;②在不同光照条件的可塑性响应过程中,水曲柳幼苗的水力结构与叶片光合生理之间存在协同关系。
1 材料与方法
1.1 试验地概况
试验在中国科学院长白山森林生态系统定位站(128°28′ E,42°24′ N)进行。该区域具有受季风影响强烈的温带大陆性气候,冬季寒冷而漫长,夏季温暖多雨且短暂[31]。年均降雨量600~900 mm,多集中于生长季;年均气温4.5~7.8 ℃[32]。该地区典型的地带性植被类型为阔叶红松林,其结构复杂、组成独特且生物多样性丰富,主要建群树种有红松(Pinuskoraiensis)、水曲柳、椴树(Tiliatuan)、蒙古栎(Quercusmongolica)和五角枫(Acerpictum)等[33]。
1.2 供试材料及试验设计
以2年生水曲柳幼苗为试验材料,幼苗来源于吉林省安图县白河镇光明育苗公司基地。于2017年5月17日,将80株生长健康、长势相近的幼苗移栽到直径为30 cm、高为28 cm的花盆中,每盆盛入干土约6 kg,每盆栽植1株,浇足水分。在野外空旷区生长1个月后,将幼苗随机分成4组,每组20盆分别置于4个处理区中。利用不同层数的黑色尼龙网(对照、1层、2层、3层)搭建遮阴棚进行不同光照处理,遮阴棚间距3 m以上,以减少各处理间相互干扰。于试验期间晴天的中午,使用便携式光合仪(Li-6400,Li-Cor.,USA)光量子探头测定对照和3个遮阴处理下的光合有效辐射强度,即光通量密度(PPFD),每个遮阴棚中选择3个位置进行测定并计算平均值,3个遮阴处理的相对光强分别约为全光照的60%、30%和15%。试验期间(2017年7—8月)当地全光照辐射强度(排除阴天)日最大值平均为(1 533.80±44.79) μmol/(m2·s)。所有光照处理下的幼苗均采用相同的抚育管理措施(除草和浇水等),每隔1周调整同一光照处理下各个花盆的摆放位置,以确保其受光条件基本一致。于2018年7—8月测定各光照处理下幼苗的生长、光合生理和水力性状等指标。由于15%相对光强处理下只有少量幼苗存活且幼苗个体太小,无法进行光合特性和水力性状的测定,该处理仅进行了幼苗生长及生物量测定。
1.3 指标测定方法
1.3.1 气体交换参数测定
于2018年7月中旬晴朗的上午(9:00—11:30),用Li-6400便携式光合测定仪测定水曲柳叶片的气孔导度(Gs)、净光合速率(Pn)。测定时设定叶室内PPFD为1 400 μmol/(m2·s),CO2含量400 μmol/mol,叶室温度23.5~26.5 ℃。每个光照处理下随机选取3株幼苗,每株幼苗选取枝条上部第2~3片成熟健康的叶片。
1.3.2 光响应曲线测定
每个光照处理下随机选取3株幼苗,利用Li-6400便携式光合测定仪测定水曲柳的光响应曲线,测定时设置CO2含量400 μmol/mol,叶室温度23.5~26.5 ℃,控制叶室内PPFD由强到弱依次为:1 400、1 000、600、400、200、100、50、20、0 μmol/(m2·s)。为确定最大净光合速率(Pn,max),测定前将幼苗放在饱和光强下诱导30 min,测定期间叶片在每个光强下停留200 s。对测定数据利用Photosynthesis软件进行光响应曲线拟合,计算出Pn,max、光补偿点(LCP)、光饱和点(LSP)、暗呼吸速率(Rd)和表观量子效率(AQY)。
1.3.3 植物水力导度测定
每个光照处理下随机选取4株水曲柳幼苗测定植株水力导度。于测定日清晨,在距离土壤表面5 cm处给幼苗主干围一圈塑料膜,向塑料膜注入蒸馏水的同时将其剪断,地下部分立刻连接到充满水的硅胶管上,防止空气进入导管,剪下的地上部分也迅速插入清水中。测定前,将幼苗主干截面在水下切平,剥去切口端2 cm左右的树皮,连接至高压导水率仪(HPFM Gen3;Dynamax Corp.,Elkhart,IN,USA)的压力耦合器上[34-35]。仪器设置为稳态测定模式,在0.5 MPa压强下将去气的蒸馏水灌注枝条内,达到稳定流率(通常需要10 min左右)获得地上部分的水力导度(Kshoot)。根系水力导度(Kroot)采用HPFM瞬时测定方法[36]。测定导水率后,将枝条上所有的叶片用扫描仪扫描,利用Image J(US National Institutes of Health,Bethesda,MD,USA)计算总叶片面积(LA)。根据每株植物Kshoot和Kroot的测定结果,计算出整株植物水力导度(Kplant),并用LA对测定值标准化。最后将全部叶片在70 ℃烘箱下烘干至质量恒定,称其干质量(M),计算比叶面积(SLA)。
1.3.4 生长性状及器官间生物量分配测定
于8月中旬测定水曲柳幼苗的生长性状及各器官间生物量分配。每个光照处理下随机选出3~4株幼苗测定株高和基径后收获整株,将其分成粗根(直径 >2 mm)、细根、茎、叶柄和叶片5部分分别装入信封中,置于70 ℃烘箱下烘干至质量恒定,称量各部分生物量。计算幼苗的根(RMR)、茎(SMR)、叶生物量占比(LMR)以及根冠比(R/S)。此外,每株幼苗随机选取部分叶片进行扫描并用Image J程序计算叶片大小。
1.3.5 枝条木质部解剖观测

1.4 数据处理
通过表型可塑性指数(Pi)评估各指标的环境可塑性,Pi= (Mmax-Mmin) /Mmax。其中:Mmax是某个指标在4种光照处理下的最大平均值,Mmin是最小平均值;Pi在0~1之间,其值越大表示可塑性越高[37]。
利用SPSS 20.0对试验数据进行单因素方差分析(ANOVA),采用最小显著性差异法(LSD)进行差异显著性检验(α= 0.05),用Sigmaplot 14.0作图。
2 结果与分析
2.1 不同光照处理下水曲柳幼苗生长及生物量分配特征
光照强度对水曲柳幼苗生长及生物量分配整体上存在较大影响(表1)。水曲柳幼苗的株高、基径、总叶面积以及比叶面积在不同光照处理下存在显著差异(P<0.05),其中株高和总叶面积均在60%相对光强下较大,在30%和15%相对光强下最小。随着光照强度的减弱,水曲柳幼苗的基径逐渐减小,但比叶面积呈增加的趋势。与100%相对光强相比,遮阴处理下水曲柳幼苗各器官生物量、总生物量、根生物量占比以及根冠比较低,而茎、叶和地上生物量占比较高。
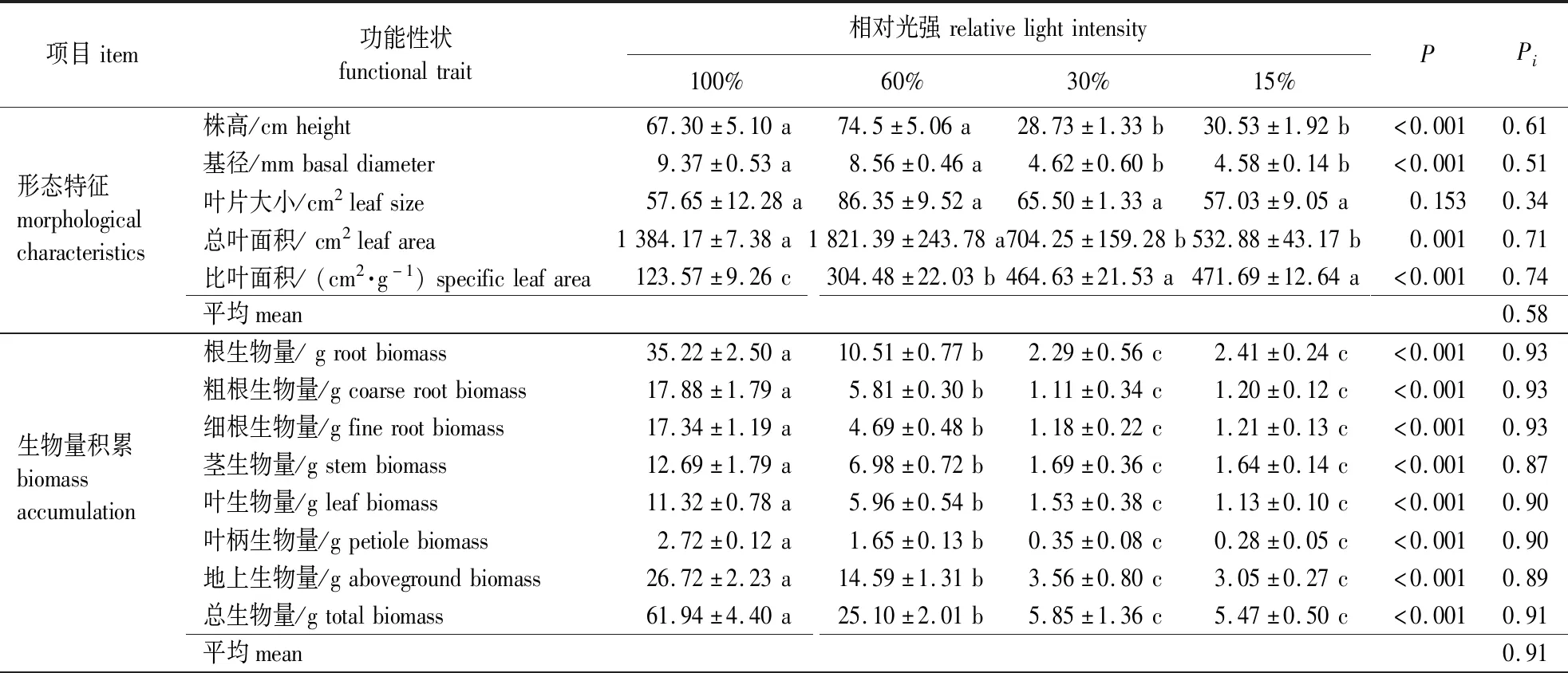
表1 不同光照处理下水曲柳幼苗生长、生物量分配及光合生理特征Table 1 The growth,biomass allocation and photosynthetic characteristics of Fraxinus mandshurica seedlings grown under different light treatments

表1(续)
水曲柳幼苗各功能性状的可塑性指数(Pi)平均值由高到低依次为:生物量积累(0.91)>形态特征(0.58)>光合生理(0.57)>生物量分配(0.34)。在各形态指标中以比叶面积Pi最高(0.74),其次是总叶面积Pi(0.71),叶片大小Pi(0.34)最低;而叶片光合生理指标的Pi以Pn(0.91)、Gs(0.90)和LCP(0.73)较高(表1)。
2.2 不同光照处理下水曲柳幼苗的光合特性
在3种光照处理下,水曲柳幼苗Pn随PPFD的变化总体上呈先上升后下降的趋势(图1A)。100%相对光强下生长的幼苗,当叶室内PPFD小于600 μmol/(m2·s)时,Pn表现出随PPFD增加而增加的趋势,之后随PPFD增加略有下降,但仍然保持在最大值附近。而在60%和30%相对光强下,当PPFD分别小于200和400 μmol/(m2·s)时,Pn表现出随PPFD增加而增加的趋势;随后,在较高光强时Pn随着PPFD的增加均表现出显著下降的趋势,在PPFD达到1 400 μmol/(m2·s)时,60%和30%相对光强下幼苗的Pn分别只有其最大值的38.2%和15.4%。遮阴处理显著降低了水曲柳的Pn,在相同PPFD下,100%相对光强下生长的幼苗其Pn值最大。
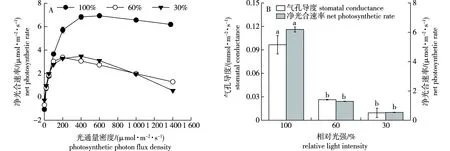
图1 不同光照处理下水曲柳幼苗的光响应曲线、气孔导度和净光合速率Fig.1 The light response curve,stomatal conductance and net photosynthetic rate of Fraxinus mandshurica seedlings grown under different light treatments
在不同光照处理下,水曲柳幼苗的AQY、LSP、Pn,max以及LCP均存在显著差异(表1,P<0.05)。遮阴处理降低了LSP、Pn,max以及LCP,却使得AQY增大,各指标测定值分别在100%和30%相对光强下最大。随遮阴程度的增加,水曲柳幼苗的Gs和Pn总体上呈下降的趋势,以100%相对光强最高,但两指标在60%和30%相对光强下无显著差异(图1B,P>0.05)。
2.3 不同光照处理下水曲柳幼苗的水力学特征
在100%相对光强下,水曲柳幼苗的Kplant、Kroot以及Kshoot均显著高于60%和30%相对光强下的相应值(P<0.05),但Kroot、Kshoot在60%和30%相对光强间并无显著差异(P>0.05)(图2)。

图2 不同光照处理下生长的水曲柳幼苗的水力导度(A)和木质部解剖结构(B)Fig.2 The hydraulic conductance standardized by total leaf area (A) and xylem anatomical structure (B) of Fraxinus mandshurica seedlings grown under different light treatments
水曲柳幼苗的导管密度和导管直径在不同光照处理下存在显著差异(P<0.05),在60%相对光强下导管密度最小,而Dv和D95最大。
3 讨 论
水曲柳为喜光树种,虽然在幼苗阶段该树种可以忍受一定的郁蔽环境,但弱光下并不利于其生长和存活。在不同的光照处理下,水曲柳幼苗主要通过调节自身形态和生理特征来适应变化的光环境,这些性状的可塑性响应增强了幼苗在弱光环境下对光的捕获能力和光能利用效率,从而提高了水曲柳的生存适合度。本研究结果表明,水曲柳幼苗的水力结构特征在不同光照处理下存在显著差异,水分传输功能与叶片气体交换之间在响应生长光环境变化时存在显著的耦合关系。
3.1 光强对水曲柳幼苗生长及光合特性的影响

3.2 光强对水曲柳幼苗水力性状的影响
随着相对光强的增加,水曲柳幼苗Kshoot、Kroot和Kplant都显著增加(P<0.05),这与文献中Kshoot随光强增加而增加以及阳生幼苗的Kroot高于阴生幼苗Kroot的结果相一致[42-43]。在土壤-植物-大气连续体中,植物根系是液态水分流动的最大阻力所在,而Kroot是用来表征植物根系吸水能力的一个重要水力学参数,对Kplant至关重要[44]。根系的水分吸收和传导效率受根系的生长和生物量分配的强烈影响[21-22],如细根生物量的减少会显著降低根系的水分吸收面积以及水分传导能力[35,45]。本研究发现水曲柳幼苗在高光照下具有较高的细根生物量,这有利于增强根系对水分的传输能力。另外,植物水分传输能力的改变通常伴随着水分传导组织结构(如导管或管胞直径)的变化,根据哈根泊肃叶方程(Hagen-Poiseuille equation),导管或管胞的理论导水率与其直径的4次方成正比,较大的导管或管胞可显著提高木质部的水分传输效率,但直径较大导管上的纹孔往往对干旱诱导的气种扩散(air seeding)更加敏感,导致较低的干旱栓塞抵抗力[16,46-47]。本研究结果表明,与30%相对光强相比,水曲柳幼苗在60%相对光强下具有较大的导管,其导水率也较高;但在100%相对光强下水曲柳幼苗具有更高的Kshoot,而其导管直径较小,导管密度较大,这可能是强光环境下生长的植物面临更高的蒸腾需求,木质部水势更低更多,但是较小的导管有助于其提高栓塞抵抗力,从而增强植物在水分胁迫条件下的生存能力[24,47-48]。水曲柳幼苗提高茎干木质部的水分传输安全性,对保护其碳投入成本较高的茎干具有重要的适应意义,而且茎干在整株水分传导阻力的占比相对较小[49-50],即使茎干水力导度在强光条件下有所下降,对幼苗Kplant的限制也较小。
3.3 水曲柳幼苗对光强梯度变化的水分-光合生理协同响应
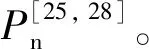
水曲柳是地带性植被演替顶级群落(阔叶红松林)的优势树种[29],其幼苗具有一定的耐阴性,可以通过一定的可塑性调整来适应异质性的光环境。本研究发现强光照环境下更适宜水曲柳幼苗的生长,这与韩有志等[1]发现较高光强下水曲柳幼苗更新和存活的数量明显高于较弱光强的结果一致。在郁闭度较高的林下或较小的林窗中,光合有效辐射低[52],水曲柳幼苗的生长受到限制,较高的可塑性可增强其耐阴性从而提高生存能力。当有新的林窗形成、林下光照条件得到明显改善时[53],较高的可塑性可使水曲柳幼苗有效地利用充足的光能迅速生长,并占据优势地位,增加其最终到达林冠层的概率。因此,在森林经营管理中,应适当采取抚育择伐等措施,降低林冠郁闭度,增加林窗面积,促进水曲柳等喜光目标树种幼苗的生长。
总之,虽然水曲柳幼苗能够通过形态和生理的可塑性调整来适应一定程度的弱光环境,进而提高其光竞争力和生存适合度,但其生物量积累、水分传输能力以及叶片气体交换速率均在100%相对光强下最大,表明水曲柳幼苗对光强仍有较高的需求。水曲柳幼苗的光合和水力性状随光照强度的改变都具有较高的可塑性,二者对光强梯度变化的响应存在显著耦合关系,较高的可塑性增强了水曲柳在森林异质性较高的光环境中的生存适合度。通过模拟林下光环境,从幼苗更新角度探讨水曲柳对不同光照强度的响应及机理,对促进森林更新具有重要的理论和应用研究价值。
